Scarica Elementi e composti. Metabolismo e più Sbobinature in PDF di Biochimica solo su Docsity!
BIOCHIMICA
Lezione 1 Elemento, atomo, composti Elemento sostanza formata da atomi dello stesso tipo uguali tra loro. Un elemento è formato da circa 100 atomi. Composto è costituito da due o più elementi di tipo diverso che possono essere scissi nelle sue parti. Atomo è la parte più piccola di un elemento e ne conserva le proprietà chimico-fisiche; esso è formato da: Elettrone (e- = -1): hanno carica negativa; girano attorno al nucleo situate nelle nubi elettroniche e hanno una grandezza di 9,11 x 10*-28; Protone (p+ = +1): hanno carica positiva e sono situati nel nucleo; hanno una massa maggiore rispetto a quella di e- di circa 2000 volte maggiore; Neutrone (n = 0): hanno carica neutra e sono sempre situati nel nucleo. Ogni elemento ha: Numero atomico indicato con la lettera Z ed indica il numero di p+ (che in un elemento neutro equivale ad un numero di e-); Numero di massa indicato con la lettera A ed indica il numero di neutroni e protoni presenti nel nucleo. Tramite questo valore sottraendolo a Z, si trova il numero di neutroni. Isotopo sono due atomi che hanno uguale numero di Z ma diverso numero A. Gli isotopi sono stabili e molto radioattivi quindi decadono ed emettono radiazioni. La loro reattività varia in base alla massa. Es. C12 A = 12 C13 A = 13 Z rimane invariato nell’elemento varia solamente A. La massa di un atomo per definizione, è l’unità di massa atomica di 1/12 della massa atomica della massa del C12. Tavola periodica Nella tavola periodica viene indicato:
Elemento, A e Z (numero di massa e numero atomico), T° fusione, Elettronegatività. La tavola periodica è divisa in periodi (righe) e gruppi (colonne); gli elementi che sono sullo stesso periodo hanno e- del guscio esterno con lo stesso livello, mentre gli elementi che si trovano nello stesso gruppo hanno gli stessi e- di valenza. Gli elementi nella tavola periodica sono divisi in base alle caratteristiche chimico- fisiche in: Metalli solidi, duri e ottimi per condurre calore e corrente elettrica; Non metalli sono situati sulla parte dx della tavola periodica e H è compreso tra questi, hanno caratteristiche opposte ai metalli, Semimetalli hanno caratteristiche intermedie tra i due tipi precedenti, Gas nobili chimicamente sono inerti ed hanno raggiunto l’ottetto. Tramite la posizione degli elementi nella tavola periodica, si possono valutare: Energia di prima ionizzazione energia necessaria per allontanare un e- dallo strato esterno. Ciò porta alla formazione di un catione e più il legame tra l’atomo ed e- è “forte” maggiore sarà l’energia che c’è da immettere per rompere tale legame ma anche sarà elevata la quantità d’energia liberata. Affinità elettronica energia liberata quando si aggiunge un e- in un atomo neutro; in questo caso si viene a creare un anione. Elettronegatività la media aritmetica tra l’energia di prima ionizzazione e l’affinità elettronica. Tale valore indica la “fame di elettroni” intesa come la necessità di acquistare o cedere e- per raggiungere l’ottetto. Tale valore aumenta nella tavola periodica da sx verso dx e dal basso verso l’alto; gli elementi più elettronegativi sono F,O,N Cl. Le proprietà dell’elemento, tra cui la reattività dipendono dagli e-; essi sono la più piccola parte di un atomo che, grazie alla fisica quantistica le teorie che girano attorno agli elettroni: Teoria atomica moderna e- sono considerati come particelle ma anche come onde energetiche; in qualsiasi caso non è possibile definire velocità e posizione di e-, si può solamente definire, in termini di probabilità, la regione dello spazio dove si trova e-.
