Scarica Microbiologia Generale: La Cellula Batterica - Struttura e Funzioni e più Dispense in PDF di Microbiologia E Batteriologia solo su Docsity!
MICROBIOLOGIA
[GENERALE]
Testi di riferimento:
MICROBIOLOGIA MEDICA- Sherris J.C.- (5a
ediz. 2011)- I ediz Italiana - 2013
PRINCIPI DI MICROBIOLOGIA MEDICA –
M. La Placa – 14aed. Ed. Esculapio
– EdiSES - 2014
Appunti Medicina/Farmacia/CTF
MICROBIOLOGIA
[GENERALE]
Argomenti trattati:
1. CELLULA BATTERICA
2. LA SPORA
3. METABOLISMO BATTERICO
4. REPLICAZIONE
5. COLTIVAZIONE BATTERICA
6. TERAPIA ANTIMICROBICA
7. FARMACO-RESISTENZA
8. PATOGENESI
9. TOSSINE
10.MICETI
a. MICETI LIEVITIFORMI
11.VIRUS
12.REPLICAZIONE VIRALE
a. FASI DEL CICLO REPLICATIVO VIRALE
13.METODI DI COLTIVAZIONE VIRALE
14.FASI DELLA PATOGENESI VIRALE
15.FARMACI ANTIVIRALI
16.PREVENZIONE
17.DIAGNOSI MICROBIOLOGICA
STORIA
A Leeuwenhoek, ottico e naturalista olandese, si deve la costruzione del primo microscopio.
Questo era semplicemente una lente di ingrandimento vicino ad una piccola vite che permetteva di mettere a fuoco ciò che si trovava sul vetrino.
Francesco Redi, nel 1600, smontò l’assunto, sino ad allora radicato nella comunità scientifica, della generazione spontanea , secondo cui, da un pezzo di carne, a temperatura ambiente, si sviluppava una vita (le larve), e la carne si avviava così al processo di putrefazione. Grazie al suo esperimento, Redi dimostrò che in realtà non si trattava di generazione spontanea, ma di cattiva conservazione, che permetteva, in questo modo, il depositarsi di insetti o microbi, in particolare, che avviavano la carne al processo di putrefazione. In particolare, Redi, pose la carne in due differenti contenitori: a. Uno chiuso ermeticamente grazie a delle garze; b. Uno lasciato aperto.
I risultati evidenziarono come l’alimento sigillato, aveva una più lunga conservazione.
Pasteur, nel 1860 , intuì che l’innalzamento della temperatura , causava una riduzione della carica microbica. Semplicemente, intuì che, del brodo esposto all’aria aperta andava subito incontro a contaminazione, ma se questo veniva posto in un contenitore con collo ripiegato, e successivamente bollito, veniva mantenuto sterile. Il collo ripiegato dell’ampolla, sfavoriva l’ingresso dell’aria, mentre la bollitura, permetteva la completa fuoriuscita dell’aria all’interno. Pasteur dedusse che l’aria presentava elevata carica microbica.
La scoperta del primo vaccino , però, si deve a Jenner , e ai suoi studi sul vaiolo. Mentre, l’ungherese Semmelweis , fu il primo a sottolineare l’importanza del lavaggio delle mani , specialmente a causa dell’elevata mortalità in ambito ostetrico. Così come Joseph Lister , il quale fu promotore della sterilizzazione degli strumenti chirurgici. Anche in questo caso, l’innalzamento della temperatura era il concetto basilare nella riduzione della carica microbica. Ma qual è la differenza tra DISINFEZIONE e STERILIZZAZIONE.
DISINFEZIONE
Riduzione della carica microbica.
STERILIZZAZIONE
Eliminazione di ogni forma di vita.
POSTULATI DI KOCH
I postulati di Koch sono dei criteri destinati a stabilire la relazione di causa-effetto che lega un microrganismo ad una particolare patologia. Robert Koch fu il primo ad adottare sperimentalmente alcuni criteri, già in precedenza formulati da Henle, che altro non sono se non quattro regole generali pe r stabilire se un certo microrganismo sia o meno la causa di una certa malattia.
In particolare, Koch isolò dai tessuti di animali malati i bacilli del carbonchio , un particolare microorganismo, li coltivò in laboratorio e ne identificò il ciclo vitale. A questo punto, inoculò il microorganismo in tessuti animali sani, adesso resi infetti. In questo modo, Koch dimostrò l’esistenza di un qualche agente infettante, trasmissibile da un animale all’altro. L’insorgenza della patologia nell’animale precedentemente sano, permetteva l’isolamento dello stesso microorganismo patogeno che, se reinoculato in un nuovo animale sano, causava la patologia.
In poche parole, Koch riuscì a dimostrare le caratteristiche fondamentali di una patologia trasmissibile da un soggetto malato ad uno sano, il quale a sua volta si ammalerà. I postulati sono i seguenti:
- Il presunto agente responsabile della malattia in esame deve essere presente in tutti i casi riscontrati di quella malattia.
- Deve essere possibile isolare il microrganismo dall'ospite malato e farlo crescere in coltura pura.
