Scarica Sbobine Microbiologia - cdl in biotecnologie e più Sbobinature in PDF di Microbiologia solo su Docsity!
MICROBIOLOGIA GENERALE
N.B. diapositive: indice preciso delle cose trattate. (quello che non c’è nelle slide non va studiato) (in tal modo stiamo circoscrivendo ciò che ci interessa: per rendere chiaro ciò che c’è dentro ciò che c’è fuori: sappiamo in che ambito muoverci) LEZIONE 1 - alla scoperta del mondo microbico I microrganismi sono tutto ciò che sostanzialmente non vediamo: batteri, muffe, alghe, protozoi ecc… Definizione: I microrganismi sono:
- organismi (organismo: struttura organizzata)
- viventi (termodinamicamente instabile, complessità: la vita finisce)
- totipotenti permanenti (la cellula può fare tutto ad adempiere a tutte le funzioni che la vita richiede e resta tale in ogni momento, non come il nostro organismo che perde questa capacità ai primi stadi dello sviluppo). Il metabolismo sta in un triangolo i cui vertici sono:
- Identità: chi è l’organismo, con quel patrimonio genetico. (soggetta ad evoluzione per mutazione e selezione)
- Riproduzione: identità resta tendenzialmente uguale tranne qualche lieve modificazione. (mutazione / segregazione / combinazione)
- Evoluzione: nasce per lungo periodo dalla sequenza di riproduzione. Viene trascinata dalla selezione e altri fattori
Un organismo dotato di metabolismo sta fra la sua identità (cellula simile a chi l’ha generata e simile a quellr che genererà), una riproduzione ed una evoluzione.
- Bilancio tra riproduzione ed evoluzione → selezione
- Bilancio tra identità e riproduzione → fedeltà
- Bilancio tra identità ed evoluzione → mutazione TASSONOMIA: legge del taxon, disposizione dei soldati in battaglia → modo in cui ordinare le nostre informazioni sul mondo dei viventi. Ogni classificazione riposa su una teoria e da essa (questa classificazione) partirà una nuova teoria. Con la teoria cellulare nascono diverse letture del mondo, ognuna delle quali sviluppa qualcosa di interessante. Con la nascita del microscopio elettronico ci fu un cambiamento nella visione del mondo: si possono leggere le infrastrutture delle cellule → prima divisione tra eucarioti e procarioti, seguirono poi ulteriori classificazioni sempre più precise e simili a quelle che abbiamo noi oggi.
- organismi superiori eucariotici: Protisti, protozoi (hanno dato vita ad animali), alghe (nella loro evoluzione: piante), funghi ( lieviti, muffe – funghi superiori)
- eukarya (piante e animali). Tutti questi organismi sono caratterizzati dall’assorbimento: i nutrienti vengono presi sotto forma di soluzione, mentre gli animali (protozoi) ingeriscono. (SCHEMINO) Assorbimento antitetico alla mobilità
- procarioti: bacteria e archaea
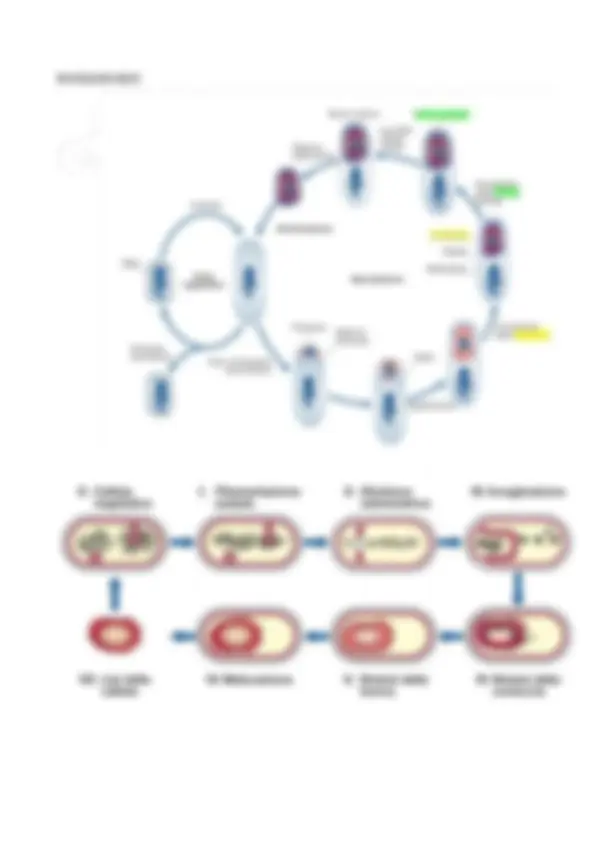
Lezione 2 - STRUTTURA E FUNZIONE DELLE CELLULE PROCARIOTICHE I batteri hanno dimensioni intorno al micron (1-2) e si dividono in due CATEGORIE MORFOLOGICHE fondamentali:
- Cocchi : possono stare da soli o in gruppo ✓ diplococchi - streptococchi → crescita su un solo piano ✓ tetradi → crescita su più piani: 4 cellule disposte ai 4 lati di un quadrato immaginario (2 direzione di crescita) ✓ sarcina → otto cellule ai vertici di un cubo ideale con 3 direzioni di crescita ✓ stafilococco (simile ad un grappolo d’uva)
