Télécharge Modelisation des instabilies du cortex d’actine et plus Lectures au format PDF de Physique sur Docsity uniquement!
THESE DE DOCTORAT DE L’UNIVERSIT` E´
PIERRE & MARIE CURIE - PARIS 6
Ecole doctorale : Physique de la R´^ ´ egion Parisienne
Sp´ecialit´e : Interface Physique - Biologie
Pr´esent´ee par :
Guillaume SALBREUX
Sujet de la th`ese :
Mod´elisation des instabilit´es
du cortex d’actine
Th`ese dirig´ee par Jacques PROST et Jean-Fran¸cois JOANNY
soutenue publiquement le 17 Octobre 2008 devant un jury compos´e de :
M. Guillaume CHARRAS Rapporteur M. Jean-Baptiste FOURNIER Rapporteur Mme. Martine BENAMAR M. Frank JULICHER M. Jacques PROST M. Jean-Fran¸cois JOANNY Membre invit´e M. Stephan GRILL Membre invit´e
Remerciements
La fin de la these est un moment tres agr´eable : on peut regarder derriere soi et faire des bilans. Je me revois en effet il y a trois ans,a la rentr´ee scolaire 2005, avec ma regle et mon cahier de texte :a cette ´epoque, je n’´etais pas encore docteur, et j’´etais donc bien na¨ıf. En particulier je croyais encore que l’ATP ´etait un classement de tennis, que le cortex ´etait une souris d´esireuse de conqu´erir le monde, et des Prost c´elebres je ne connaissais que le coureur automobile. Heureusement mes deux directeurs de these, Jean-Fran¸cois Joanny et Jacques Prost, m’ont rapidement montr´e l’ampleur du travail. Leur culture, leur vivacit´e, leur intarissable curiosit´e, leur rapidit´e dans l’´echange des id´ees, sont presque spec- taculaires. Chaque discussion avec eux est une le¸con en soi : non seulement a travers les connaissances scientifiques qu’ils transmettent, mais aussi par la fa¸con d’aborder les problemes, d’en s´electionner les aspects int´eressants, de rebondir pour proposer de nouvelles id´ees. Leur cot´e humain est tout aussi impressionnant : toujours a l’´ecoute, jamais ils n’inhibent leurs ´etudiants, ni n’imposent une vision des choses a priori : tout ce qu’ils enseignent est par la force de l’exemple. J’ai du mala imaginer un environnement plus favorable pour un th´esard, et je suis content d’avoir l’occasion de leur exprimer toute ma reconnaissance et mon admiration. Plus largement je remercie les membres de mon jury : Martine Ben Amar, Jean- Baptiste Fournier, Guillaume Charras, Stephan Grill et Frank J¨ulicher se sont joints a Jacques et Jean-Fran¸cois pour constituer un jury impressionnant, dont les ques- tions ont ´et´e particulierement riches et perspicaces. Merci en particulier a mes deux rapporteurs, G. Charras et J.B. Fournier, d’avoir pris le temps de lire mon manus- crit et d’en avoir fait des rapports d´etaill´es. Et bien sˆur le th´eoricien serait vraiment triste s’il n’avait pas son ami l’exp´e- rimentateur pour lui montrer ses manips et faire le vrai travail. Je remercie donc Pramod Pullarkat, qui est maintenant a l’universit´e de Bangalore mais qui, lorsque nous avons travaill´e ensemble, ´etaita l’universit´e de Bayreuth. J’ai beaucoup appr´e- ci´e nos discussions sur ses oscillations, ainsi que ces petits cabanons a vin chaud qui font le charme des march´es de no¨el bavarois. Plusa l’Est, je remercie ´egalement Ewa Paluch et son groupe au MPICBG a Dresde, et en particulier ceux avec qui nous avons parl´e de blebs : Jean-Yves Tinevez, Ulrike Schulze, Julia Roensch et Ja- kub Sedzinski. Lors de mes trois voyages dans cette ville nous avons beaucoup parl´e, comment´e la vie du bleb sous toutes ses formes, et nous avons pass´e de nombreux moments dans le quartier cool et branch´e de Dresde que je n’h´esiterai pasa qualifier de sympatoches. Cette collaboration a ´et´e particulierement agr´eable grˆacea l’´ecoute, a l’´energie d´eploy´ee, au souci de vraiment r´epondre aux questions qui animent Ewa et son groupe. Dans le mˆeme labo, j’ai beaucoup appr´eci´e les discussions que nous avons eu avec Mirjam Mayer et Stephan Grilla propos de leur travaux sur C. Ele- gans, et pas tres loin au PKS, je remercie ´egalement Frank J¨ulicher qui m’a recu l’an dernier dans son labo pour une journ´ee qui m’a beaucoup appris. De retoura
Table des mati`eres
- I Introduction
- A Quelques aspects du cytosquelette
- A.1 -Principaux constituants
- A.2 -Le cortex d’actine
- B Mod´elisation du cytosquelette : les gels actifs
- B.1 -Principe d’une description hydrodynamique
- B.2 -Le cas des gels actifs
- B.3 -D´erivation pour un n´ematique
- II Le cortex d’actine : r´egulation de l’´epaisseur et instabilit´es actives
- A Cortex dans une cellule sph´erique