- Ogni volta che una coltura pura del microrganismo viene inoculata in un ospite sano (ma suscettibile alla malattia), si riproduce la malattia.
- il microrganismo deve poter essere isolato nuovamente dall'ospite infettato sperimentalmente
Se positivi , abbiamo la prova della patogenicità del microrganismo e della sua influenza in un determinato quadro patologico.
Sebbene già nell’800 fosse già evidente l’implicazione di microorganismi quali i batteri, nell’avvento di determinate patologie, solo nel 1900 fu possibile iniziare lo studio sui virus , ed in particolare grazie agli studi condotti da Ivanovskij , il quale isolò, dalle foglie di tabacco, un virus definito: microorganismo filtrabile , dato che era tanto piccolo, da riuscire ad attraversare le maglie di un sistema di filtrazione. In particolare, Ivanovskij identificò delle foglie di tabacco fenotipicamente ‘’malate’’, che sottoposte a filtrazione, davano un estratto in grado di contagiare delle foglie sane.
Sebbene Ivanovskij propose un primo approccio al mondo della virologia, solo grazie all’avvento del microscopio elettronico , che sfrutta un fascio di elettroni, che fu possibile osservare veramente i virus.
Nel 1929, Alexander Fleming , scoprì casualmente la penicillina. In particolare, durante studi sullo stafilococco, Fleming lasciò la piastra a temperatura ambiente, la quale venne colonizzata da un micete. La replicazione del micete, impediva la crescita dello stafilococco grazie al rilascio, sulla piastra (quindi in ambiente di coltura) di un metabolita secondario: l’ antibiotico. Altro studioso degno di nota è Albert Bruce Sabin , medico e virologo polacco, famoso per aver sviluppato il più diffuso vaccino contro la poliomielite. Il merito di Sabin fu quello di riuscire ad isolare un ceppo di poliovirus attenuato in vitro, dunque un virus immunogeno, ma non patogeno.
SCOPERTA VIRUS
Il primo fu il virus influenzale. 1970 - Ebola 1980 – virus dell’HIV 2000 – Sars, virus a trasmissione 2016 – Zica
Esistono patologie a trasmissione inter-umana, e patologie a trasmissione mediante veicolo o vettore. Il veicolo può essere un alimento, mentre il vettore un animale o un insetto in particolare. La Zica è una patologia a trasmissione tramite vettore, in questo caso tramite artropode: in particolare la zanzara.
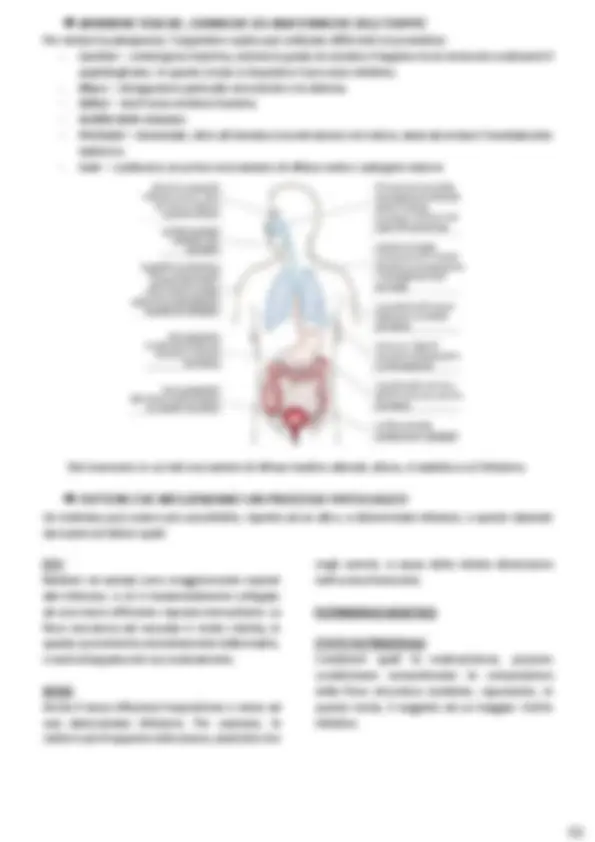
STRUTTURA
COMPOSIZIONE CHIMICA
L’ Acqua rappresenta circa l’80% del peso totale della cellula, e rappresenta il solvente in cui sono disciolti i vari costituenti organici e inorganici, che così possono muoversi ed interagire.
Tra i composti inorganici : SODIO POTASSIO MAGNESIO CALCIO FERRO ZINCO FOSFORO ZOLFO
Tra i composti organici : Sono presenti varie sostanze organiche a basso peso molecolare, per lo più macromolecole o polimeri, ognuno formato da un certo numero di sostanze organiche a basso peso molecolare (monomeri) riunite insieme secondo precise e peculiari sequenze.
COLORAZIONE DI GRAM
Al fine di poter facilitare lo studio morfologico dei batteri, si adottano diverse metodiche di colorazione: in particolare, le colorazioni adottate in batteriologia sono:
COLORAZIONI SEMPLICI Si eseguono mettendo a contatto un unico colorante con il preparato contenente il batterio da colorare. I coloranti più utilizzati a questo scopo sono il cristalvioletto, la fucsina basica ed il blu di metilene.