- Forme allungate: ✓ bacillo: lunghezza 2/3 volte diametro ✓ coccobacillo: ibrido tra cocco e bacillo ✓ diplobacillo: due bacilli ✓ streptobacillo: con più bacilli che continuano a riprodursi sullo stesso piano (forma a collana) ✓ vibrione ✓ spirillo: forma spiralata, con una sezione circolare ✓ spirochete: forma spiralata algale con una sezione non circolare ma a rettangolo stretto Questi microrganismi hanno DIVERSI MODI DI CRESCITA , in base alla forma finale che devono assumere. In tutte le cellule è presente un elemento detto FTsZ, proteina essenziale per la divisione cellulare simile alla tubulina, che al momento della divisione si aggrega formando un anello, nel piano equatoriale ove recluta le altre proteine della divisione. Dunque forma il setto centrale e separa le due cellule.
- Cocchi: solo FTsZ
- Bacilli: hanno la MreB presente in tutte le forme allungate
- Forme vibrioniche: crescentina , uno dei filamenti intermedi, responsabile della crescita differenziale del lato concavo rispetto al convesso.
I microrganismi procarioti si dividono in due categorie in base alla PARETE :
- Didermi – gram negativi → la loro particolarità è l’avere un periplasma esterno ed una membrana esterma: due pareti
- Monodermi – gram positivi: parete come una spunga, permeabile, aperta ma spessa, unica parete Tutto deriva da una colorazione che rende possibile la distinzione in queste due grandi categorie:
- Fissaggio al calore : le cellule vengono fissate al calore su un vetrino (cellule sopra ad una fiamma – può essere anche al vapore o all’aria).
- Colorante : Viene utilizzato un colorante, il cristal violetto, molecola planare ed aromatica, fluorescente che colora indistintamente tutte le cellule di violetto
- Soluzione di Lugol : si aggiunge la soluzione di lugol o ioduro di potassio e la cellula va a formare un complesso iodio iodurato cristal violetto di dimensioni enormi che rimangono intrappolate nella parete dei gram positivi.
- Lavaggio con decolorante: lavando il tutto con un decolorante come acetone o alcol, avremo due possibilità:
- quelle strutture che non sono ingabbiate tra membrana esterna e parete, per decolorazione si decolorano, perché c’è poco da colorare → hanno due pareti ma sono sottili
- mentre quelle con la parete più spessa rimangono colorate: colorazione sull’azzurro
- Per sicurezza si una la safranina , colorante generico, che entra nella cellula colora tutte le cellule in due modi distinti:
- Dal trasparente al rosso: didermi, gram negativi
- Dall’azzurrino al violetto più acceso: monodermi, gram positivi La prima funzione della parete è quella di dare la forma alle cellule.
Nella membrana ci sono alcune sostanze, disciolte tra gli acidi grassi che presentano degli anelli aromatici apolari (solo nello strato centrale), il loro scopo è quello di irrigidire la membrana, garantendo la giusta fluidità. Ne abbiamo due tipi:
- steroli: negli eucarioti (colesterolo)
- opanoidi: nei batteri La membrana ha la funzione di:
- contenere il citoplasma
- barriera selettiva
- trasduzione del segnale
- siti di reazione ed immagazzinamento
- biosintesi componenti cellulari
- produzione di energia La membrana è anche l’obiettivo di alcuni antibiotici: ci sono alcuni agenti che bloccano la produzione di proteine e dunque la funzionalità della membrana. I peptidi:
- alterano equilibrio elettrochimico della membrana creando delle fratture che comportano alla lisi delle sostanze
- attuano l’inibizione delle funzioni vitali Quali sono le batteriocine, sostanze di origine batterica che possono inibire le funzioni cellulari?