- A.1 -G´eom´etrie du probl`eme et orientation des filaments
- A.2 -Polym´erisation et d´epolym´erisation de la couche
- A.3 -Pression hydrostatique et osmotique
- A.4 -Contraintes dans la couche et tension du cortex
- A.5 -R´egulation de l’´epaisseur par le treadmilling
- B Instabilit´es induites par les myosines : formation de trous
- B.1 -Instabilit´e dans une couche en l’absence de treadmilling
- B.2 -Instabilit´e dans une couche en pr´esence de treadmilling
- B.3 -Lien avec la formation de blebs
- C Conclusions
- III Oscillations de forme de fibroblastes en suspension
- A R´esultats exp´erimentaux
- A.1 -Caract´eristiques de l’oscillation
- A.2 -Role de l’adh´esion au substrat
- A.3 -Influence du calcium externe
- A.4 -Influence des traitements agissant sur la myosine
- B Description du mod`ele
- B.1 -Canaux m´ecanosensibles
- B.2 -R´egulation de l’activit´e des myosines par le calcium
- B.3 -Dynamique de la couche
- C Diagramme des phases
- C.1 -Comparaison entre les modes
- C.2 -Diagramme de stabilit´e du mode
- C.3 -Influence du treadmilling
- D Conclusions 2 Table des mati`eres
- IV Formation d’anneaux contractiles
- A Description du mod`ele
- A.1 -Couplage hydrodynamique entre flux et param`etre d’ordre
- A.2 -Mod
ele effectifa deux dimensions
- B Applications
- B.1 -Formation d’un anneau dans une cellule `a g´eom´etrie cylindrique
- B.2 -Fermeture d’un anneau contractile dans le plan du cortex
- B.3 -Cytocin`ese dans une cellule sph´erique suspendue
- B.4 -Flux corticaux dans l’embryon de C. Elegans
- C Conclusions
- V Blebs induits par ablation du cortex
- A Exp´eriences
- A.1 -Montage exp´erimental
- A.2 -R´esultats exp´erimentaux
- B Equilibre du bleb et ´elasticit´e de la cellule
- B.1 -Elasticit´e du cortex
- B.2 -Elasticite du milieu interne
- C Dynamique
- C.1 -Perm´eation du cytosol `a travers le cytoplasme
- C.2 -Friction de la membrane
- C.3 -Comparaison aux donn´ees
- D Repolym´erisation du cortex
- D.1 -Influence sur le volume du bleb
- D.2 -Ouverture du trou
- E Conclusions
- A Notations
- B Polynˆomes de Legendre
- C Th´eorie de coque mince active pour une sph`ere faiblement d´eform´ee
2 Table des mati`eres
contre, une description des ph´enomenes non-lin´eaires dominants peut ˆetre ajout´eea ce modele g´en´erique. Dans cette these, nous utilisons cette approche pour ´etudier quelques problemes relatifsa la couche d’actine corticale. Le cortex ´etant une structure dynamique, nous sommes fr´equemment amen´es a interpr´eter son comportement par une analyse de stabilit´e. En fonction du contexte, nous cherchonsa identifier les parametres pertinents contrˆolant les instabilit´es eta ´etablir les seuils critiques au-dela desquels elles se d´eveloppent. Dans le premier chapitre, nous donnons un aper¸cu des connaissances de biologie cellulaire n´ecessairesa notre ´etude, et nous introduisons le modele des gels actifs. Dans le chapitre 2, nous discutons de diff´erents m´ecanismes par lesquels la cellule pourrait contrˆoler l’´epaisseur de la couche corticale, et nous montrons qu’elle peut se d´estabiliser spontan´ement sous l’effet des contraintes exerc´ees par les myosines. Dans le chapitre 3, nous pr´esentons les travaux exp´erimentaux de P. Pullarkat sur l’oscillation spontan´ee de fibroblastes en suspension, et nous exposons une analyse de ce ph´enomene ou l’instabilit´e ´evoqu´ee dans le chapitre 2 est coupl´ee a la r´eponse calcique de canaux m´ecanosensibles. Des pr´edictions th´eoriques sont obtenues en ac- cord avec l’exp´erience. Dans le chapitre 4, nous incluons dans notre description du cortex un parametre d’ordre n´ematique correspondant a l’orientation des filaments, et nous d´erivons des ´equations effectivesa deux dimensions pour le cortex. Nous montrons que le ph´enomene de flux cortical et de formation d’anneau de filaments,a l’œuvre dans divers systemes biologiques, peut ˆetre interpr´et´e par la r´eponse hydro- dynamique du cortexa un gradient de concentration des myosines. Les blebs sont des protusions membranaires d´epourvues de couche corticale : dans le chapitre 5 nous exposons les r´esultats d’une collaboration avec le groupe d’E. Paluch au MPI-CBG a Dresde sur la croissance de blebs form´es artificiellement par ablation laser du cortex. Nous proposons diverses hypotheses pour les m´ecanismes contrˆolant l’´equilibre et la dynamique du bleb, et nous d´erivons syst´ematiquement des pr´edictions th´eoriques que nous comparons aux r´esultats exp´erimentaux.