COLORAZIONI DIFFERENZIALI
Utilizzano, invece, più coloranti, usati in tempi successivi, e consentono di evidenziare differenze di colorazione tra specie batteriche diverse, e talora, individuare particolari strutture intracellulari. Il più importante metodo di colorazione differenziale è rappresentato dal metodo di colorazione di Gram.
STEP :
il preparato viene trattato con una soluzione di Cristal violetto per 2-3 minuti; si allontana quindi il colorante e si mordenza la colorazione mediate un trattamento di 1 minuto con una soluzione di iodio e ioduro di potassio in acqua ( liquido di Lugol ); a questo punto il preparato è trattato per 1-2 minuti con un decolorante (alcool etilico, acetone) e infine, ancora per 1-2 minuti con un secondo colorante (fucsina, safranina) di colore facilmente differenziabile (rosso) da quello del primo colorante usato (violetto).
A fine processo, alcuni batteri appaiono colorati in violetto , e prendono il nome di Gram positivi. Probabilmente ciò accade perché il decolorante non riesce ad asportare il complesso cristal violetto-iodio dalle cellule. Altri, invece, sono decolorati dall’alcool o dall’acetone ed assumono quindi il colore rosso del secondo colorante, e sono detti Gram negativi.
Dunque, il diverso comportamento dei batteri alla colorazione di GRAM, è riconducibile ad una diversa permeabilità degli involucri cellulari. In questo senso, è la permeabilità è maggiore nei Gram negativi , e minore nei Gram positivi.
Gram positività e Gram negatività, comunque, sono strettamente correlate a strutture e funzioni diverse, dunque la colorazione rappresenta un ottimo metodo per poter distinguere tra batteri significativamente differenti.
ARCHITETTURA
Essenzialmente, la cellula batterica non è altro che la cellula procariotica per eccellenza; caratterizzata da piccole dimensioni e dall’assenza di compartimenti intracellulari distinti e separati da membrana. L’assenza della membrana nucleare , implica l’immersione diretta della struttura cromosomica (estremamente semplice) nel citoplasma, il quale è delimitato verso l’esterno da una membrana citoplasmatica****.
Dalla membrana citoplasmatica , si dipartono delle invaginazioni, che costituiscono il sistema dei mesosomi. Mentre, lo strato più esterno è costituito da un sacculo o parete cellulare , alla cui superfice si trova spesso uno strato di materiale quasi sempre di natura polisaccaridica, denominato capsula.
Talvolta, in alcune specie batteriche, sono presenti delle appendici libere, i flagelli , che sono lo strumento della locomozione batterica. Oppure ancora i pili o peli , utili ai processi di coniugazione.
NUCLEOIDE BATTERICO
Il nucleo è la zona cellulare in cui è localizzato il DNA, depositario dell’informazione genica cellulare. A causa della basofilia della cellula batterica, dovuta principalmente alla ricchezza di RNA, per molto tempo si è discusso circa la presenza di un nucleo, o comunque di una struttura nucleare equivalente o simile a quella delle cellule eucariotiche. Tuttavia, attraverso l’utilizzo di micrografie elettroniche, fu possibile osservare, all’interno del citoplasma, una o più aree formate da materiale filamentoso, privo di qualunque membrana.
Tale materiale filamentoso, è costituito da DNA, e prende il nome di nucleoide.
Il nucleoide appare costituito da un unico e lunghissimo filamento con una struttura circolare. Allora, la struttura nucleare del batterio, altro non è che una singola molecola di DNA, strettamente raggomitolata e direttamente immersa nel citoplasma. Tale struttura può essere considerata l’equivalente di un cromosoma. Tuttavia, a differenza di quanto avviene in un cromosoma eucariotico, il DNA batterico non risulta essere strettamente impacchettato con gli istoni, ma lassamente complessato ad alcune proteine acidiche , da cui è, appunto, facilmente dissociabile.
Molto spesso, una cellula batterica può contenere più di un cromosoma; ed una particolare caratteristica è che il cromosoma batterico è collegato alla membrana batterica, in corrispondenza di zone caratteristiche. Oltre al cromosoma, i batteri possono anche contenere un certo numero di DNA, sempre a struttura circolare, che prendono il nome di plasmidi , dotati di una relativa autonomia replicativa, e in grado di conferire al batterio che li contiene, numerosi vantaggi funzionali.
CITOPLASMA
Essenzialmente privo dei tipici organelli eucariotici, il citoplasma batterico presenta granulazioni citoplasmatiche, che di norma rappresentano accumuli di materiale nutritivo di riserva. Solitamente, sono accumuli di glicogeno, polisaccaridi o polifosfati. Queste strutture hanno il ruolo principale, dunque, di accumulare, all’interno del citoplasma, sostanze nutritive.