Legame ionico dato da METALLO + NON METALLO dove uno dei due cede un e- e diventa un catione mentre l’altro acquista e- e diventa anione. A stabilire chi cede e chi accetta e- è il valore di elettronegatività. I due elementi sono attratti fino ad una distanza minima di attrazione. Legame covalente formato da NON METALLO + NON METALLO e consiste nella condivisione di uno o più e- per permettere a due elementi di completare la valenza infatti l’elettrone in comune non viene ceduto/acquisito ma condiviso quindi e- “gira” attorno ad entrambi gli elementi. I legami che si possono formare sono di tipo singolo, doppio o triplo. Osservando i legami intermolecolari, si ricorda che il più importante è il legame ad idrogeno. Esso è una forza intermolecolare dove H è legato covalentemente ad un elemento molto elettronegativo di un’altra molecola. Vengono a crearsi dei dipoli (positivi e negativi), dove H è attratto all’elemento elettronegativo presente sull’altra molecola. È il tipo di legame tra le molecole più forte e tipico delle molecole d’acqua che non hanno una struttura lineare dati i dipoli (essi indicano anche l’addensamento di carica) ; tale legame ha un ruolo importantissimo a livello biologico (nelle proteine). Carbonio Il carbonio è il maggior costituente dei composti organici (oltre che H e O e a volte P). C ha Z=6 e ha 4 orbitali semi occupati nello stato fondamentale Se però il C entra nello stato eccitato, uno degli elettroni presenti in 2s trasla in 2p per la regola di Pauli e l’orbitale s sale di livello energetico.
In tale modo C può formare 4 legami covalenti stabili. Molto spesso all’interno della molecola sono presenti dei gruppi funzionali che determinano le caratteristiche della molecola. Tali legami sono molto deboli e, con le reazioni tra altre molecole, sono quei legami che si rompono e tali gruppi vengono persi. C-C-C = scheletro carbonioso della molecola; una molecola a livello biochimico è definita insatura se presenta doppi e/o tripli legami mentre è definita satura se presenta solamente legami singoli. Ibridazione del carbonio Il carbonio può andare contro ad ibridazione: tale procedimento porta all’aumento “livello” di un orbitale di “livello” inferiore che porta l’ottenimento di orbitali ibridi equivalenti gli uni tra gli altri a livello energetico. Essi sono definiti isoenergetici. I tipi di orbitali ibridi sono: sp3 1s +3p, ha energia maggiore dell’orbitale s ma minore dell’orbitale p e porta la geometria molecolare di un tetraedro con angolo 109,5° tipica degli alcani. sp2 1s +2p; ha come geometria molecolare triangolare con angolo a 120° e presenza di doppi legami. È la forma tipica degli alcheni. sp 1s+1p, ha come geometria molecolare lineare con angolo di 180° ed è la tipica configurazione degli alchini.
Gli idrocarburi sono molecole dove l’atomo di C si lega solamente ad H quindi non ci sono etero atomi e in tale categoria si hanno gli alcani, alcheni ed alchini quindi molecole lineari definiti alifatici; ma troviamo in questa classificazione anche gli idrocarburi aromatici con una tipica struttura ciclica come il benzene; in tale struttura ci sono doppi legami all’interno della molecola. I gruppi alchilici sono denominati contando il numero di carboni e aggiungendo il prefisso -ANO; es. CH3-CH2-CH2-CH3 butano. Mentre i radicali alchilici sono ramificazioni della catena principale della molecola derivata dall’alcano che perde un atomo di H; il suffisso è -ILE e per denominare una molecola si prende il nome della molecola e si aggiunge desinenza -ILE. Lo stato di ossidazione Dall’ossidazione dei composti ossigenati, l’organismo ricava energia. Il numero d’ossidazione rappresenta la carica formale che si attribuisce ad un elemento in un composto supponendo che i legami siano di tipo ionico in modo da segnare gli h+ all’atomo più elettronegativo. Es. metano supponiamo che i legami siano di tipo ionico (quindi si ha un trasferimento di e-). C è più elettronegativo quindi dato che è legato a 4 H ha 4 cariche negative se il legame fosse di tipo ionico le chiamerebbe a se le cariche negative. Es. metanolo 3 cariche negative e una positiva dato il legame con O. Le reazioni redox