Chapitre I
Introduction
A Quelques aspects du cytosquelette
Le cytosquelette d´esigne l’ensemble des constituants cellulaires qui d´eterminent l’architecture de la cellule. Trois familles sont g´en´eralement distingu´ees : les struc- tures impliquant l’actine et la myosine, les microtubules et une troisieme cat´egorie moins ´etudi´ee, les filaments interm´ediaires. Le cytosquelette assure les taches m´e- caniques de la cellule : il contrˆole sa forme, lui permet de migrer et de se diviser physiquement en deux cellules filles lors de la cytocinese. La source d’´energie pour tous ces m´ecanismes est l’hydrolyse de l’ad´enosine triphosphate (ATP), un nucl´eo- tide utilis´e par tous les ˆetres vivants et qui constitue la r´eserve ´energ´etique de la cellule. Cette ´energie est utilis´ee pour polym´eriser des filaments ou actionner des moteurs mol´eculaires, ces deux processus pouvant g´en´erer des forces. Dans cette partie nous pr´esentons les m´ecanismes fondamentaux impliqu´es dans le fonctionne- ment du cytosquelette.
A.1 Principaux constituants
A.1.1 L’actine et les prot´eines qui la r´egulent
Actine G et actine F
Fig. I.1: Repr´esentation sch´ematique d’un filament d’actine
L’actine est une prot´eine tres abondante : elle repr´esente de 5 a 10% du nombre total de prot´eines d’une cellule non musculaire [ 12 ]. Elle existe dans la cellule sous forme monom´erique (actine G) et sous forme de filaments polym´erises (actine F). Le monomere d’actine est une petite prot´eine, d’une masse mol´eculaire de 43kDa,
A. Quelques aspects du cytosquelette 5
ou Cm est la concentration de monomeres dans le milieu, kof fp et kbof f sont les taux de
d´epolym´erisation et kponCm et kbonCm les taux de polym´erisation. En l’absence d’ATP, le filament doit ˆetre a l’´equilibre thermodynamique, ce qui impose que ses deux extr´emit´es aient la mˆeme constante de dissociation KD, qui est aussi la concentration de monomeres `a l’´equilibre Cmc :
Cmc = Kd =
kpof f kpon
kof fb kbon
(I.3)
La concentration Cmc, de l’ordre de 1μM, est aussi appel´ee concentration critique, car pour une concentration de monom`eres qui lui est inf´erieure, les filaments polym´e- risent, alors que pour une concentration plus ´elev´ee ils d´epolym´erisent. En pr´esence
actine-ATP actine-ATP actine-ADP actine-ADP bout barb´e bout pointu bout barb´e bout pointu kon(μM−^1 .s−^1 ) 11.6 1.3 3.8 0. kof f (s−^1 ) 1.4 0.8 7.2 0. Kd(μM) 0.12 0.62 1.9 1.