Nel citoplasma batterico sono presenti i ribosomi , strumento universale della sintesi proteica. Come quelli eucariotici, i ribosomi batterici sono formati da RNA (60%) e proteine (40%) e sono c ostituiti da 2 sub unità asimmetriche : una più grande ed una più piccola. Le 2 sub unità, in particolare, hanno differenti coefficienti di sedimentazione:
MESOSOMI
La membrana citoplasmatica di alcuni batteri presenta, in corrispondenza di alcune zone, invaginazioni che si approfondano nel citoplasma e possono assumere una struttura complessa multi stratificata contenenti una serie di vescicole, tubuli e lamelle. Queste strutture sono più frequenti e complesse nei batteri Gram positivi, e prendono il nome di mesosomi. Rappresentano delle specializzazioni strutturali e funzionali, impegnate in diverse funzioni tra cui: divisione cellulare, secrezione di esoenzimi, e fosforilazione ossidativa.
PARETE CELLULARE (o SACCULO)
Una caratteristica fondamentale delle cellule batteriche, è quella di essere racchiuse da una parete rigida, molto simile alla parete delle cellule vegetali, ma strutturalmente molto più complessa. La parete presenta profonde differenze tra batteri Gram positivi e batteri Gram negativi:
GRAM POSITIVO
Presenta la membrana citoplasmatica con la tipica sezione trilaminare delle membrane biologiche.
GRAM NEGATIVO
Presenta una struttura differente, con gli involucri esterni formati da 3 diversi strati, che dall’interno verso l’esterno comprendono: a. Membrana citoplasmatica come nei Gram positivi; b. Uno strato di materiale apparentemente omogeneo e denso agli elettroni; c. Un’ulteriore struttura trilaminare che avvolge all’esterno la cellula batterica.
In entrambi i casi, comunque, la superficie esterna della cellula è circondata da uno strato di materiale mucoso (di spessore variabile), che prende il nome di strato mucoso o capsula. Il componente fondamentale della parete cellulare è rappresentato dal peptidoglicano , un polimero costituito dalla ripetizione di un’unità strutturale formata da 2 carboidrati azotati :
- N-acetilglucosamina ;
- Acido muramico – un derivato di un acido a 3C (acido D-lattico).
N-acetilglucosamina e acido muramico sono legati tra loro da un legame β-1,4.
PEPTIDOGLICANO
Al gruppo carbossilico dell’acido muramico, è legato un tetra-peptide , costituito solitamente da aa come: L-alanina, acido D-glutamico, L-lisina e D-alanina. il peptidoglicano, dunque, presenta una propria fisionomia, sia per il fatto che siano presenti amminoacidi con configurazione D, sia per il fatto che siano presenti aa esclusivi dei batteri.
Il polimero peptidoglicano è costituito da acido N-acetilmuramico ( NAM ) e N-acetilglucosamina ( NAG ) legati con legame β-1,4, a formare lunghe macromolecole lineari. Diversi polimeri lineari, sono poi collegati tra loro da legami peptidici che si stabiliscono tra D-alanina terminale di un tetra-peptide e l'acido meso-diaminopimelico in posizione 3 nel tetra-peptide adiacente. L'insieme dei polimeri lineari, collegati trasversalmente tra di loro, a livello dei tetra-peptidi , direttamente o attraverso ulteriori ponti peptidici, forma così una rigida struttura che avvolge interamente la cellula batterica.
Dunque, per costituire una struttura veramente rigida, NAM e NAG non possono soltanto essere legati tra loro, ma esistono anche dei legami trasversi, che permettono in questo modo di costituire un reticolato.
La composizione del peptidoglicano può presentare una certa variabilità tra specie batteriche diverse, soprattutto a livello della componente amminoacidica (in particolare nei batteri Gram positivi).
Per esempio, Escherichia Coli (Gram negativo) presenta ponti peptidici che legano la D-Alanina all’acido diaminopimelico. Oppure, lo Stafilococco, presenta ponti glucidici che legano D-Alanina e Lisina all’acido diaminopimelico.
Altri batteri, come la Chlamydie , hanno una composizione chimica della parete completamente differente dagli altri batteri; per esempio, caratteristica è la mancanza di acido muramico ; oppure, come nel caso dei micoplasmi , che sono caratterizzati dalla totale assenza della parete cellulare.
Il legame trans-peptidico che permette la formazione del ‘’ reticolato ’’ della parete, avviene ad opera di particolari enzimi. Farmacologicamente, è possibile bloccare l’azione di tali enzimi, impedire che si formino i legami trasversali, e impedire che si formi la parete. Un esempio di farmaci che agiscono a questo livello, è rappresentato dalla Penicillina. La Penicillina agisce a livello delle Penicillin Binding Protein , interferendo con la loro funzionalità, impedisce al batterio di sopravvivere.
GRAM POSITIVI – Struttura esterna
I Gram positivi presentano una parete cellulare molto spessa (200-800A), che circonda e racchiude completamente il batterio. Questa è formata da numerosi strati di peptidoglicano, e minori quantità di altri polimeri, come gli acidi teicoici. Il peptidoglicano rappresenta il 90% circa della parete. Acidi teicoici e loro derivati, sono polimeri di alcoli polivalenti (come glicerolo o ribitolo), esterificati con acido fosforico, cui possono legarsi a loro volta numerosi monosaccaridi e/o amminoacidi diversi (in configurazione D, come la D-alanina). Gli acidi teicoici sono altamente antigenici e presentano una notevole diversità di composizione nelle diverse specie di batteri Gram-positivi.
