Scarica Controllo del ciclo cellulare: Sintesi e duplicazione del DNA - Prof. Papa e più Dispense in PDF di Citologia solo su Docsity!
Funzioni del nucleo
Ciclo Cellulare
Nei tessuti a elevato ricambio cellulare le cellule hanno vita limitata nel tempo, al termine della
quale si dividono in due cellule figlie che hanno le stesse caratteristiche della cellula madre. Il peri-
odo che intercorre fra l’origine di una cellula da una divisione e il momento in cui questa, a sua vol-
ta, si divide costituisce il ciclo cellulare.
TEORIA CELLULARE
Tutti gli organismi consistono di una o più cellule;
La cellula è l’unità di base della struttura di tutti gli organismi;
Tutte le cellule originano da cellule preesistenti;
Le cellule generano da altre cellule tramite la divisione di cellule già esistenti;
Una cellula duplica il suo DNA e poi si divide in due;
Il ciclo di duplicazione di divisione è detto ciclo cellulare ed è alla base della riproduzione di tutti
gli esseri viventi.
DURATA DEL CICLO DI DIVISIONE IN ALCUNE CELLULE EUCARIOTICHE
- Cellule di embrione iniziale di rana: 39 minuti.
- Cellule di lievito: 1.5/3 ore.
- Cellule dell'epitelio intestinale di mammifero: circa 12 ore
- Fibroblasti di mammifero in coltura: circa 20 ore.
- Cellule del fegato umano: circa 1 anno.
SISTEMA DI CONTROLLO DEL CICLO CELLULARE
Gli eventi del ciclo cellulare avvengono secondo un ordine di eventi stabilito;
Ogni evento deve essere concluso prima che inizi il successivo;
Se la sintesi del DNA rallenta anche la mitosi e la citochinesi devono rallentare;
Se il DNA è danneggiato il ciclo deve arrestarsi, cosicché la cellula possa riparare il danno.
Fasi del ciclo cellulare
Il ciclo comprende quattro fasi, nel corso delle quali una cellula si sviluppa, replica i suoi costituenti
molecolari e, infine, si divide in due cellule figlie. Il ciclo vitale della cellula comporta la regolare
successione delle fasi G 1 , S , G 2 e M. L’intero ciclo ha una durata compresa fra alcune ore a
qualche giorno. Le fasi G 1 , S, G 2 costituiscono il periodo di intercinesi , caratterizzato dalla pre-
senza di un compartimento nucleare delimitato e ben individuabile. La fase G 1 è contraddistinta da
attivi fenomeni trascrizionali e da conseguenti sintesi proteiche; la cellula neoprodotta deve ac-
crescersi e duplicare i suoi organuli prima di potersi a sua volta dividere. In fase S, la sintesi di
nuovo DNA è accompagnata da sintesi di istoni che passano rapidamente nel nucleo andando a
costruire (con il DNA) nuova cromatina. Nella fase G 2 , il citoplasma sintetizza nuove proteine, sia
di membrana sia destinate alla costruzione dell’apparato mitotico, rimane essenzialmente un peri-
odo di preparazione alla successiva divisione.
Le cellule che vanno incontro a un profondo differenziamento, costruendo nuovi organuli o apparati
e ingrandendosi (come è il caso delle cellule nervose o muscolari), di solito sono incapaci di di-
vidersi e formano i tessuti perenni o stabili. In questo caso il ciclo si blocca in G 1 , senza che vi sia
duplicazione del materiale genetico (quando G 1 si prolunga nel tempo indefinitamente prende il
nome di fase G 0 ).
Controllo del ciclo cellulare
I momenti chiave nel controllo del ciclo cellulare sono:
Passaggio dalla fase G 1 alla fase S , che nei lieviti sembra essere il più importante ed è chiama-
to start e nei mammiferi punto di restrizione ;
Passaggio dalla fase G 2 alla fase M.
Le molecole che controllano i veri eventi del ciclo cellulare sono proteinchinasi ciclino-dipendenti
dette Cdk , che fosforilano, attivandole, le varie proteine che danno inizio e regolano i processi
principali delle varie fasi, come la duplicazione del DNA, la condensazione dei cromosomi, la dis-
gregazione dell’involucro nucleare alla mitosi ecc.
Le Cdk sono sempre presenti nella cellula, ma funzionano solo se sono attivate dalle proteine
chiamate cicline delle quali ne esistono di versi tipi che si uniscono alle Cdk.