- Colicine > 40kDa (dall’escherichia coli di solito)
- Microcine > 10kDa (gram positiva)
- Batteriocine 5-30kDa
Ogni microrganismo sviluppa queste sostanze per danneggiare altre cellule esercitando azione tossica su:
- parete
- membrana
- sintesi DNA e RNA
- sintesi proteica Antibiotici e membrane :
- polimixine: pori sulle membrane, membrana non contenitiva
- lipopeptidi, daptomicina: codina che si intrufola nella parte idrofilica e va in quella idrofobica dove crea un foro mediante il metabolismo del Ca, favorendo la fuori uscita di K → perdita potenziale osmotico
- Polieni: si teme una nuova pandemia a causa di un lievito, la candida auris. Anfotericina B → crea canali complicati e mastodontici che intearagisce con le molecole che stabilizzano la membrana come gli opanoidi e gli steroli. Ergosterolo
- Peptidi: presenti nell’industria alimentare → bloccano sintesi del peptidoglicano (parete) e provocano fori nella membrana. TUTTI PRUDOCONO PORI
PARETE : caratteristica dei procarioti, ma presente anche in alghe e funghi.
Funzioni della parete:
- Turgore cellulare : la parete grossolana è rigida e dunque evita che la cellula esploda quando la cellula è posta in ambiente isotonico In condizioni ipotoniche la parete evita l’esplosione e favorisce un equilibrio fra pressione osmotica (da fuori a dentro) e pressione idrostatica. Acidi lipoteicoici: servono ad ancorare la membrana alla parete e in condizioni ipertoniche (fuoriuscita acqua), la parete trattiene la membrana vicino a sé e ne evita il collasso – plasmolisi (la cellula è schiacciata, non turgida e il citoplasma non ha spazio per esistere)
- Protezione meccanica : la membrana è delicata, mentre la parete è un carro-armato robusto
- Conferisce e mantiene la forma
- Contribuisce all’isolamento dall’esterno con la membrana
- Favorisce la difesa da altri microrganismi
- Fa parte del sistema di “attacco ” ad altre cellule La parete mureica può essere indebolita o rimossa in due modalità:
- Antibiotici che inibiscono la formazione del peptidoglicano
- Enzimi specifici che lo degradano Questi trattamenti portano alla lisi osmotica, ma tuttavia in ambiente isotonico si possono ottenere cellule sferoidali prive di parete: i protoplasti → se trasferiti in ambiente ipotonico: lisi. Il cuore della parete è il peptidoglicano, ha un complessa struttura reticolare formata da catene glicaniche, costituita da da due amnozuccheri alternati, legati da un legame β (1→4), di diversa lunghezza:
- NAG
- NAM
Pareti a confronto: Il peptidoglicano funziona come un cavo che forma una specie di spirale come nel bacillus subtilis, si avvolge intorno alla cellula. (la struttura è stata studiata solo su pochi batteri)
La sintesi del peptidoglicano
Ci sono diversi passaggi:
- FASE 1 – citoplasma: sintesi dei precursori
I precursori sono NAM e NAG legati all’UDP → UDP-NAM + UDP-NAG
- UDP-NAG: a partire dal fruttosio- 6 - P + UTP
- UDP-NAM: a partire da UDP-NAG + PEP grazie MurA e ligasi mur La struttura non è quella finale perché necessita di maturazione, infatti ne troviamo 5 al posto di 3.
- FASE 2 – transmembrana: sintesi dell’unità monomerica. I precursori devono essere trasportati nello spazio periplasmatico per essere poi inseriti nella parete. Il trasporto è garantito dal bactoprenolo (con due P), un alcol fortemente idrofobo che consente di attraversare la membrana. E avvengono i seguenti processi:
- NAM + bactoprenolo → LIPIDE I
- Lipide I + NAG → LIPIDE II N.B. Il bactoprenolo, terminato il trasporto, viene defosforilato e ribaltato pronto per un nuovo ciclo.