Tab. I.1: Cin´etique de polym´erisation et de d´epolym´erisation des bouts barb´es et pointus d’un filament d’actine, en presence d’ATP (reproduit de [ 116 ])
d’ATP, l’´energie apport´ee par l’hydrolyse de l’ATP li´e aux monomeres d’actine place le filament hors d’´equilibre et les constantes de dissociation au bout barb´e et au bout pointu deviennent diff´erentes (les taux et constantes sont reproduits dans le tableau A.1.1). Le m´ecanisme est le suivant : les monomeres d’actine possedent un site de liaisona l’ATP. Les monomeres d’actine-ATP sont incorpor´es au bout barb´e, puis ils sont hydrolys´es dans le filament en monomeres d’actine-ADP, qui ont une constante de dissociation plus ´elev´ee. Dans ce cas il n’y a pas de vrai ´equilibre pour le filament, mais selon la valeur des concentrations l’une des deux extr´emit´es polym´erise plus que l’autre. Les valeurs du tableau A.1.1 montrent que le filament polym´erise pr´ef´e- rentiellement sous forme ATP et au bout barb´e. Pour cette raison on appelle aussi le bout barb´e “bout plus” et le bout pointu “bout moins”. En particulier, et comme illustr´e sur la fig. I.3, pour une concentration en monomeres KDb < Cm < Kdp les filaments polym´erisent au bout barb´e et d´epolym´erisent au bout pointu : dans cette situation les monomeres sont ins´er´es `a un bout du filament sous forme ATP, pro- gressent dans le filament, sont hydrolys´es en ADP et finalement se d´etachent au bout pointu. Par analogie avec la progression d’un tapis roulant, on appelle ce m´ecanisme
le “treadmilling”. En particulier pour une concentration Cm =
kbof f −kpof f kbon−kpon ≃^0.^14 μM il existe un ´etat stationnaire qui v´erifie dn dtp = dn dtb , pour lequel les filaments ont une longueur constante.
Les prot´eines associ´ees `a l’actine
La concentration d’actine dans la cellule est tres ´elev´ee (entre 10 et 300μM, [^116 ]), tres au-del`a des constantes de dissociation des deux bouts du filament. Malgr´e
6 Chapitre I. Introduction
Fig. I.3: Courbe de gauche : en l’absence d’ATP les deux extr´emit´es ont la mˆeme concentration critique, qui est aussi la concentration d’´equilibre. Courbe de droite : en pr´esence d’ATP les concentrations critiques sont diff´erentes. Dans la r´egion cen- trale les filaments polym´erisent au bout barb´e et d´epolym´erisent au bout pointu : la position des monomeres dans le filament ´evolue t comme sur un tapis roulant, un ph´enomene appel´e “treadmilling”
cela, les filaments ne se forment pas spontan´ement, car in vivo la polym´erisation et la d´epolym´erisation des filaments sont contrˆol´ees par des prot´eines ayant des fonctions diverses vis-`a-vis de l’actine (voir tableau I.2) :
- Des prot´eines de s´equestration s’attachent aux monom
eres et les empˆechent de polym´eriser. La thymosine s’attache assez faiblementa l’actine (Kd ≃ 1. 2 − - 6 μM avec l’actine ATP [ 116 ]), mais est tr
es abondante dans certaines cellules en regard de l’actine (20μM pour 18μM d’actine dans les oeufs de Xenope par exemple[ 116 ]), suffisamment pour s´equestrer la plupart des monomeres d’actine. Dans les r´egions ou la polym´erisation de l’actine est requise, une seconde prot´eine est activ´ee, la profiline. La profiline entre en comp´etition avec la thymosine pour la liaison aux monomeres, et est capable de s’attacher a l’actine plus fortement (KD ≃ 0. 1 μM [ 116 ]). Contrairementa la thymosine la profiline contribue a la polym´erisation de l’actine en catalysant l’echange de l’ADP en ATP dans le monomere. Elle empˆeche ´egalement la polym´erisation aux bouts pointus, ce qui favorise la directionnalite de la polym´erisation, du bout plus vers le bout moins.
- La prot´eine de coiffage (capping protein) recouvre le bout barb´e avec une grande affinit´e (Kd = 0. 1 nM [ 129 ]) et inhibe ainsi la polym´erisation des fila- ments. Sa cin´etique d’attachement est rapide (4s−^1 [ 116 ]), ce qui semble ˆetre un facteur important dans la r´egulation de la longueur des filaments [ 19 ]. A la membrane une famille de lipides, les phosphoinositides, peuvent sous leur forme phosphoryl´ee enlever la coiffe du filament, ce qui stimule localement la polym´erisation (PIP2 [ 129 ]). La tropomoduline est une prot´eine qui coiffe le bout pointu des filaments.
8 Chapitre I. Introduction
s’attache (distance ∼ 14 nm). L’α-actinine forme ´egalement des liaisons entre filaments parall`eles, mais de facon plus lˆache (distance ∼ 40 nm).
- Des prot´eines de coupure (severing protein), telles que la gelsoline ou l’ADF- cofiline, s’attachent aux filaments et les sectionnent. L’ADF-cofiline applique une torsion au filament qui entraˆıne sa rupture [ 93 ]. La gelsoline est activ´ee par le calcium et par PIP2, et peut
a la fois sectionner les filaments et coiffer leur bout barb´e [ 138 ] : ces deux effets contribuenta diminuer la taille typique des filaments dans un r´eseau, ce pourquoi elle porte ce nom, ´etant susceptible d’induire des transitions sol-gel dans les r´eseaux d’actine [ 155 ]. L’ensemble de ces m´ecanismes donne lieu `a une cin´etique complexe de fabrication et d’´elimination d’un r´eseau, que nous d´etaillons dans le paragraphe suivant dans le cas du lamellipode.