Sebbene la funzione degli acidi teicoici sia ancora sconosciuta, sembra che essi siano utili all’ancoraggio della parete cellulare alla membrana citoplasmatica sottostante. Ad esempio, nello Streptococco Pyogenes, gli acidi teicoici sono associati alla proteina M, che sporge dalla membrana citoplasmatica, formando una serie di micro fibrille che contribuiscono all’adesione del batterio alle superfici mucose.
Aminozuccheri carichi, aa del peptidoglicano e radicali fosforici degli acidi teicoici, rendono la parete cellulare dei Gram-positivi una struttura altamente polare. La parete impedisce il passaggio alle molecole idrofobiche, in grado di danneggiare la struttura della membrana citoplasmatica, mentre risulta permeabile alle molecole idrofile come zuccheri e aa. La polarità della parete dei Gram positivi, permette il legame con cationi utili, probabilmente, a garantire un’ambiente ionico adeguato al funzionamento degli enzimi presenti nella membrana citoplasmatica e, in particolare, a quelli preposti alla sintesi del peptidoglicano ed al suo corretto inserimento nella struttura della parete che richiedono, ad esempio, adeguate concentrazioni di Mg. Inoltre, la capacità di legare cationi, consente ai Gram positivi, di tollerare concentrazioni saline molto più elevate rispetto ai Gram negativi.
Tale caratteristica viene sfruttata in laboratorio, per ottenere terreni di coltura selettivi, in grado di favorire la crescita dei Gram positivi, piuttosto che dei Gram negativi.
Dunque, complessivamente, i Gram negativi hanno una struttura trilaminare , costituita da: membrana – parete – membrana esterna.
SECONDA MEMBRANA Mentre il foglietto interno è costituito (come di norma) da fosfolipidi, il foglietto esterno è formato da uno strato di complesse molecole che costituiscono quello che prende il nome di lipopolisaccaride batterico ( LPS ). Il lipopolisaccaride consta di 3 porzioni:
- PORZIONE LIPIDICA – Denominata lipide A , rappresenta _l’endotossina_** , ed è un glicolipide costituito da un disaccaride fosforilato ( glucosamina ), esterificato con una serie di acidi grassi saturi da 12 a 16 atomi di carbonio. Rappresenta la componente più interna.
__** Il Lipide A rappresenta la componente tossica dei batteri Gram negativi, che agisce nel momento in cui il batterio viene lisato e rilascia il lipide. Dunque, finché il batterio è integro, il Lipide A rappresenta solamente una componente strutturale, priva di azione tossica. Nel momento in cui, però, il batterio viene lisato, il lipide viene rilasciato ed esercita la sua tossicità. Il rilascio in alte dosi del Lipide A, stimola particolarmente la risposta immune, e stimola da una parte l’attivazione del complemento, dall’altra, la produzione di interleuchine e TNF, e di citochine. Dunque, il rilascio di Lipide A ha azione tossica perché attiva, improvvisamente, il rilascio di interleuchine e citochine, che richiamano i macrofagi e causano l’innalzamento della temperatura corporea. Al lipide A è ancorata , e proiettata all’esterno della membrana, la porzione polisaccaridica (catena laterale), composta a sua volta da 2 parti:
- CATENA ZUCCHERINA – corta, che forma la parte centrale o ‘’ core ’’ della molecola. Ha una struttura costante in tutti i batteri gram-negativi e presenta 2 zuccheri particolari: l’acido chetodeossiottanoico( KDO ) ed un eptoso.
- CATENA POLISACCARIDICA – rappresenta la componente più esterna , lunga, fino a 40 zuccheri, con proprietà antigeniche (che rappresentano il cosiddetto antigene 0 dei batteri gram-negativi). Questa catena è formata da una serie di sub unità tri-, tetra-, o penta-saccaridiche ripetute, comprendenti zuccheri diversi. Le varie catene polisaccaridiche sono strutture polari, in grado di legare cationi bivalenti (Mg2+) che costituiscono dei ponti tra le diverse catene, conferendo in questo modo compattezza al rivestimento.
L’insieme delle catene polisaccaridiche presenti sulla superfice dei batteri gram-negativi, sebbene formi una struttura relativamente lassa, è tuttavia efficace per la polarità delle catene polisaccaridiche e dei cationi ad esse legati, nell’escludere i composti idrofobici. Dunque, sia Gram-positivi, che negativi, proteggono la membrana citoplasmatica dalla possibile interazione con composti idrofobici dannosi; ed in particolare:
GRAM POSITIVI
Attraverso una compatta e spessa parete di polimeri peptidoglicanici e acidi tecoici.
GRAM NEGATIVI
Attraverso una parete più lassa, ma altrettanto efficace, costituita da polisaccaridi.