Molecole alla base dello studio: Amminoacidi sono venti e sono l’unità fondamentale delle proteine ed ormoni e sono substrati energetici Basi nucleotidiche 3 in comune tra DNA ed RNA e una differente ed oltre a costituire acidi nucleici e ed enzimi hanno un ruolo di trasportatori d’energia Zuccheri substrati energetici ed elementi di riconoscimento cellulare Lipidi sono fondamentali perché costituiscono le membrane cellulari e sono una fonte d’energia. Amminoacidi Le proteine hanno un ruolo cruciale in varie funzioni vitali come emoglobina fondamentale per trasposto 02 nel sangue ed mioglobina che immagazzina 0, gli enzimi sono catalizzatori di reazioni, altre proteine come actina e miosina del muscolo hanno azione contrattile quindi funzione di movimento, ruolo segnale come ormoni es. insulina, strutturale come collagene, e protezione come anticorpi. Le proteine sono polimeri di unità amminoacidica; l’ordine sequenziale degli amminoacidi è definito dal DNA. L’amminoacido è una molecola organica con struttura base comune a tutti gli
amminoacidi con un carbonio centrale definito α legato ad H a un gruppo -COOH ed
-NH2 e con gruppo -R che varia da amminoacido ad amminoacido differenziandoli. Tutti tranne la glicina hanno C alfa con 4 legami diversi mentre la glicina ha 2 H legati a C. Tali amminoacidi soni definiti asimmetrici o chirali (esclusa la glicina): quindi gli amminoacidi uguali tra di loro” sono speculari l’un l’altro ma non sovrapponibili definiti per questo enantiomeri. Sono definiti come stereoisomeri L o D in grado di far ruotare i piano della luce polarizzata verso dx o sx. gli amminoacidi delle proteine sono della serie L. la chiralità è una qualità non appartenete alla glicina. Nella formula possiamo notare -COO- e -NH3+ perché a PH fisiologico 7 il gruppo carbossilico è dissociato ha perso H+ mentre il gruppo amminico acquista un H+ quindi le due cariche si annullano. Sempre a PH fisiologico i 20 amminoacidi sono classificati in base al gruppo -R in: Polari o non polari, Carica positiva o negativa,
Aromatici. Si ha una suddivisione alimentare: Essenziali assunti con la dieta perché il corpo non li sintetizza, Non essenziali il corpo li sintetizza e quindi non è necessario produrli con l’alimentazione, Semi-essenziali il corpo li produce solo in certe condizione ed è preferibile assumerli in modo da non sintetizzarli a partire dagli essenziali da cui derivano. Condizionatamente- essenziali in certe condizioni assunti con la dieta a causa di patologie. Osserviamo gli amminoacidi: Alifatici non polari: _Glicina; _Prolina ha una conformazione particolare perché ciclizza sulla catena rendendo la struttura della proteina che la contiene rigida; _Metionina contiene un atomo di zolfo non terminale quindi non è terminale ma è legato a due atomi di C. Alifatici polari: idrofili solubili (non carichi) _Cisteina: secondo atomo con S ma è un tiolo infatti ha -SH, importante perché tale gruppo tende a legare con un altro -SH di una cisteina portando un’interazione tra i due S perdendo H formando ponte di solfuro. Quando si hanno 2 cisteine legate tra loro con ponte di disolfuro si chiama cistina; _Serina e Treonina la polarità è data dal gruppo ossidrile, ci sono eteroatomo (O e H) che permettono di essere idrofili; _ Asparagina e Glutammato hanno anch’essi N.
