!
APPRENDIMENTO E MEMORIA
Fattori genetici e ambientali
Contribuiscono alla capacità di mantenere una performance in un compito nel tempo, o acquisire una memoria e applicarla nel tempo.
L’apprendimento è il meccanismo attraverso cui delle esperienze “se ne fa tesoro”e la prossima volta in cui si è sottoposti allo stesso stimolo di
applicherà un adattamento più efficace a quel contesto;
La memoria è quel processo attraverso cui le conoscenze e esperienze si una situazione ambientale vengono codificate in strutture, conservate e
eventualmente richiamate all’utilizzo.
L’informazione viene trasformata in un CODICE per il suo mantenimento a lungo termine in vista di un possibile richiamo.
Neurologia classica
1820: Gall elaborò una teoria secondo cui il cranio potesse variare, vi fossero differenze da individuo a individuo e che i “bozzi” fossero dovuti •
a una espansione di un’area cerebrale sottostante; le funzioni che Gall cercava sulla superificie erano la moralità, l’affidabilità,…
1860 circa: Broca costruì una mappa delle funzioni sulla superficie della corteccia e in questo periodo ci si iniziò a interrogare se anche la •
memoria fosse localizzabile e se avesse un suo sito
Fino alla metà del XX secolo: convinzione che la memoria non fosse una funzione indipendente, ma trasversale, dipendente dalle altre funzioni, •
non localizzabile e quindi non separata dalle altre;
Inizio 900’: vari tentativi per localizzarla •
Metà 900’: si è ritenuta la memoria non localizzabile •
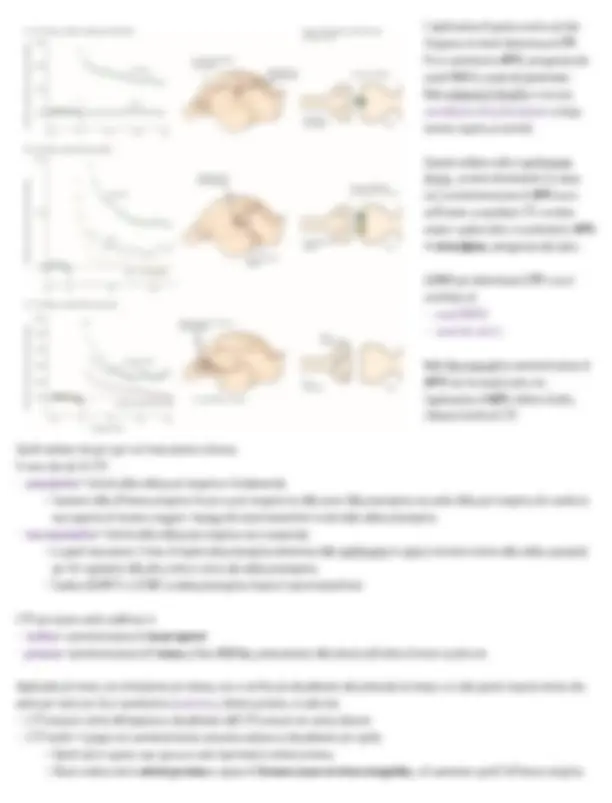
Lashley
Fece vari esperimenti per localizzare la memoria nel cervello del ratto in un labirinto: il ratto poteva circolare liberamente per memorizzare la
struttura;
esperimento che anticipò ciò che Tulving in seguito chiamò mappa mentale •
= memoria/ rappresentazione spaziale del labirinto che presentava permanenza nel tempo ◦
se si toglieva il ratto dal labirinto e lo si rimetteva dopo una settimana sembrava che esso sapesse dove andare, con velocità di risoluzione > •
dimostra una chiara curva di apprendimento con riduzione del tempo ◦
si concluse che vi era una MEMORIA ‣
A questo punto si applicò una manipolazione comportamentale per misurare il grado di apprendimento
= quanto tempo impiegava il ratto a fare il labirinto la seconda volta
poi si fecero dei lavori per individuare l’area che, se lesionata, portava a perdita di memoria, costruiva una mappa della aree e del loro impatto •
sulla performance una volta rimosse
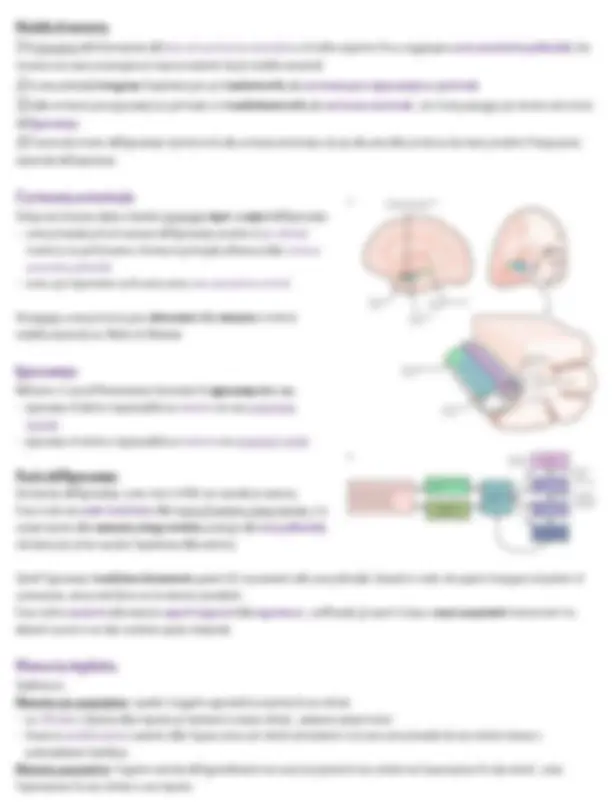
Penfield (10 anni dopo)
Osservò che alcuni siti dei lobi temporali se stimolati , evocano nel paziente esperienze pregresse.
Paziente HM
Paziente la cui osservazione fece cambiare completamente idea sulla memoria:
gli venne asportato l’ippocampo bilaterale, l’amigdala è una porzione minore della porzione basale dei lobi temporali •
Venne fatto per RIMUOVERE un FOCOLAIO EPILETTICO FARMACORESISTENTE ◦
fu rilevata una amnesia particolare in cui: •
La memoria antecedente era conservata 1.
Normale padronanza de linguaggio 2.
Quoziente intellettivo invariato 3.
Mancava la capacità di fissare nuovi ricordi a lungo termine 4.
quindi questo paziente dimostrò che vi è una dissociazione tra processo di mantenimento memoria e costruzione memoria a lungo termine;
Inoltre vi è una dissociazione del mantenimento mantenimento a breve di un contenuto informativo e del processo di storage