Scarica Espressione del genoma e più Appunti in PDF di Genetica solo su Docsity!
Espressione del Genoma
L’ Espressione del genoma è specifica del tipo di cellula e del suo stato. Essa richiede la sintesi di molecole di RNA ( trascrizione ) che eventualmente dirigeranno la sintesi di proteine ( traduzione ).
- l’RNA polimerasi non ha bisogno di un primer
- l’RNA sintetizzato non rimane appaiato allo stampo
- la trascrizione è meno accurata della replicazione ( 1 mismatch/ 1000 nt, assenza di meccanismi di riparazione).
Le RNA polimerasi
I batteri hanno una unica RNA polimerasi, mentre le cellule eucariotiche ne hanno 3 *: le RNA Pol I, Pol II e Pol III. La RNA Pol II è responsabile della trascrizione della maggior
parte dei geni, inclusi quelli codificanti per proteine, e alcuni piccoli RNA nucleari (U 1 , U 2 ,
U 4 , U 5 ). La RNA Pol I , localizzata nel nucleolo, trascrive il gene per il precursore degli
RNA ribosomiali ( 40 % dei trascritti totali), la RNA Pol III trascrive i geni per i tRNA, alcuni
piccoli RNA nucleari (es. U 6 ) e l’RNA ribosomiale 5 S. La struttura 3 D dell’RNA polimerasi
ricorda una chela (simile alla mano delle DNA polimerasi).
Batteri Archebatteri RNAP I RNAP II RNAP III
! A'/A'' RPA1 RPB1 RPC
!' B RPA2 RPB2 RPC
D RPC5 RPB3 RPC
L RPC9 RPB11 RPC
$ K RPB6 RPB6 RPB
(+ altre 6) (+ altre 9) (+ altre 7) (+ altre 11)
Procarioti Eucarioti
(*) Nella cellula eucariotica esistono altre RNA polimerasi : l’RNA polimerasi mitocondriale (simile a quella del fago T 7 ) e l’RNA polimerasi dei cloroplasti (simile a quella batterica).
Le tre fasi della trascrizione
La trascrizione si sviluppa in tre fasi distinte: inizio , allungamento e terminazione. L’inizio richiede l’intervento di un promotore, ovvero una sequenza di DNA, cui si lega la RNA polimerasi (insieme ad altri fattori di inizio), prima di iniziare la sintesi dell’RNA in direzione 5 ’! 3 ’. Il promotore è monodirezionale dato che sono uno dei due filamenti funge da stampo. Dopo la sintesi di un piccolo frammento di RNA (ca. 10 nt) si passa alla fase di allungamento nella quale la polimerasi si lega ancora più saldamente all’RNA, svolge il DNA davanti, lo riavvolge dietro, stacca la catena di RNA dallo stampo, funziona da correttore di bozze. Quando tutto il gene è stato trascritto, la polimerasi si ferma e rilascia l’RNA prodotto. Separazione di circa 14 basi Formazione del complesso ternario
Le trascrizione nei batteri
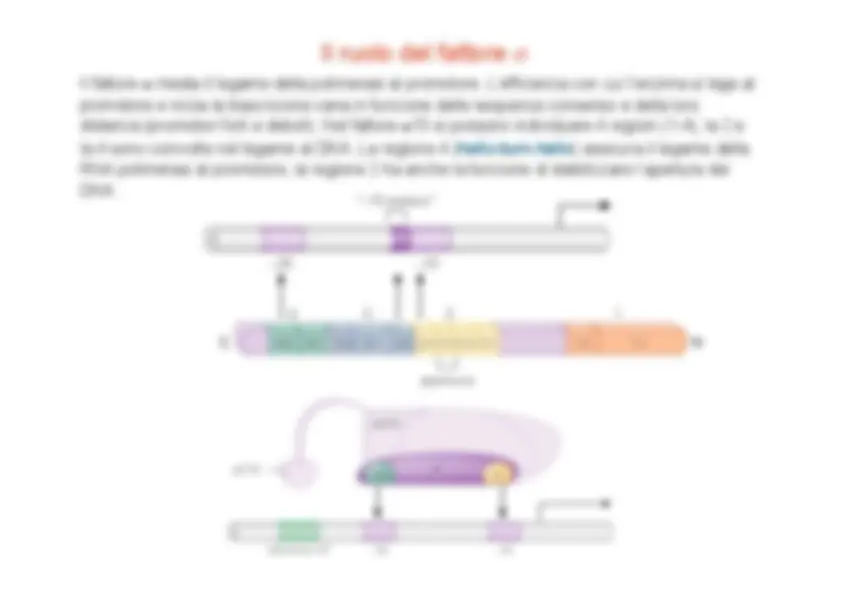
Il nucleo (core) dell’RNA polimerasi ha un’affinità intrinseca per il DNA basata su interazioni fra l’enzima (basico) e il DNA (acido) e si lega stabilmente a siti di legame aspecifici sul DNA. La specificità della RNA polimerasi per il promotore è conferita da un fattore proteico, chiamato sigma , che forma con la proteina core la RNA polimerasi oloenzima. Il fattore sigma predominante è "^70 che lega due sequenze consenso a - 35 e a - 10.