RIASSUMENDO La parete cellulare è utile ai batteri in quanto:
- conferisce rigidità e forma;
- protegge la cellula da shock osmotici;
- la protegge dagli scambi con l’esterno in maniera incontrollata;
- determina le proprie capacità tintoriali;
- condiziona gli scambi con l’esterno, perché sia nei Gram+, che nei Gram- sono presenti le porine, proteine che hanno un ruolo attivo nel trasporto dei nutrienti;
- contiene importanti fattori di patogenicità: l’endotossina nel caso dei batteri Gram- e il fattore cordale, costituenti caratteristici della parete dei micobatteri che rappresentano anche in questo caso una struttura antigenica.
La parete può essere lisata attraverso l’utilizzo di una soluzione contenente Lisozima , un enzima in grado di rompere il legame β-1,4-glicosidico tra NAM e NAG. Dunque in ambiente ipotonico , il batterio privo di parete, non ha più la protezione osmotica e va incontro a lisi. In ambiente isotonico , invece, il batterio privo di parete prende il nome di protoplasto , e può sopravvivere solo se mantenuto in condizioni di equilibrio. In condizioni adeguate, comunque, un batterio privato della propria parete, può in qualche modo risintetizzarla. Proprio tale caratteristica ha permesso di studiare le diverse fasi di sintesi della parete batterica.
MECCANISMI DI TRASPORTO La membrana esterna, oltre ad escludere i composti idrofobici, non consente, a causa della sua natura lipidica, il passaggio delle molecole idrofiliche. Per questo motivo, la cellula ha evoluto una serie di canali specifici, che permettano la diffusione passiva di numerose molecole come zuccheri, aa e ioni.
I canali sono costituiti da particolari molecole proteiche, dette porine , che in coppia o sotto forma di trimeri, attraversano la membrana esterna. Le porine costituiscono un canale abbastanza ampio da permettere la diffusione passiva di tutti i composti idrofili di dimensioni inferiori a 600-700 dalton. L’ingresso di molecole di maggiori dimensioni richiede, invece, la presenza di sistemi di trasporto specializzati (carrier).
La membrana esterna è collegata alla sottile parete cellulare attraverso una serie di lipoproteine presenti in circa 700.000 copie. Anche le porine, che attraversano la membrana esterna, contribuiscono all’ancoraggio della membrana esterna al peptidoglicano, attraverso legami non covalenti.
PERIPLASMA o SPAZIO PERIPLASMICO – dei Gram negativi
Il doppio sistema di membrane che avvolge i batteri Gram negativi, delimita un compartimento, costituente il 20-40% del volume cellulare, chiamato spazio periplasmico o periplasma. Il periplasma, oltre al peptidoglicano, contiene una sorta di gel di proteine, con diverse funzionalità:
- Proteine – in grado di legare molecole idrofile di basso peso molecolare come zuccheri, amminoacidi e ioni. In questo modo, impediscono che tali sostanze possano modificare il gradiente di concentrazione e me veicolano l’introduzione in ambiente cellulare attraverso diffusione passiva.
- Enzimi – come fosfatasi, proteasi o nucleasi, in grado di degradare le grosse molecole introdotte attraverso i sistemi di trasporto, in modo da ridurle a dimensioni compatibili al passaggio attraverso i sistemi di trasporto della membrana citoplasmatica (digestione);
- Enzimi detossificanti – in grado di attivare alcuni farmaci antibatterici.
FLAGELLI
I flagelli batterici sono formati da un unico filamento sprovvisto di membrana, e sono tipici di batteri di forma cilindrica come bacilli, vibrioni e spirilli. A seconda della zona di inserzione dei flagelli, si distinguono:
BATTERI CON FLAGELLI POLARI Quando i flagelli sono localizzati su uno o su entrambi i poli cellulari.
BATTERI PERITRICHI
Batteri con più flagelli sparsi su tutta la superficie cellulare.
In base al numero di flagelli, i batteri possono essere distinti in:
- MONOCRITI – 1 solo flagello;
- ANFITRICHI – fagelli da entrambi i lati della cellula;
- LOFOTRICHI – gruppo di flagelli da un solo lato;
- PERITRICHI – flagelli sparsi su tutto il perimetro cellulare.
Un singolo flagello è composto da 3 parti:
Filamento – formato dalla ripetizione di una serie di sub unità proteiche di flagellina. Le flagelline sono dotate di spiccate proprietà antigeniche e rappresentano l’antigene H dei batteri provvisti di motilità.
Gancio – struttura proteica, sempre formata da diverse sub unità, con diametro leggermente inferiore al filamento.
Corpo basale – costituito da diverse sub unità di 15 diverse proteine, che si aggregano a formare la parte più prossimale del flagello, chiamata bastoncello , caratterizzato, nei Gram negativi, da una serie di 4 anelli:
- Anello L – da Lipopolisaccaride, in corrispondenza della membrana esterna.
- Anello P – da Peptidoglicano, in corrispondenza della parete cellulare.
- Anello S – da Super membrana, superiormente alla membrana plasmatica.
- Anello M – da Membrana, in corrispondenza della membrana plasmatica.