I tre complessi principali di ciclina-Cdk sono:
- G 1 /S-Cdk , formato dalla ciclica G 1 /S e dalla proteinchinasi Cdk2;
- S-Cdk , formato dalla ciclica S e da Cdk
- M-Cdk , formato dalla ciclica M e dalla proteinchinasi Cdk1;
4*. G 1 -Cdk, formato dalla ciclica G 1 e da due proteinchinasi, Cdk4 e Cdk6, questo quarto comp-
lesso esiste nella maggior parte delle cellule.
Quest’ultimo complesso è il primo che entra in funzione nel ciclo cellulare e attiva la trascrizione
dei geni per la ciclica G 1 /S e la sua successiva sintesi. Questa ciclica si unisce alla proteinchinasi
Cdk2 formando il primo complesso che è responsabile dell’inizio degli eventi che porteranno alla
duplicazione del DNA.
Negli eucarioti la duplicazione comincia a livello di determinate sequenze, chiamate siti di origine
della replicazione, alle quali si lega un gruppo di proteine che costruiscono il cosiddetto complesso
di riconoscimento dell’origine. A questo complesso si unisce una proteina regolatrice detta Cdc6 e,
successivamente alcune DNA elicasi. Si forma così il complesso prereplicativo che però si attiva
solo se stimolato dal secondo complesso. Il complesso G 1 /S-Cdk (primo) induce la sintesi delle
cicline S e la formazione del complesso S-Cdk (secondo), che dà inizio alla duplicazione del DNA
attivando il complesso prereplicativo. Dopo che la duplicazione è iniziata S-Cdk (secondo) fa dis-
sociare Cdc6 e le DNA elicasi dal complesso prereplicativo impedendo l’inizio di un’ulteriore repli-
cazione del DNA. Il secondo complesso rimane in funzione per tutta la fase G 2 e per il periodo in-
iziale della mitosi. Nel frattempo, nella fase G 2 viene sintetizzata e si accumula la ciclica M che,
unendosi a Cdk1, forma il complesso M-Cdk o MPF (fattore per la promozione della mitosi).
Questo complesso, inizialmente non attivo per la presenza di gruppi fosfato ad azione inibitoria
nella molecola della proteinchinasi, viene attivato da una fosfatasi, chiamata Cdc25, che rimuove i
suddetti gruppi fosfato.
La mitosi, però, non inizia prima che la duplicazione di tutto il DNA sosia completata in modo cor-
retto. Ciò è controllato dal cosiddetto punto di controllo della replicazione.
TRASCRIZIONE E MATURAZIONE DEGLI mRNA (MESSAGGERO)
Trascrizione
Durante la trascrizione, la RNA polimerasi "copia" l'informazione contenuta in un gene sul DNA in
una molecola di mRNA. Questo processo è simile nei procarioti e negli eucarioti. Una differenza
notevole, invece, è che l'RNA polimerasi degli eucarioti si associa con gli enzimi di verifica dell'm-
RNA durante la trascrizione, in modo da far procedere velocemente la modificazione dopo l'inizio
della trascrizione. Il prodotto non modificato o parzialmente modificato è chiamato pre-mRNA, che
una volta modificato prende il nome di RNA maturo.
Modificazioni del pre-mRNA eucariotico
La maturazione dell'mRNA differisce molto tra gli eucarioti ed i procarioti. L'mRNA procariotico è
già maturo dopo la trascrizione e non richiede di essere controllato, tranne che in alcuni casi. Il
pre-RNA eucariotico invece richiede una lunga verifica.
Maturazione dell'mRNA
La maturazione dell'mRNA è una serie di processi chimici che trasformano una molecola di pre-
mRNA (trascritto primario) in mRNA. Le tappe della maturazione sono:
- lo splicing ; consiste nell’eliminaione dei tratti intronici e nell’unione dei tratti esonici;
- il capping (rivestimento in 5’);
- la poliadenilazione (aggiunta di una sequenza poliadenilica all’estremità 3’).
Le molecole di snRNA prendono parte alla maturazione.
RNA editing
L'editing è la modifica della composizione nucleotidica dell'mRNA. L'editing crea un codone di stop
prematuro, che sottoposto a traduzione produce una proteina più corta.