Promotori in E.coli
Filamento coding Filamento stampo TSS
Il ruolo del fattore "
Il fattore " media il legame della polimerasi al promotore. L’efficienza con cui l’enzima si lega al promotore e inizia la trascrizione varia in funzione delle sequenze consenso e della loro distanza (promotori forti e deboli). Nel fattore " 70 si possono individuare 4 regioni ( 1 - 4 ), la 2 e la 4 sono coinvolte nel legame al DNA. La regione 4 ( helix-turn-helix ) assicura il legame della RNA polimerasi al promotore, la regione 2 ha anche la funzione di stabilizzare l’apertura del DNA.
La RNA polimerasi non richiede un primer
La trascrizione non richiede l’intervento di un primer, ma è necessaria una interazione molto specifica con il ribonucleotide di inizio, che normalmente è una A. Dopo l’ingresso del ribonucleotide di inizio nel sito attivo, segue una fase denominata inizio abortivo , nella quale la polimerasi sintetizza brevi frammenti di RNA (< 10 nt) che vengono rilasciati senza che la polimerasi si dissoci dallo stampo, sul quale inizia poi la sintesi di un nuovo frammento. Dopo diversi tentativi, quando la RNA polimerasi riesce a sintetizzare un frammento più lungo di 10 nt, inizia la fase di allungamento (v= 50 nt/s) e si ha poi il distacco del fattore ". La transizione dalla fase di inizio abortivo a quella di allungamento comporta un cambiamento strutturale della regione " 3. 1. Un meccanismo analogo viene ricapitolato dalla RNA polimerasi a singola subunità come la RNA polimerasi T 7. polimerasi T 7
Terminazione della trascrizione
Alcune sequenze, dette terminatori inducono il rilascio della RNA polimerasi e del trascritto. I terminatori batterici sono di due tipi: i) rho-indipendenti , e ii) rho-dipendenti. Nel secondo caso la terminazione richiede il fattore proteico rho (che lega siti specifici detti rut ). I terminatori rho-indipendenti, detti anche terminatori intrinseci, sono costituiti da una corta sequenza palindromica (ca. 20 nt) cui segue una sequenza di 8 coppie A:T. L’interferenza provocata dalla struttura stem-loop unita alla debolezza del legame A:U favorisce il distacco del trascritto dallo stampo. La struttura stem-loop assunta dall’RNA provoca la rottura del complesso di allungamento Sono evidenziate le mutazioni che hanno dimostrato un effetto deleterio sull’attività del terminatore
La trascrizione eucariotica
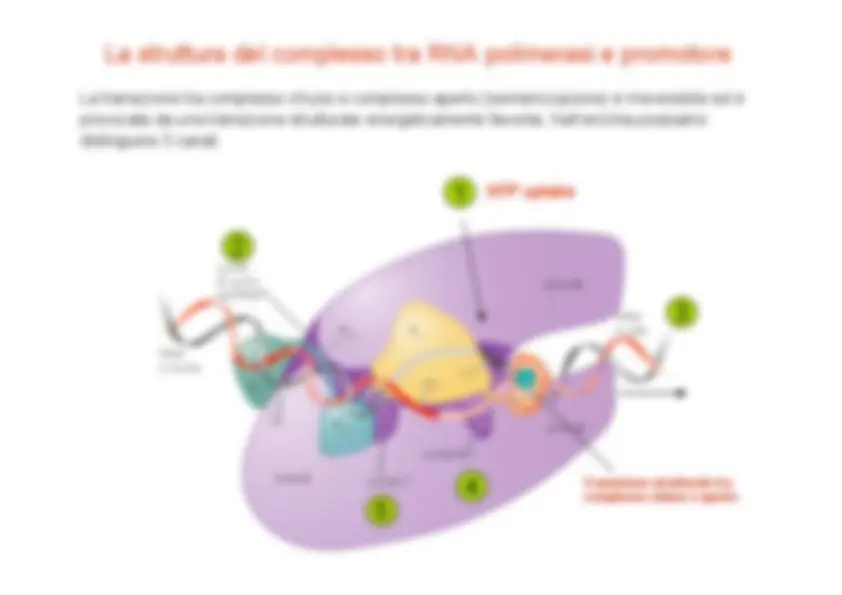
La struttura della RNA polimerasi eucariotica è molto simile a quella batterica. Tuttavia, mentre nei procarioti interviene un solo fattore addizionale ("), negli eucarioti sono richiesti numerosi fattori di inizio addizionali denominati GTF ( fattori generali di trascrizione ). Dato che In vivo, il DNA è avvolto sui nucleosomi, sono richiesti anche ulteriori fattori proteici quali il complesso mediatore , proteine che modificano la cromatina, ecc. Il promotore base (core promoter) riconosciuto dalla RNA Pol II è costituito da circa 40 bp a monte e a valle del sito di inizio e contiene sequenze consenso quali BRE , TATA box , INR e DPE , riconosciute da specifici fattori proteici. Di fatto i promotori eucariotici contengono solo alcuni ( 2 o 3 ) di questi 4 elementi. Altre sequenze regolatorie sono localizzate ad una distanza variabile dal core promoter, principalmente a monte, quali UAS (upstream activating sequences), enhancers , silencers , boundary elements e insulators , che a loro volta sono riconosciuti da proteine regolatorie che attivano o reprimono il processo di trascrizione.