Un r´eseau d’actine dans la cellule : le lamellipode
Fig. I.4: Formation, croissance et d´epolym´erisation du gel d’actine dans le lamel- lipode, adapt´e de [ 117 ]
Comme nous venons de le mentionner, des prot´eines r´egulent la formation des filaments et leur assemblage en structures plus complexes. Le lamellipode est une protusion cellulaire impliqu´ee de facon essentielle dans la motilit´e cellulaire sur un substrat. Il se forme a l’avant de la cellule en mouvement et fournit une partie de la force m´ecanique n´ecessairea son avanc´ee [ 136 ]. Sa forme plane rend son observation plus facile que celle des autres structures du cytosquelette : de ce fait il a ´et´e beau- coup ´etudi´e, et a permis de comprendre la facon dont les interactions mol´eculaires d´ecrites dans le paragraphe pr´ec´edent donnent naissance a une structurea l’´echelle de la cellule (fig. I.4). L’actine est polym´eris´ee sous sa forme ATP `a la membrane par
A. Quelques aspects du cytosquelette 9
Arp2/3, selon le m´ecanisme de nucleation dendritique : Arp2/3 est activ´e, nucl´ee un nouveau filament qui s’allonge rapidement, jusqu’a ce qu’une prot´eine de coiffage ar- rˆete sa croissance. Les nouveaux filaments ´etant ajout´es aux anciens avec un angle de 70 ◦, le r´eseau d’actine du lamellipode est branch´e avec cet angle caract´eristique. Le gel croˆıt ainsi globalement depuis la membrane a une vitesse d’environ 1μm.min−^1 [ 146 ], puis il est d´epolym´eris´e par l’ADF-cofiline. Seuls les monomeres d’actine-ADP peuvent ˆetre rompus par l’ADF-cofiline, le temps d’hydrolyse de l’ATP maintient donc une distance minimale de gel non d´epolym´eris´e [ 117 ]. Le gel du lamellipode se d´eplace globalement en un mouvement de tapis roulant (treadmilling), diff´erent du treadmilling ´evoqu´e dans la partie A.1.1 pour un filament d’actine simple en ce que le r´eseau entier est impliqu´e dans le mouvement.
A.1.2 La myosine II
Fig. I.5: Structure g´en´erale de la myosine II : les deux tˆetes globulaires contiennent les sites de liaison a l’actine, la queue permet aux myosines de s’associer entre elles pour former des oligomeres (adapt´e de [ 2 ])
Les myosines sont des moteurs mol´eculaires qui utilisent l’´energie produite par l’hydrolyse de l’ATP pour fournir une force en interagissant avec un filament d’actine. Pres de 150 types de myosines ont ´et´e r´epertori´es [ 2 ], mais nous ne nous int´eressons ici qu’a la myosine II non-musculaire qui interagit avec la couche d’actine corticale. Notons que d’autres isoformes de la myosine II existent dans toutes les cellules mus- culaires. Elles sont associ´ees a l’actine dans une structure contractile, le sarcomere, responsable de la contraction des fibres musculaires. La mol´ecule de myosine II est constitu´ee des ´el´ements suivants (fig. I.5) :
Tˆetes
Les deux tˆetes globulaires sont constitu´ees d’un domaine catalytique ATP-ase, qui contient ´egalement le site de fixation a l’actine, et d’un domaine r´egulateur (ou cou) qui relie la tˆetea la queue. L’hydrolyse de l’ATP induit un changement de conformation qui d´eplace le cou avec un angle de ∼ 70 ◦^ vers le bout barb´e. Dans le mod`ele du “coup de force” la myosine utilise ce changement conformationnel pour exercer une force sur un filament d’actine. L’interaction actine/myosine d´ecrit le cycle suivant, sch´ematis´e sur la fig. I.6 :
- Configuration attach´ee : Lorsque ni ATP, ni ADP ne sont li´es `a la myosine, la tˆete est fortement attach´ee au filament d’actine, on parle d’´etat “rigor”.
A. Quelques aspects du cytosquelette 11
- Configuration libre Lorsqu’une mol´ecule d’ATP se lie
a la myosine, cette der- niere se d´etache du filament. L’hydrolyse de l’ATP en ADP-Pi provoque un changement de conformation, d´epla¸cant la tˆete d’environ 10nm, et armant la myosine pour la production de force `a l’´etape suivante du cycle. - Coup de force L’accrochage au filament d’actine provoque la lib´eration du phosphate inorganique Pi. La myosine retrouve alors sa configuration initiale, ´energ´etiquement favorable, et est ainsi capable d’exercer une force sur le fi- lament. Simultan´ement, la tˆete perd la mol´ecule d’ADP et retourne `a l’´etat rigor, fortement attach´e au filament.