alcuni enzimi avendo un determinato amminoacido che è esposto un amminoacido e può accettare gruppo fosfato e tale fosforilazione va ad influenzare l’attività dell’enzima inibendo o attivando l’enzima e ciò permette la regolazione della loro attività metabolica dell’enzima. Gli amminoacidi sono molecole ionizzabili da cui grado e tipo dipende dal PH da cui dipende l’attività della molecola. Il PH si calcola con: PH= -log |H+| -log della concentrazione degli idrogenioni. Ricordiamo che gli enzimi sono proteine con funzione cruciale per il metabolismo e le attività corporee. Punto isoelettrico è il valore di PH al quale le cariche positive e negative dell’amminoacido si equivalgono e la carica è uguale a zero; il concetto si estende anche sulle proteine e varia da molecola a molecola (come con l’amminoacido). Valori di PH superiori al punto isoelettrico portano l’amminoacido ad avere una carica negativa mentre valori bassi portano una carica positiva. Il punto isoelettrico varia da amminoacido ad amminoacido. Peptidi e proteine Un polimero di aa. può formare peptide (da 2 a 1000 amminoacidi con peso < dalton) o proteina (> 5000 dalton con migliaia di amminoacidi). La proteina più grande è la titina (3 milioni dalton). Gli amminoacidi si legano tra loro attraverso un legame peptidico tra il gruppo carbossilico di un aa. e il gruppo amminico di un altro aa. con la perdita di H2O grazie all’interazione del OH del carbossilico e H dell’amminico. Il legame peptidico è molto forte che si rompe in condizioni estreme di T°, PH o enzimi idrolitici cioè proteasi. Per convenzione una proteina si indica con l’inizio dalla parte sx ammino-terminale e con la fine dx carbossilica. Il legame peptidico è una struttura planare e rigida, la libera rotazione è impedita infatti si hanno punti di rigidità nella catena. La proteina ha 4 gradi di struttura: Primaria la sequenza degli amminoacidi legati tra loro con legame peptidico; è il primo livello strutturale della proteina e corrisponde alla catena polipeptidica e l’ordine degli amminoacidi dipende dal DNA; la struttura tridimensionale della proteina che raggiungerà dipende dagli amminoacidi legati tra di loro in una certa sequenza; la si può ottenere tale struttura quando si denatura la proteina andando a rompere le interazioni delle strutture successive. Possono esistere infine proteine con identici amminoacidi ma diversa sequenza da cui dipende poi anche la funzione della proteina infatti alterazioni minime amminoacidiche possono alterare la proteina ed essere alla base di malattie es. anemia falciforme dove due amminoacidi sono sostituiti con altri due. Secondaria prima organizzazione tridimensionale; è intrinseco nel legame peptidico la capacità di formare un legame H che permette di ottenere conformazioni 3D tipiche dove H e la catena R non sono coinvolte nella struttura infatti le catene sporgono all’esterno non integrandosi:
_α elica ripiegamento elicoidale dello scheletro peptidico con catene laterali rivolte all’esterno; quasi sempre destrogiro e si forma con iterazione gruppi -NH legame peptidico e -COO del legame presente ogni 4 residui amminoacidici più avanti con passo 0,54nm. È il legame peptidico che determina la formazione dei legami H. in alcune proteine 2 o più filamenti possono avvolgersi ulteriormente l’uno sull’altro formando una struttura detta cordone intrecciato tipica delle proteine fibrose molto resistenti. Gli amminoacidi idrofobici sono rivolti all’interno della struttura per evitarne il contatto col solvente. _β foglietto ripiegamento disteso con distanza tra i due amminoacidi è di 0,35nm e si ha una forma a zig -zag dove le catene laterali non intervengo nella struttura e sono perpendicolari al piano della struttura. Più foglietti beta associati tra loro in modo longitudinale formano una struttura a foglietto ripiegato tutto ciò grazie a legami H. l’andamento dei foglietti può essere parallelo, antiparallelo o misto. Terziaria strutture secondarie organizzate nello spazio; è la struttura tridimensionale complessa della proteina, i motivi appena visti si organizzano in strutture diverse uniti da anse (aa. legati tra loro