La parte prominente è rappresentata da un lungo filamento elicoidale che protrude dalla cellula per 5- 10μm e che, attraverso un gancio tubolare, è connesso alla complessa struttura del corpo basale che ancora il flagello ali involucri cellulari e rappresenta il motore del movimento rotatorio del filamento.
N.B. – Nei Gram positivi , la struttura del corpo basale è molto più semplice , dato che sono presenti solo
l’anello P ed M , in rapporto alla diversa organizzazione degli strati superficiali della cellula.
Gli anelli ancorano il flagello alla cellula e fungono da ‘’ motore ’’ per il movimento del flagello stesso. L’energia utile al movimento, è generata dal potenziale di membrana durante il trasporto di elettroni nel corso del processo di fosforilazione ossidativa o, nei batteri anaerobi , per idrolisi dell’ATP. Il movimento rotatorio dei flagelli può avvenire sia in senso orario, che antiorario. Poiché i flagelli sono strutture elicoidali con passo sinistrorso, una rotazione antioraria provoca una propulsione monodirezionale ( swimming ) della cellula, mentre una rotazione oraria, provoca un movimento improduttivo detto tumbling (avanti e indietro).
Il movimento rotatorio dei flagelli, è condizionato da una serie di chemiocettori di superficie, che rispondono a stimoli di numerose sostanze.
Le Spirochete , sono batteri particolarmente sottili, con forma elicoidale, flagellati. I flagelli, in questo caso, non si trovano esternamente alla membrana, ma sono degli endoflagelli , di natura proteica, localizzati internamente al corpo del batterio. Tali flagelli si muovono in maniera retrattile, permettendo al flagello di invadere efficacemente una mucosa.
FIMBRIE
O pili, sono appendici proteiche che si proiettano all’infuori degli involucri cellulari. Originano dalla membrana plasmatica, e si estendo per 0.2 – 2 μm all’esterno. Le fimbrie sono formate da piline ripetute, specifiche per le diverse specie batteriche. Anche in questo caso, le sub unità si organizzano con una simmetria elicoidale intorno ad un asse immaginario, a formare una serie di rigide strutture cilindriche.
La capacità delle fimbrie di legarsi a specifici substrati, è conferito da altre proteine, che prendono il nome di adesine. Le fimbrie infatti sono specifici organi di ancoraggio in grado di interagire con vari residui di carboidrati presenti nelle glicoproteine di membrana di varie cellule animali e sono in grado, quindi, di condizionare l’iniziale colonizzazione della superficie mucosa e l’avvio del processo infettivo.
Altra varietà di fimbrie , è costituita da curli , proteine della famiglia delle proteine amiloidi , che hanno la capacità di mediare l’adesione batterica ad una grande varietà di proteine dell’organismo ospite quali il plasminogeno, la fibronectina e numerose proteine di adesione intercellulare, e in grado interagire con le proteine del complesso maggiore di istocompatibilità di classe I (MHC I); inoltre sono implicate nella formazione di biofilm.
Oppure ancora, i pili F , o pili sessuali , in genere molto più lunghe delle fimbrie adesiniche, che giocano un ruolo fondamentale nei processi di coniugazione batterica , consentendo la connessione iniziale tra i due batteri interessati al processo di scambio di materiale genetico.
GLICOCALICE
Oltre alla capsula, altri batteri presentano un glicocalice , una struttura più lassa, composta da fibre polisaccaridiche, che fornisce importanti capacità adesive.
SPORA
Altra struttura accessoria sintetizzata da batteri sporigeni, che rappresenta una forma di resistenza. Tali batteri producono spore nel momento in cui si trovano in condizioni ambientali sfavorevoli alla vita. La spora è molto resistente a:
- Calore
- Essiccamento
- Radiazioni e raggi UV
- Colorazione
- Agenti disinfettanti e chimici
- Lisozima Tale resistenza è dovuta alla presenza di numerosi rivestimenti esterni, che racchiudono l’informazione genetica (il DNA) del batterio. Il materiale genetico è racchiuso da una membrana citoplasmatica e da una parete cellulare rudimentale. Tra i rivestimenti citati in precedenza:
- Una corteccia – ricca di acido adipicolinico e calcio , che conferiscono elevata resistenza;
- Un rivestimento interno – costituito da proteine simil-cheratiniche molto solubili, ricche di cisteina.
- Un rivestimento esterno ;
- Un esosporio.
LA SPORA
I batteri si riproducono per scissione semplice. Alcuni bacilli Gram positivi, aerobi ed anaerobi (appartenenti rispettivamente ai generi Bacillus e Clostridium ) in determinate condizioni ambientali danno luogo alla formazione di particolari strutture denominate Spore.
Le Spore batteriche sono delle endospore , ovvero: originano all’interno della cellula madre (detta sporangio ), che in seguito a rottura le rilascia libere nell’ambiente circostante. Le spore rappresentano una forma di resistenza, e consentono ai batteri di sopravvivere in ambienti sfavorevoli.