Il complesso di pre-inizio
L’insieme dei GTF, la RNA polimerasi e il promotore formano il complesso di pre-inizio (PIC). Il principale fattore di trascrizione della RNA Pol II è TFIID (TPB + TAFs). Il di legame di TBP al DNA ne induce una marcata distorsione e consente il reclutamento di altri fattori GTF e della stessa RNA polimerasi. L’apertura dei due filamenti a livello del promotore richiede l’intervento di TFIIH e l’idrolisi di ATP. Il passaggio dall’inizio abortivo alla fase di allungamento è segnato dalla fosforilazione della coda CTD (C-terminal domain) dell’enzima su residui di Ser all’interno di ripetizioni eptapeptidiche (PTSPSYS, 52 ripetizioni nei mammiferi). La fosforilazione dell’enzima facilita l’ evasione della polimerasi dal promotore.
Il ruolo degli altri fattori proteici
Il DNA è normalmente impaccato nei nucleosomi e nella cromatina. Pertanto, l’inizio della trascrizione in vivo richiede l’intervento di altri fattori proteici quali il complesso mediatore, proteine regolatrici della trascrizione (attivatori) e enzimi che rimodellano la cromatina.
Allungamento della Trascrizione
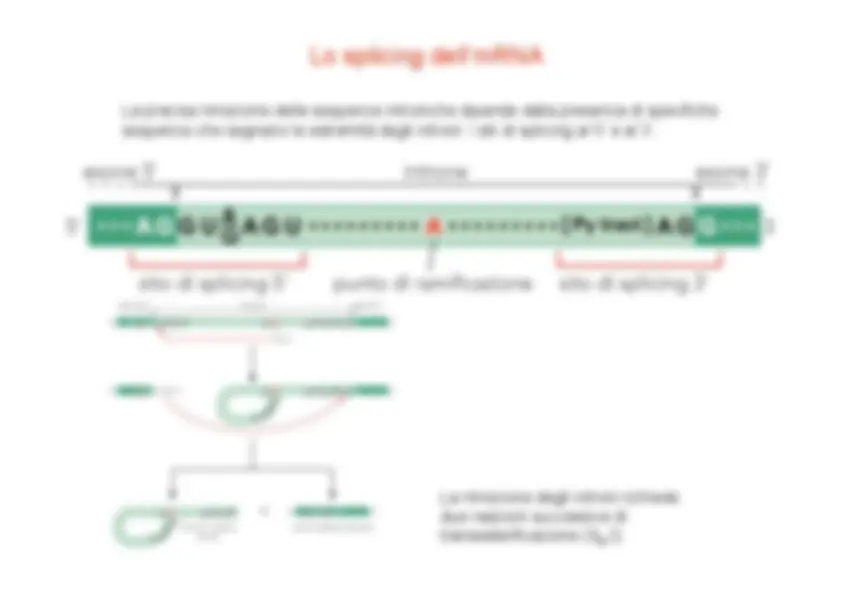
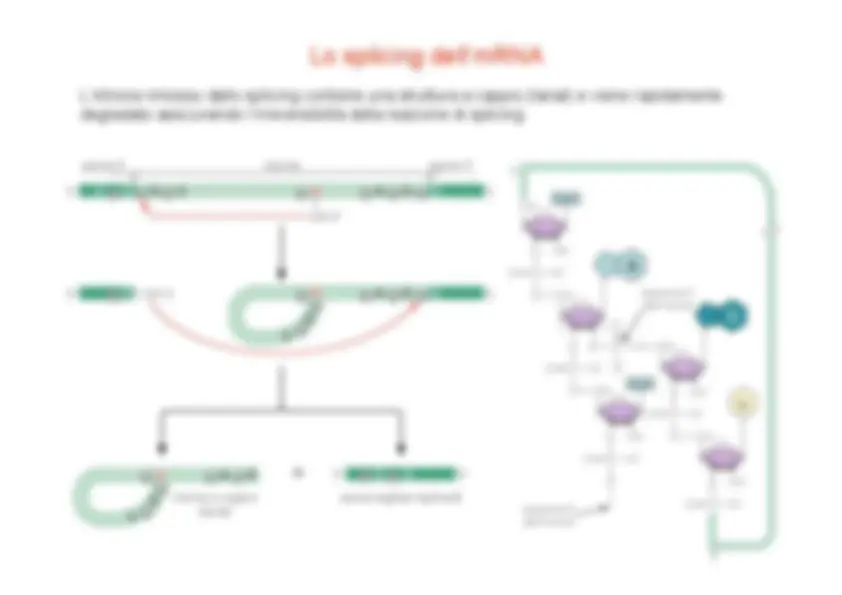