Queue
La queue mesure environ 150nm de long et 2nm de diametre, et elle permeta la myosine de polym´eriser en filaments bipolaires (contenant plusieurs tˆetes de chaque cot´e de la queue). Cette propri´et´e est cruciale puisqu’elle permet `a la myosine d’exercer des forces relatives entre plusieurs filaments (fig. I.6c)
Chaˆınes l´eg`eres de r´egulation
A l’ensemble tˆetes+queue, constitu´e de deux chaˆınes lourdes d’environ 2000 acides amines, s’ajoute deux chaˆınes l´egeresa chaque tˆete de moins de 200 acides amin´es. L’une de ces deux chaˆınes est importante pour la r´egulation de l’activit´e de la myosine, et est appel´ee RLC pour “regulatory light chain”, ou chaˆıne l´egere r´egulatrice (l’autre chaˆıne est appel´ee chaˆıne l´egere essentielle et stabilise la queue des chaˆınes lourdes). Lorsque les RLC sont non phosphoryl´ees, la queue de la myo- sine recouvre le site de liaison a l’actine pr´esent sur la tˆete, ce qui empˆechea la fois l’attachement aux filaments et l’oligom´erisation (voir fig. I.6b). Les prot´eines MLCK (pour myosin light chain kinase) et MLCP (myosin light chain phosphatase) r´egulent la phosphorylation des chaˆınes l´egeres : la premiere active les myosines en les phosphorylant et la seconde a l’effet inverse. Ces deux prot´eines sont elles-mˆemes r´egul´ees par des voies de signalisation sur lesquelles nous revenons dans la partie A.2.1.
A.1.3 Les microtubules
Les microtubules sont des polymeres r´esultants de l’assemblage dynamique de dimeres de tubuline α et β en protofilaments, eux-mˆemes associ´es en un cylindre creux d’une ´epaisseur de 25nm. Leur nucl´eation est assur´ee par des complexes ap- pel´es MTOC (microtubule organizing center), dont font partie en particulier les centrosomes. Un complexe de γ-tubuline pr´esent dans le MTOC assure la forma- tion du cylindre et coiffe le bout moins des filaments. Les microtubules croissent donc par polym´erisation a leur bout plus, de facon tres dynamique : des ´evenements de “catastrophe” et de “sauvetage”, au cours desquels le filament d´epolym´erise tres rapidement, alternent avec les phases de croissance. Ces instabilit´es sont dues a la pr´esence d’une couronne de tubuline-GTP au bout pointu qui est d´estabilis´ee par l’hydrolyse des monomeres GDP en tubuline-GDP [ 2 ] Le rˆole des microtubules dans les m´ecanismes cellulaires sont multiples [ 2 ], mais nous ne nous int´eressons ici
12 Chapitre I. Introduction
qu’a leur rˆole en association avec le cortex d’actine. On peut distinguer deux types d’interactions entre l’actine et les microtubules : r´egulatrices, lorsqu’une voie de si- gnalisation est impliqu´ee, ou structurelles, lorsqu’il existe un lien physique entre les deux ´el´ements [ 125 ]. Un lien physique entre le cortex et le bout plus des microtubules astraux pourrait ainsi contrˆoler m´ecaniquement la position des centrosomes, et jouer un rˆole essentiel dans le positionnement du fuseau central lors de la division cellulaire [ 142 ]. Dans le lamellipode, un flux conjoint de microtubule et d’actine a ´et´e observ´e [ 148 ], les microtubules ´etant entraˆın´es par le flux r´etrograde. L’interaction r´egula- trice semble essentiellement m´edi´ee par les petites GTP-ases : la d´epolym´erisation des microtubules active RhoA [ 123 ], et leur assemblage active Rac1. A l’aide des ces voies de r´egulations, les microtubules sp´ecifient ainsi le site de constriction dans la cytocinese (voir partie A.2.3). Ils d´eterminent ´egalement la polarit´e de la cellule dans la motilit´e sur un substrat [ 125 ]. Celle-ci pourrait s’´etablir grace `a une boucle de retroaction positive entre les GTPases et les microtubules [ 150 ] : Rho active la formine mDia, ce qui d´estabilise les MTs et favorise donc Rho. Sym´etriquement, Rac active la kinase Pak1 qui stimule la croissance des microtubules.