che non formano strutture secondarie) di struttura variabile non casuali che ne da e specifiche caratteristiche. Una struttura terziaria formata è definita come dominio o subunità. Perché si formi tale struttura si ha un ripiegamento della catena polipeptidica porta il posizionamento di uno dei residui idrofobici amminoacidici all’interno della catena, si avvicinano per interazione idrofobiche, di cariche opposte o di cisteina (-SH) per formare ponte di disolfuro (unico legame covalente tra le catene laterali). Una proteina lineare è definita unfolding e si organizza nello spazio dando un folding con parti idrofobe all’interno e le parti che possono fare interazioni all’esterno. Ciò determina la struttura nativa di una proteina. Nella formazione della struttura terziaria -R sono fondamentali tanto è che se sono lontani nella sequenza possono interagire grazie a vari ripiegamenti. Tutte le interazioni stabilizzano la struttura terziaria persa in condizioni denaturanti; in caso di denaturazione si agisce con dei fattori tra cui temperatura elevate e PH si va a rompere i legami della proteina tranne per i punti di solfuro che per romperli serve l’utilizzo di un agente riducente; poi si ottiene a proteina linearizzata. Una volta tolto l’agente denaturante la proteina grazie alle sue capacità, torna nello stato di struttura terziario perché cerca stabilità. Il Weston Blout si usa in laboratorio per lo studio delle proteine: una volta denaturate si fanno scorrere le proteine su questo gel e poi verranno successivamente trattate.
tipo di struttura secondaria e la terziaria è molto semplice. Tali proteine hanno ruolo strutturale; es.α cheratina molto resistente alla tensione presente in capelli e unghie è costituita da una α elica, destrorsa con super struttura coil- coil che più sono formano i coto filamenti e più coto filamenti danno fibrille. Quindi due catene α cheratine con stessa direzione avvolte l’una sull’altra è visto come super avvolgimento sinistrorso, i gruppi -R si inseriscono vicini alternati in modo regolare. Altro es. collagene resistente alla tensione costituisce tendine cartilagine e matrice osso ed è costituito per 1/3 glicina poi si ha prolina (che forniscono resistenza alla tensione) ed idrossiprolina. L’elica del collagene ha struttura unica destrorsa con tre amminoacidi di residuo sempre coil-coil ma con caratteristiche di struttura terziaria e quaternaria e tale super-avvolgimento dà le caratteristiche della proteina vi sono tre catene polipeptidiche separate super avvolte le une sulle altre in maniera destrorsa. La triplice elica è stabilizzata da legami H tra -COO e -NH ogni tre residui amminoacidici. Dato che prolina ed idrossiprolina, dati gli anelli nella molecola, limitano la rotazione e stabilizzano la proteina dandone le caratteristiche. Più catene di collagene si associandosi tra loro danno fibrille e fibre che aumentano con l’età dando fragilità ai tessuti. Globulari catene polipeptidiche ripetute con forma semi-sferica; hanno più tipi di struttura terziaria e secondaria ed hanno una funzione complessa di enzimi, trasporto o regolazione. Esse hanno due ulteriori livelli d’organizzazione secondaria che determina l’avvolgimento da due o più elementi di struttura secondaria come ansa α-β-α o barile β. Il dominio è una parte stabile che si comporta in modo indipendente che si costituisce da più domini. Es. emoglobina e mioglobina hanno struttura simile ma ruoli diverse; entrambe EME proteine ma Hb trasporta O2 Mb ha funzione dei riserva O2 nel muscolo. Entrambe globulari costituite dal gruppo EME che non è proteico, esso è un composto ciclico e planare con atomo di Fe al centro che lega O2 (in maniera reversibile). Le forme ossigenate sono ossi-Hb e ossi-Mb. Il gruppo EME ha struttura tetrapirrorica e Fe al centro che da sei legami (2 per anelli 1 per proteina e 1 per O2). La differenza tra le due è che Mb ha una sola subunità è un monomero ed Hb è un tetramero quindi 4 subunità. Ogni subunità ha al suo interno un gruppi EME quindi lega 4 O2 e Mb lega solo 1 O2 dato che ha solo un EME che in entrambi i casi è all’interno della proteina perché idrofobico.