Trasporto
Un'altra differenza tra procarioti ed eucarioti è il trasporto dell'mRNA. Poiché negli eucarioti
trascrizione e sintesi proteica avvengono in compartimenti separati, l'mRNA eucariotico deve es-
sere trasportato dal nucleo cellulare al citoplasma. Gli mRNA maturi vengono riconosciuti mediante
le loro modificazioni e poi esportati tramite il poro nucleare, trasporto mediato da proteine tra
cui NXF1.
Traduzione (sintesi proteica)
Poiché l'mRNA procariotico non necessita di essere trasportato, la sintesi proteica (avviene nel
citoplasma, operata dai ribosomi) può iniziare immediatamente dopo l'inizio della trascrizione.
Quindi si può dire che la traduzione procariotica è accoppiata alla trascrizione ed avviene co-
trascrizionalmente.
L'mRNA eucariotico che è stato modificato e trasportato al citoplasma (detto a questo punto RNA
maturo), può poi essere tradotto dal codice genetico. La traduzione può avvenire in ribosomi liberi
nel citoplasma oppure nel reticolo endoplasmatico mediante una signal recognition particle (re-
gione di riconoscimento del segnale). Quindi, diversamente dai procarioti, la traduzione negli eu-
carioti non è direttamente accoppiata alla trascrizione.
Degradazione
Dopo un certo lasso di tempo il messaggio si degrada nei suoi componenti nucleotidici, di solito
con l'assistenza dell'mRNasi. Grazie al controllo che subisce, l'mRNA eucariotico è generalmente
più stabile di quello procariotico. Per essere degradato, l'mRNA eucariotico, necessita di fattori che
lo destabilizzino: tra questi fattori destabilizzanti troviamo gli elementi ARE (elementi ricchi di AU) e
gli elementi IREs (Internal Ribosome Entry site). Questi elementi si attivano quando si dissociano
dalle proteine alle quali sono legati, attivando anche endonucleasi che catalizzano la degradazione
dell’mRNA.
NUCLEOLO E SINTESI DEGLI RNA RIBOSOMIALI
Il nucleolo è l’equivalente morfologico dell’attività di trascrizione e di maturazione dei geni medi-
ante ripetuti che codificano per gli rRNA, oltre che dell’assemblaggio di questi ultimi con proteine
per formare i ribosomi.
Il nucleolo si colora con coloranti sia basici, perché ricco di RNA, sia acidi, per la contemporanea
presenza di numerose proteine.
Struttura del nucleolo
Il nucleolo mostra tre distinte porzioni: il centro fibrillare , la componente fibrillare densa e la com-
ponente granulare.
Nella componente fibrillare densa e nel centro fibrillare si trova il DNA che codifica per gli rRNA e
costituisce il cosiddetto organizzatore nucleolare.
La componente granulare è costituita dai prodotti della maturazione dell’rRNA sotto forma di
granuli nucleoproteici, precursori delle subunità ribosomiali.
Meno costante è la presenza di una terza componente detta amorfa , perché appare priva di strut-
tura. Questa componente, essenzialmente proteica, potrebbe corrispondere al nucleoscheletro
(matrice nucleolare).
Il nucleolo si dissolve all’inizio della mitosi, durante la provasse, e si riforma al termine della divi-
sione.
Processing dei trascritti ribosomiali
Ciascun gene è formato dalle sequenze codificanti gli rRNA 18S, 5,8S e 28S, separate da sequen-
ze spacer interagenici producendo lunghe molecole di RNA di 45S, precursore dell’rRNA.
La molecola di RNA 45S appena trascritta viene unita a varie proteine, formando un complesso
ribonucleoproteico di 80S, e quindi si stacca dal gene.
Le varie fasi della maturazione
dell’ rRNA e dei ribosomi sono
controllate da centinaia di parti-
celle costituite da proteine e da
snoRNA che sono in grado di
guidare il corretto posizionamento
e l’attività degli enzimi che modifi-
cano gli rRNA. Queste particelle di
snoRNP si uniscono agli RNA 45S
ancora in fase di trascrizione
( knob ) e ciò fa pensare che le
prime fasi della maturazione in-
i z i a n o a n c o r p r i m a c h e l a
trascrizione sia completata.
Altre funzioni del nucleolo
Regolazione genica
Negli eucarioti il controllo dell’espressione genica opera a vari livelli: trascrizionale, post-
trascrizonale e tradizionale. Inoltre in cellule diverse alcuni geni siano stabilmente repressi e ciò è
alla base del differenziamento.