A.2 Le cortex d’actine
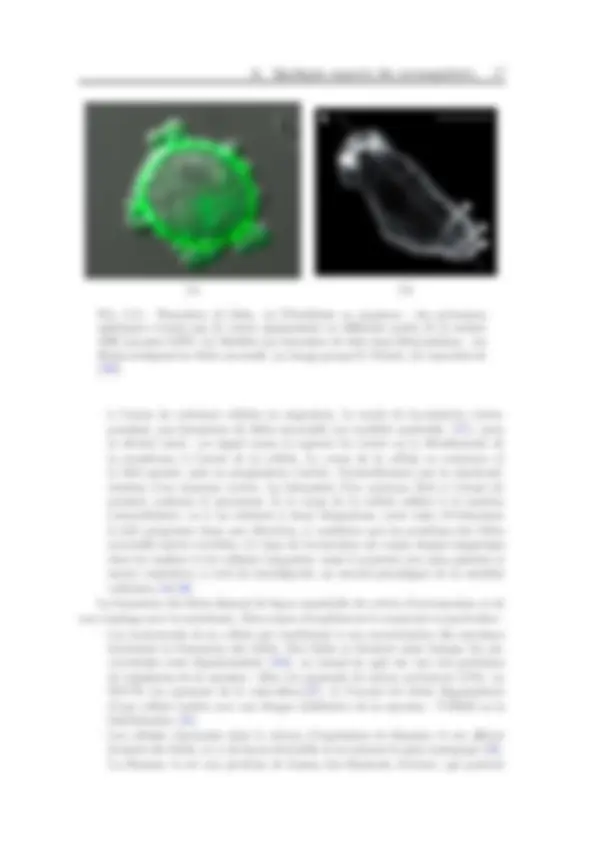
(a) (b) (c)
Fig. I.7: Cortex d’actine visualis´e dans differentes conditions. (a) cellule motile : la cellule migre dans la direction de la fleche et un lamellipode est pr´esenta l’avant de la cellule. (image actine-GFP) (b) Fibroblaste dont les microtubules ont ´et´e d´epolym´eris´es : sous l’effet de la tension du cortex la cellule s’arrondit (image myosine-GFP) (c) Cellule en division : le cortex d’actomyosine est plus dense dans le plan ´equatorial qui se contracte (image DIC/myosine GFP) (a) reproduit de [ 48 ], (b) et (c) groupe E. Paluch.
A.2.1 Propri´et´es
Organisation du cortex
Le cortex d’actine est une couche de filaments d’actine reli´es a la membrane de la cellule, d’environ 1μm d’´epaisseur (fig. I.7). Les myosines II y sont tres concen- tr´ees, et les forces qu’elles y font naˆıtre en font une structure contractile qui donne
14 Chapitre I. Introduction
Attachement du cortex `a la membrane
Les acteurs mol´eculaires de la liaison du cortex avec la membrane ne sont pas non plus tr`es bien connus. Deux questions se posent : quelles prot´eines nucl´eent et polym´erisent les filaments, et quelles prot´eines assurent l’adh´esion de la membrane au cortex.
- Polym´erisation Arp2/3, le nucl´eateur d’actine dans le lamellipode, parait ˆetre un candidat naturel pour polym´eriser le cortex. Il est effectivement pr´esent dans le cortex de Dictyostelium, mais il est absent de son anneau contractile lors de la division cellulaire [ 67 ], il n’est pas pr´esent
a la membrane d’un bleb de cellule tumorale humaine lors de la reformation du cortex [ 23 ], et il n’est pas n´ecessairea la polym´erisation de l’actine dans C Elegans [ 132 ], au contraire des formines. Ces dernieres (voir la partie sur la nucl´eation de l’actine) pourraient jouer le role de nucl´eateur, bien que mDia1 ne soit pas non plus pr´esent au cortex des blebs. mDia2 pourrait ˆetre un candidat plausible : des cellules Hela soumisesa DIP, une prot´eine inhibitrice de mDia2, forment des blebs de fa¸con anormale, ce qui sugg`ere que le cortex est affaibli en son absence [ 39 ]. - Adh´esion cortex-membrane Des prot´eines qui lient sp´ecifiquement l’actine et des prot´eines transmembranaires existent : la ponticuline est une glycopro- t´eine transmembranaire qui se lie au cot´e des filaments d’actine [ 153 ], et est n´ecessaire au maintien du cortex dans Dictyostelium [ 64 ]. La famille ERM, constitu´ee de trois prot´eines tr
es proches (ezrine-rad´eeine-mo´esine), aurait un rˆole essentiel dans l’adh´esion membrane-cortex. Ces trois prot´eines possedent un site d’attachement au bout barb´e des filaments a leur bout C-terminal, et pourraient s’attachera la queue cytoplasmique de nombreuses prot´eines transmembranaires a leur bout N-terminal [ 15 ]. Elles sont normalement au- toinhib´ees et sont activ´ees par phosphorylation d’un de leurs r´esidus, ce qui permeta la cellule de les r´eguler. La Rho-kinase en particulier est capable de les activer. Elles sont localis´ees dans la cellule a l’interface cortex/membrane au niveau de l’anneau contractile, des microvilli ou des sites d’adh´esion [ 145 ]. L’ezrine est n´ecessairea la liaison du cortex a la membrane lors de sa reforma- tion dans un bleb mais n’est pas requise pour la nucl´eation de l’actine [ 23 ]. Un article r´ecent [ 85 ] montre que la mo´esine est essentiellea l’arrondissement de la cellule qui pr´ecede la mitose, et peut mˆeme la d´eclencher en l’absence de myo- sine. Les auteurs attribuent cette propri´et´ea l’apparition d’un fort couplage membrane/cortex qui augmente le module de courbure du cortex.