Per esempio, il Clostridium Tetani , è un anaerobio obbligato, che per sopravvivere al di fuori dell'ospite, dove vi sarà ossigeno, deve esistere sotto forma di spora. Infatti, solitamente, il batterio vive nell'intestino degli equini dove non vi è ossigeno, quando grazie alle feci viene rilasciato all'esterno, si trova in condizioni sfavorevoli e si adatta, sopravvivendo sotto forma di spora.
RIPRODUZIONE BATTERICA
I meccanismi di riproduzione batterica sono sicuramente più semplici rispetto a quelli delle cellule eucariotiche, ma comunque sono volti alla ripartizione del materiale genetico dalla cellula madre, alle cellule figlie. La replicazione del DNA batterico è di tipo semiconservativo. Il DNA batterico è a doppio filamento, senza introni ed istoni, codificante in entrambi i sensi. Il primo enzima realizza la forca replicativa , in cui ognuno degli stampi funge da stampo per l’acido nucleico nascente.
Il DNA batterico è organizzato in un’unica struttura circolare , che prende il nome di cromonema (rappresenta il cromosoma batterico). Questa struttura è ancorata alla membrana citoplasmatica , dunque, durante il processo di duplicazione, si formeranno 2 nuovi cromosomi , ognuno con un proprio punto di ancoraggio alla membrana.
Partendo dal punto di separazione tra le due zone della membrana in cui sono ancorate le strutture cromosomiche, si ha l’accrescimento della membrana e il conseguente allungamento cellulare.
Le 2 strutture cromosomiche vengono allontanate sempre più dal continuo accrescimento nella zona centrale. In questo modo, le due strutture cromosomiche, seguendo passivamente lo spostamento del punto di attacco alla membrana durante l’accrescimento cellulare, vengono distanziate in modo sufficiente perché la separazione delle due cellule figlie, le trovi dislocate nelle due zone corrispondenti del citoplasma. La separazione avviene per scissione semplice: la formazione di un setto che si diparte dalla membrana citoplasmatica, e che si approfonda nel citoplasma, causa una separazione netta della cellula madre, nelle 2 cellule figlie. In particolare, il setto si forma lungo un piano perpendicolare all’asse maggiore della cellula nei Bacilli ; mentre equatoriale nei Cocchi.
FASE L BATTERICA
Rappresenta un’eccezione nelle modalità di riproduzione batteriche. Alcuni batteri, infatti, sono caratterizzati dall’assenza di una parete cellulare rigida , per cui risultano molto fragili, con una forma variabile e molto sensibili a varie situazioni ambientali.
Un’analisi morfologica di questi particolari batteri, evidenzia dimensioni piuttosto variabili: cellule di notevoli dimensioni, affiancate da altre cellule di dimensioni piccolissime. Queste osservazioni fanno credere che i batteri, in fase L, seguano un particolare ciclo riproduttivo caratterizzato da:
- Cellule giganti – originate per divisione multipla del cromosoma;
- Cellule piccole – derivate da serie di divisioni citoplasmatiche.
Nonostante l’assenza della parete rigida, i batteri in fase L presentano numerosi caratteri antigenici comuni alla specie originaria, e possono nuovamente dare origine a cellule batteriche normali. Molte specie batteriche, comunque, sembrano in grado di produrre forme L, e sembra che in genere ciò accada in relazione a condizioni ambientali sfavorevoli alla sintesi del peptidoglicano (come per esempio, in presenza di antibiotici come la penicillina).
Sembra dimostrato che la produzione di forme L sia possibile in vivo e che essa possa rappresentare il meccanismo attraverso il quale si mantengono cronicamente alcune infezioni.
PRODUZIONE DELLE SPORE
Come già detto, la produzione di spore è tipica di alcuni bacilli.
BACILLUS
Sono caratterizzati da spore con diametro non superiore a quello della cellula batterica (sporangio).
CLOSTRIDIUM
Sono caratterizzati da spore con diametro maggiore rispetto allo sporangio. La cellula appare ingrossata, quindi, in corrispondenza della spora.
STRUTTURA
Il citoplasma è circondato da membrana plasmatica, nel cui versante interno è addossato il materiale nucleare. La membrana appare rivestita da una rudimentale parete cellulare, costituita da peptidoglicano. La super struttura, appare costituita da diversi strati di membrane molto voluminose, che dall’interno verso l’esterno sono:
CORTECCIA (o Cortex)
Formata da peptidoglicani con inglobati residui citoplasmatici dello sporangio.
2 RIVESTIMENTI (o Coats)
Esterno ed interno , composti da proteine ricche di legami disulfidrilici, contenenti una piccola quantità di lipidi (1-2%) e una certa quantità di peptidoglicano.
ESOSPORIO
Una sottile membrana strutturalmente complessa, di composizione fosfolipo-proteica simile a quella della membrana plasmatica, contenete varie quantità di acidi teicoici , tracce di acido diaminopilemico e glucosamina.
Un componente caratteristico della spora, che non ha riscontro nella cellula vegetativa, è l’ acido dipicolinico , un acido dicarbossilico che, insieme a grandi quantità di calcio, si trova localizzato nella cortex di cui sembra contribuisca a stabilizzare la struttura.