REGOLAZIONE GENICA NEI PROCARIOTI
Nei procarioti i geni che controllano un’unica funzione metabolica formano un’unica unità
trascrizionale continua, chiamata operone , a
monte della quale si trova un unico promotore.
Esistono due meccanismi di regolazione genica
nei procarioti:
- Induzione enzimatica , riguarda di solito enzimi
addetti al catabolismo di metaboliti che il batterio
trova in abbondanza nel substrato su cui vive;
- Repressione enzimatica , riguarda gli enzimi
addetti alla sintesi (anabolismo) di metaboliti
normalmente presenti nel citoplasma batterico.
Questi meccanismi di controllo sono essenzial-
mente basati sul principio del risparmio: essi per-
mettono alla semplice, ma efficiente cellula pro-
cariotica la sintesi di quelle proteine di cui essa ha
effettivamente bisogno in un dato momento e in
un particolare ambiente.
REGOLAZIONE GENICA NEGLI EUCARIOTI
La caratteristica peculiare della regolazione genica degli eucarioti è la trascrizione differenziale.
Regolazione a livello trascrizionale
La regolazione genica a livello trascrizionale dipende dalla presenza a monte del gene di sequen-
ze regolative, che possono trovarsi anche a migliaia di basi dal gene stesso, quali intensificato (o
potenziatori), che attivavano o intensificano la trascrizione, e silenziatori , che invece la reprimono.
Queste sequenze sono riconosciute in modo specifico dalle proteine regolatrici del grippo dei fat-
tori di trascrizione, che possono essere attivatori o repressori della trascrizione.
Regolazione genica a livello post-trascrizionale
Uno dei più noti meccanismi di controllo post-trascrizionale è quello dello splicing alternativo , che
agisce durante la maturazione dell’hnRNA. Con questo meccanismo, a partire da un unico hnRNA,
mediante le combinazioni diverse di vari esoni, si possono ottenere mRNA e proteine differenti.
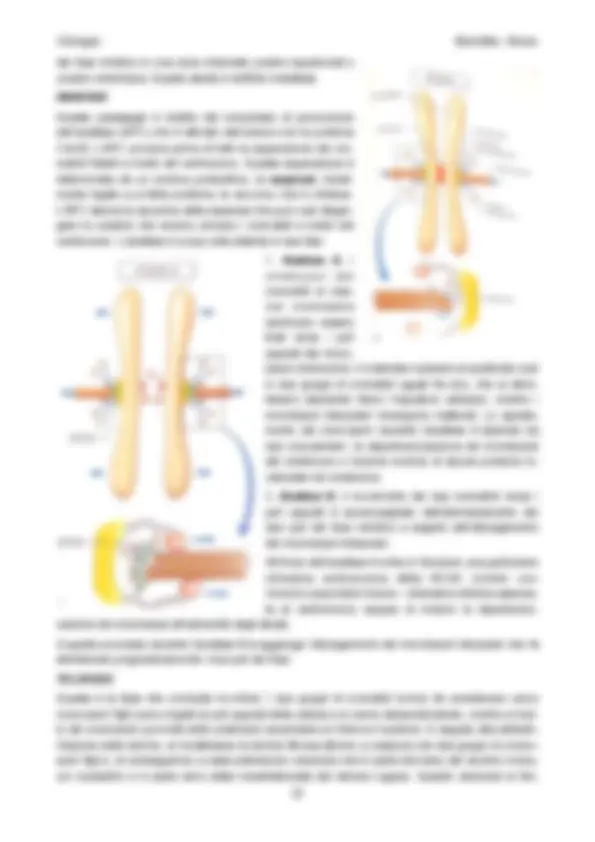
Fase S
Nella fase S, o sintesi, avviene la duplicazione del DNA.
Prima di questa fase i cromosomi sono costituiti da una doppia elica di DNA e la quantità di DNA
per nucleo corrisponde a un valore definito 2C; dopo la duplicazione i cromosomi sono costituiti da
due doppie eliche di DNA e la quantità di DNA corrisponde a un valore di 4C.
Meccanismo di duplicazione del DNA
NUCLEOSIDI 3-P
Ognuno dei filamenti di DNA può fare da stampo. Il nucleotide A
si lega preferenzialmente con T, il nucleotide G si lega pref-
erenzialmente con C. Per questo ogni filamento può fungere da
stampo per la formazione del filamento complementare.
Il DNA fa da stampo per duplicare se stesso.