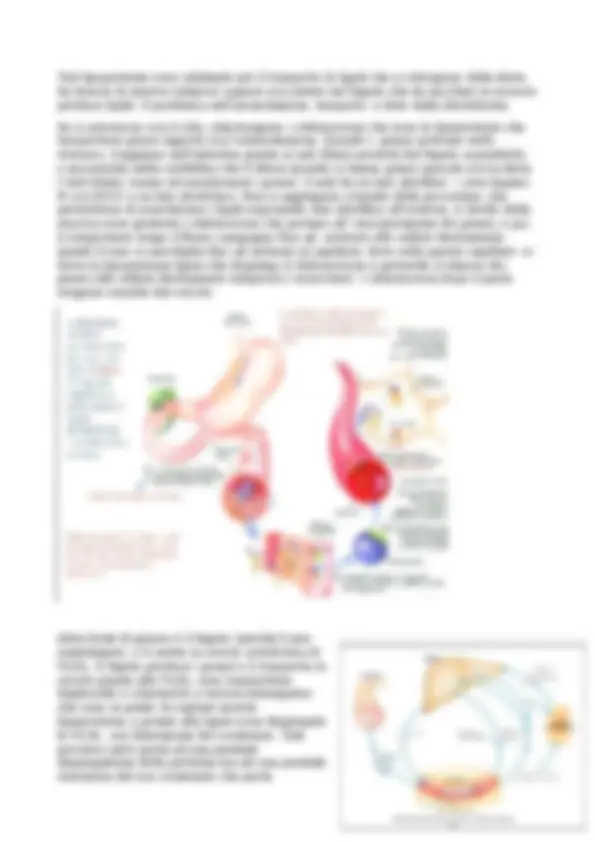
Mioglobina 1 solo dominio 153 aa. con α eliche che formano ripiegamenti. È nel muscolo per riserva O2; essa ha un’affinità per O2 rispetto ad Hb questo perché quest’ultima ha funzione di trasporto per poi cedere O2 ai tessuti mentre Mb ha funzione di riserva quindi per “acchiappare” O2 da Hb deve saperla strappare per questo affinità maggiore per O2. Mb mantiene O2 legato a se anche se la pressione di O2 scende. Emoglobina è formata da 4 subunità ha 4 EME ed è presente negli eritrociti e ne da la colorazione rossa che varia in base alla percentuale di EME legati ad O2 (più rosso se ricche di O2 più scuro se poco O2) ed occupa il 30% del globulo rosso motivo per cui non hanno organelli cellulari e nucleo e fanno fermentazione. Tali cellule hanno vita 120 gg. Le subunità sono unite tra loro per interazioni non covalenti che porta una struttura quasi sferica. Hb è una proteina allosterica cioè è regolata tramite un ligando (O2) definito effettore allosterico, si lega alla proteina in una zona definendone l’attività. Ricordiamo che si legano una molecola di O2 alla volta ma si ha una cooperattività positiva cioè la prima molecola di O2 che si lega ha scarsa affinità che però grazie al legame viene cambiata la struttura conformazionale della molecola promuovendo un’affinità maggiore per la seconda molecola di O2 e cosi via. Osservando il seguente grafico si osserva che nella mioglobina la curva è iperbolica fino a raggiungere il platò anche a basse pressioni di O2 (un parametro è il 50% di saturazione) che si ha a 2,8 mmHg; mentre Hb ha curva sigmoide la saturazione è lenta fino ad aumentare per il discorso della
periferico quando si ha una bassa pressione di O, il PH è basso e aumento pressione CO2. Altre proteine interessanti sono actina e miosina che sono i motori molecolari di origine proteica permettendo il movimento dell’organismo sia come movimento muscolare sia come migrazione degli organelli cellulari lungo i microtubuli ed usano ATP. Esse sono proteine contrattili del muscolo scheletrico e sono organizzate in filamenti che tramite interazioni transitorie che si formano, i filamenti scivolano l’uno sull’altro permettendo la contrazione. Tali proteine costituiscono 80% del muscolo Miosina è una proteina formata da 6 subunità 2 catene pesanti e 4 leggere sono le prime che ne determinano la struttura; le code delle catene pesanti sono due α eliche super avvolte. La catena pesante ha una regione carbossi- terminale e super avvolta del coil-coil con andamento sinistrorso mentre nella regione ammino-termina forma le teste date da domini globulare dove avviene idrolizzato ATP. Le catene leggere sono nella regione ammino-terminale legate alle teste delle catene pesanti. Tali proteine nel muscolo si aggregano in modo ordinato a centinaia formando i filamenti spessi infatti le code interagiscono tra loro formando le strutture