La fosforilazione del CTD, che avviene in concomitanza con l’inizio della fase di allungamento, provoca uno scambio tra i fattori di inizio e quelli necessari per l’allungamento e la maturazione dell’RNA. I fattori di allungamento includono P-TEFb (fosforilazione S 2 ), hSPT 5 , TAT-SF 1 , TFIIS (correzione di bozze) Fattori di splicing Fattori di capping
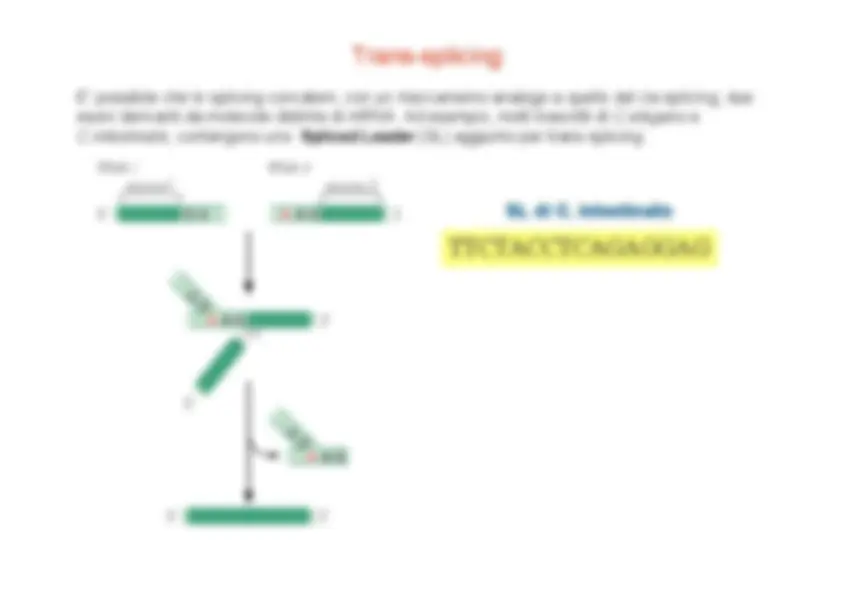
Maturazione dell’mRNA -
aggiunta della coda di polyA
Quando la polimerasi raggiunge il segnale di poliadenilazione, in prossimità della fine del gene, vengono reclutati dal CTD due fattori proteici: CPSF (cleavage and polyadenylation specificity factor) e CstF (cleavage stimulation factor) che a loro volta reclutano altre proteine che effettuano il taglio dell’RNA e l’aggiunta della coda di polyA. L’mRNA così maturo viene rilasciato dalla polimerasi, che per un tratto continua la sintesi di RNA sullo stampo, e quindi trasferito dal nucleo al citoplasma dove sarà tradotto.
RNA Polimerasi I
Il promotore della RNA Pol I è dedicato alla espressione di un singolo gene (presente nel genoma in copie multiple in tandem): il precursore degli rRNA, il cui livello di espressione è molto più alto di quello di tutti gli altri geni. B A Il promotore di Pol I ha una struttura bipartita: il core promoter e l’UCE (upstream control element). L’UCE lega due fattori: UBF e SL 1 (TBP+TAFs), che a loro volta stimolano la trascrizione reclutando la Pol I sul core promoter,