La replicazione del DNA è detta semiconservativa
Il genoma batterico ha una sola origine di replicazione, il genoma umano ne ha circa 10000.
Le posizioni in cui il DNA incomincia ad aprirsi si chiamano origini di replicazione e si distinguono
per una particolare sequenza (A-T). Il processo di replicazione ha inizio quando uno specifico
gruppo di proteine iniziatrici si lega a questa sequenza e, utilizzando energia derivata dall’ATP,
svolge la doppia elica. Interverranno diverse proteine deputate a duplicare il DNA ciascuna con
una funzione.
Il genoma batterico ha una sola origine di replicazione, il genoma umano me ha circa 1000.
Replicazione del DNA circolare
Nella cellula eucariotica
E’ importante che il DNA replicato ad un’origine di
replicazione, in quel punto non replichi più.
LA DNA POLIMERASI CATALIZZA L’ALLUNGA-
MENTO DEI FILAMENTI DI DNA
Il DNA è sintetizzato in direzione 5’ 3’. La DNA
polimerasi aggiunge un deossiribonucleotide al-
l’estremità 3’ di una catena polinucleotidica, quindi la
nuova catena viene sintetizzata nella direzione che va
dal 5’ verso il 3’. I nucleotidi sono nucleosidi trifosfati.
La rottura del legame fosfoanidride rende disponibile
una grande quantità di energia per alimentare ener-
geticamente la reazione di polimerizzazione.
L’attività di autocorrezione spiega perché l’attività di sintesi
della DNA polimerasi procede solo in direzione 5’-3’
❖ La DNA polimerasi è incapace di dare inizio a una catena polinucleotidica nuova;
❖ Un enzima chiamato primasi sintetizza brevi tratti (10 nucleotidi) di RNA usando DNA come
stampo;
❖ I tratti di RNA fungono da innesco (primer) per la sintesi del DNA;
❖ Al filamento guida, l’innesco serve solo per cominciare la sintesi all’origine di replicazione;
❖ Sul filamento lento sono continuamente necessari degli inneschi.
La DNAprimasi
Inneschi di RNA sul filamento lento
Sul filamento lento sono continuamente necessari
nuovi inneschi.
- Alla forcella replicativa si espone un nuovo tratto
di basi libere sulle quali vengono sintetizzati nuovi
inneschi di RNA;
- La DNA polimerasi aggiunge un deossiribonu-
cleotide all’estremità 3’ di questo innesco, dando
inizio ad un filamento di DNA e allungandolo finchè
non si imbatte nell’innesco a RNA successivo;
- Gli inneschi a RNA vengono eliminati da una
nucleasi;
- Una DNA polimerasi detta polimerasi riparativa
sostituisce il DNA all’RNA, usando come innesco il
frammento di Okazaki;
- La DNA ligasi unisce il fosfato terminale al 5’ di
un frammento con l’ossidrile 3’ del seguente.
LE TOPOISOMERASI
Riparazione del DNA
Variazioni permanenti del DNA: mutazioni.
ANEMIA FALCIFORME
Sostituzione dell’adenina con timina:
cambiamento del 6° aminoacido
(l’acido glutammico è sostituito da
una valina) della catena β dell'emo-
globina.
Anemia cronica. I globuli rossi che
contengono emoglobina S vengono
distrutti molto prima di quelli normali.
Mentre la vita media di un globulo
rosso normale è di circa 120 giorni,
quella dei globuli "falciformi " non su-
pera in genere i 20 giorni. Questo
causa una penuria di globuli rossi, e
quindi anemia (con senso continuo di
fatica, pallore, "fiato corto" etc..)
Infezioni. I globuli rossi vengono
distrutti soprattutto nella milza, che
può così risultare danneggiata. La
milza ha un ruolo importante anche nel proteggere l'organismo dalle infezioni, ed è per questo
che per le persone affette da AF (specialmente i bambini) alcune infezioni batteriche possono
risultare molto pericolose.
Dolori alle mani e ai piedi (dactilite -hand-foot syndrome-). Dolori e tumefazioni del dorso delle
mani e dei piedi (dactilite) sono causati dall'occlusione dei capillari da parte dei globuli rossi al-
terati. Spesso questo è uno dei primi sintomi dell'AF nei bambini.
Dolori improvvisi. L'occlusione dei capillari può avvenire in modo imprevedibile in qualunque
parte del corpo, bloccando l'afflusso di sangue agli organi colpiti. La frequenza di queste "crisi" è
molto variabile. I dolori sono la manifestazione più frequente dell'AF.