dei filamenti spessi e le teste dei domini sono proiettati verso l’esterno in modo giusto ma sfalsato. Actina è un monomero e più monomeri della proteina danno i filamenti di actina, ogni monomero di actina si lega alla testa della miosina. La contrazione muscolare avviene con lo slittamento dei filamenti spessi sui sottili in entrambe le direzioni. La distribuzione è regolare dato che un filamento spesso è circondato da 6 sottili. La testa di miosina è legata ad un monomero di actina, l’ATP si lega alla testa della miosina che provoca la dissociazione dal monomero di actina; L’ATP viene idrolizzato
e diventa ADP mentre è legato alla testa del filamento spesso; ciò porta un cambiamento strutturale della testa di miosina che cambia conformazione: la testa di miosina si attacca nuovamente al filamento sottile di actina rilasciando P cambiando posizione. Il rilascio di ADP comporta il ritorno della testa nella posizione iniziale. Questo è definito come Power Stroke: il rilascio di P genera la forza contrattile che permette lo scorrimento dei filamenti di actina e miosina che ritorna allo stato originario. Considerando che in un filamento spesso ci sono moltissime teste quindi molte proteine eseguono tale slittamento che permette la contrazione, però in un qualsiasi momento circa 1-3% sono legate ai filamenti sottili quindi non c’è mai un momento in cui sono tutte dissociate altrimenti ci sarebbe uno slittamento indietro: ci sono sempre delle teste di miosina legate ai filamenti sottili che impediscono lo slittamento all’indietro. A regolare l’interazione tra actina e miosina ci sono delle proteine che devono avere interazioni che avvengono solo se è implicato il sistema nervoso. Tale compito è principalmente della tropomiosina e della troponina; la prima si lega ai filamenti sottili impedendo il legame della miosina la troponina invece attraverso stimolazione nervosa, fa l’opposto quindi favorisce l’aggancio tra i filamenti cambiando conformazione e facendo esporre le teste di miosina. Quindi la troponina libera i siti di legame della miosina permettendo la contrazione e la tropomiosina li blocca impedendo la contrazione. Enzimi Gli enzimi sono proteine in grado di catalizzare le reazioni quindi rendono le reazioni molto più veloci (senza l’aiuto dell’enzima impiegherebbero troppo per avvenire). L’enzima: non altera la K (costante) di equilibrio, non viene né consumato né modificato in modo permanente, catalizza la reazione più volte partecipando più volte al processo catalitico. Essi sono proteine globulari e il loro peso va da 10000 dalton a oltre un milione di dalton e possono aver e una sola catena polipeptidica con più subunità. Nell’uomo sono stati identificati 20.000 enzimi e il loro suffisso è -ASI. Sono classificati in base alla reazione che catalizzano: Ossidoreduttasi reazioni redox, definiti come deidrogenasi Transferasi trasferimento di gruppi funzionali,