R´egulation de l’actine et de la myosine : Rho et MLCK
Dans cette these nous avons ´et´e amen´esa d´ecrire en particulier deux voies de signalisation de la r´egulation du cortex (voir fig. I.9) :
- La petite GTPase RhoA a un double effet sur le cortex : d’une part elle ac- tive la polym´erisation d’actine en activant les formines [ 53 ], d’autre part elle active la Rho-kinase (´egalement appel´ee ROCK ou ROK) qui phosphoryle la chaine l´eg
ere r´egulatrice des myosines (RLC) [ 4 ] et inhibe la MLCP (myosin light chain phosphatase) qui d´ephosphoryle RLC [ 75 ]. Ces deux derniers effets contribuenta augmenter l’activit´e des myosines.
A. Quelques aspects du cytosquelette 15
RhoA
Rho-kinase MLC phosphatase
Myosines
MLC Ca Calmoduline kinase 2+
Formines Polymérisationde l'actine
phosphoryle
phosphoryle déphosphoryle
se lie à (^) se lie à
se lie à
phosphoryle
se lie à
Fig. I.9: R´egulation de l’actine et de la myosine par Rho et MLCK. Nous indiquons par une fl`eche l’activation et par une barre l’inhibition d’une prot´eine.
- MLCK (pour myosin light chain kinase) phosphoryle ´egalement RLC, et est r´egul´ee par le calcium et la calmoduline. Dans le chapitre III nous d´ecrirons en d´etail ces interactions. D’autres prot´eines r´egulatrices de l’actine existent : mentionnons les petites GT- Pases Rac et Cdc42. Ces deux prot´eines peuvent activer les prot´eines WASP, qui elles-mˆemes activent Arp2/3 et donc la polym´erisation d’actine. Rac a ´egalement pour cible la PI(4)P-5 kinase, qui active
a la membrane les phospholipides phosphoi- nositides en PIP2. PIP2 a plusieurs effets favorisant la polym´erisation de filaments : il peut activer Wasp et inhiber le facteur de d´epolym´erisation ADF-cofiline [ 154 ]. Dans la cellule motile, Rac et Cdc42 seraient plutˆot actifsa l’avant de la cellule, et contrˆolerait ainsi le lamellipode, alors que Rho r´egulerait la contraction du cortex a l’arriere de la cellule en activant la myosine II. C’est pourquoi les travaux exp´eri- mentaux sur le cortex que nous d´ecrivons dans cette th`ese utilisent principalement la voie de r´egulation par Rho.
A.2.2 Motilit´e et blebs
Une des fonctions du cytosquelette d’actine est de participer a la locomotion des cellules. La motilit´e cellulaire est n´ecessairea un grand nombre de ph´enomenes biologiques essentiels : elle permet la migration des fibroblastes dans la r´eparation des blessures, les mouvements de cellule impliqu´es dans la morphog´enese, ou la progression des cones de croissance neuronaux dans le d´eveloppement du syst`eme nerveux.
Role dans la motilit´e induite par un lamellipode
Le cortex intervient dans la motilit´e sur un substrat en assurant la contraction de la partie arriere de la cellule (fig. I.10). La cellule motile est polaris´ee dans la direction du mouvement : le lamellipode assure l’extension d’un front de migrationa l’avant de la cellule, et le cortex la contraction du corps cellulaire [ 12 ]. Cette derniere propri´et´e est indispensable : les cellules dont le gene de la myosine II a ´et´e supprim´e ne peuvent plus se d´eplacer. Il est remarquable en revanche qu’en d´epolym´erisant le lamellipode et le cortex `a l’avant d’une cellule de tumeur ´epith´eliale, la motilit´e est