Sindrome polmonare acuta. E' una delle complicazioni più pericolose dell'AF, simile ad una pol-
monite, causata dall'infiltrazione di globuli rossi nei polmoni o da infezioni polmonari. Nel peg-
giore dei casi, questa manifestazione può anche risultare letale.
Ictus. E' un rischio possibile, a causa dell' occlusione dei capillari che portano il sangue al cervel-
lo.
Proteggere le cellule germinali dalle mutazioni è importante perché queste possono trasmettere i
geni mutati alla generazione successiva. Proteggere le cellule somatiche dalle mutazioni è impor-
tante perché possono dare origine a varianti in grado di crescere e dividersi senza regole a danno
di altre cellule: cancro.
Il DNA viene danneggiato
Formazione di dimeri di pirimidina(indotti dalla luce ultravioletta)
RUOLO DELLA PROTEINA P53 NELLA RISPOSTA AL DAN-
NO DEL DNA
●La proteina p53 lega il DNA è agisce da fattore trascrizione le
attivando la trascrizione del gene che codifica per le proteina
p21, un inibitore della Cdk;
●Se il danno al DNA non può essere riparato, la p53 attiva la
trascrizione di geni che codificano per un gruppo di proteine
coinvolte nell’induzione della morte cellulare per apoptosi.
La sopravvivenza, la crescita è la divi-
sione delle cellule sono influenzate da
molecole segnale
I fattori di sopravvivenza promuovono la sopravvivenza delle
cellule inibendo l’apoptosi;
I mitogeni stimolano la divisione cellulare;
I fattori di crescita stimolano l’accrescimento cellulare promuovendo la sintesi delle proteine.
FATTORI DI CRESCITA E PROLIFERAZIONE CELLULARE
Negli organismi pluricellulari non sono sufficienti i nutrienti presenti nel liquido extracellulare per
indurre la crescita e la divisione delle cellule.
Sono necessarie:
● Proteine di segnalazione extracellulari, chiamati fattori di crescita.
● Mitogeni : stimolano la cellula ad entrare nella fase S del ciclo cellulare e quindi nella fase
G2 e in mitosi.
MITOGENI
Stimolano la divisione cellulare.
Nella maggior parte dei casi allentano i freni molecolari che bloccano la transizione dalla fase G
alla fase S del ciclo cellulare.
● PDGF (platelet derived growth factor), fattore di crescita piastrinico.
Le piastrine nel coagulo producono il fattore di crescita piastrinico il quale legherà a recettori
tirosina-chinasi delle cellule presenti nella lesione inducendo la loro proliferazione.
● Fattore di crescita epatico , quando una parte del fegato è asportata chirurgicamente.
Controllo del ciclo cellulare
Il ciclo cellulare può essere regolato da segnali extracellulari come per esempio i fattori di crescita
che innescano tipiche vie di segnalazione che portano alla stimolazione della
proliferazione cellulare. Queste vie di segnalazione sono costituite da cascate di segnali intracellu-
lari che conducono a fattori di trascrizione che modulano l’espressione genica.
Fattori di crescita
Stimolano l’aumento della massa totale delle cellule.
Fase G
La fase G 2 è il periodo preparatorio per la divisione cellulare.
In questo periodo si ha la sintesi di gran parte delle strutture microtubulari che formeranno l’appa-
rato mitotico. Di questo apparato, alcune parti preesistono e sono state duplicate in S (centrioli);
altre sono fabbricate in G 2 sotto forma di proteine globulari, che poi polimerizzano in microtubuli.
Sempre in G 2 sono sintetizzati vari componenti di membrana che sono utilizzati per la costruzione
di parte delle nuove membrane plasmatiche delle due cellule figlie.
Nella fase G 2 avviene la sintesi dell’estone centromerico CENP-A, della ciclina mitotica (o ciclina B
o M) e la sua unione alla Cdk per formare l’MPF che induce la cellula a entrare in mitosi.
Fase M
La mitosi è il meccanismo di divisione che le cellule usano comunemente per riprodursi. Nella mi-
tosi è essenziale che si attui una serie di processi che consentono di trasmettere alle due cellule
figlie un patrimonio genetico identico a quello della cellula madre e che vi sia una ripartizione equi-
librata anche di tutti gli altri componenti cellulari.
Fasi della mitosi