- FASE 3 – extramembrana: reazione di polimetizzazione e transpeptidazione→ formazione dei legami crociati. (si differenzia a seconda del gram)
- Polimerizzazione: transglicosilasi catalizzano la formazione tra legami glicosidici tra in nuovo monomero e il glicano nascente → legame peptidico
- Transpeptidazione: transpeptidasi catalizzano la formazione del legame crociato → legame peptidico tra le due D-alanine viene traferito al gruppo aminico libero della lisina e dunque viene liberata una delle due D-alanine. N.B. (geni sono scritti in corsivo, enzimi in carattere normale) Le transpeptidasi hanno una forte affinità per gli antibiotici B-lattamici infatti vengono definite come “proteine che legano la penicillina” → la penicillina
Lezione 4 PARETE GRAM POSITIVI La parete dei batteri gram positivi è una complessa struttura macromolecolare essenziale per:
- forma
- funzionamento
- integrità ed è costituita da uno spesso strato di mureina in cui sono inserite altre molecole:
- Acidi teicoici → dentro le maglie grosse della parete e tengono stretti i vari strati di peptidoglicani
- Acidi lipoteicoici → hanno una codina lipidica e una testa idrofila che sta fuori. Non sono molecole moto grandi, non creano una porosità, ma si ancorano alla membrana e ancorano la parete alla membrana
- Proteine di superficie Dunque questa complessa matrice polianionica:
- Definisce le caratteristiche chimico-fisiche: porosità, elasticità, resistenza alla tensione
- Controlla il movimento degli ioni e di altre molecole
- Regola il movimento e l’attività di proteine periplasmatiche
- Media: ❖ Adesione delle proteine alla parete ❖ Riconoscimento da parte di batteriofagi ❖ Interazioni con il mondo esterno ALTRI TIPI DI PARETE NEI BACTERIA Gram variabili : batteri filogeneticamente appartenenti e gram positivi che si colorano diversamente a seconda della fase di crescita. molte cellule di questi batteri sono più fragili a livello del setto di divisione rilasciano il colorante di gram, risultando fenotipicamente gram-negativi. Microbatteri : presentano una parete cellulare spessa ricca di componenti cerose che formano uno strato impermeabile → cioè rende inutilizzabili le solite colorazioni, chiusa quella di ziehl-neelsen. La parete è formata da due strati:
- Basale, o core, formato da peptidoglicano + arabinogalattano - polisaccaride ramificato Complesso dove residui di zuccheri che lo formano sono in forma furanosica (struttura ad anello a 5C)
- strato di:
❖ Acidi micolici: acidi grassi complessi a lunga catena alchilati ed
idrossilati → contribuiscono alla scarsa fluidità della membrana
❖ Cere: glicolipidi complessi con catene carboniose lunghe e
ramificate che interagiscono e si legano agli acidi micolici in modo non covalente. Archea : architetture e composizione della parete degli Archea sono molto diversificate: nella parete cellulare non sono presenti nè acido muramico ne D-
amminoacidi che caratterizzano il peptidoglicano. Gli archea metanogeni contengono lo pseudopeptidoglicano, un polisaccaride composto dalla ripetizione di subunità costituite da 2 aminozuccheri, legate da legame B (1 → 3):
- NAG
- NAT Queste differenze chimiche tra lo pseudopeptidoglicano il peptidoglicano rendono e metanogeni resistente al lisozima e agli antibiotici beta lattamici. PARETE DEI GRAM NEGATIVI I gram negativi contengono all’esterno della membrana particolari strutture che rendono più complessa la loro parete. Infatti all’esterno sono circondati da una particolare membrana esterna, differente da quelle biologiche → lo spazio compreso tra la membrana esterna ed interna prende il nome di periplasma , contenente una sottile parete mureinica con un sottile strato di peptidoglicano.
Il periplasma: compartimento acquoso che occupa dal 20 al 40% del volume
cellulare totale. N.B. Nel caso in cui in condizioni ipotoniche sia danneggiata la membrana esterna, ma non l’interno, le proteine periplasmatiche possono emergere ed essere studiate. Le proteine periplasmatiche hanno diverse funzioni:
- Acquisizione di nutrienti
- Generazione di energia
- Sintesi del peptifoglicano
- Biogenesi della membrana esterna e di altri rivestimenti
- Catabolismo iniziale di nutrienti complessi
- Protezione da antibiotici o altre sostanze tossiche Il corretto ripiegamento delle proteine nel periplasma e di altre che devono essere trasportate alla membrana esterna sono controllate dalle proteine chaperon periplasmatiche , il cui funzionamento non richiede ATP.
La membrana esterna , formata da:
- Fosfolipidi
- LPS – lipopolisaccaride con: ❖ Lipide A: formato da n-acetilglucosamina ed è ciò che ancora LPS alla membrana esterna ❖ Core che si divide in : interno - formato da 1 a 3 molecole di kg ed ha esploso ed esterno - formato da antigene 0. Il LPS rappresenta anche, nell’organismo dell’ospite, un importante segnale alla presenza batterica:
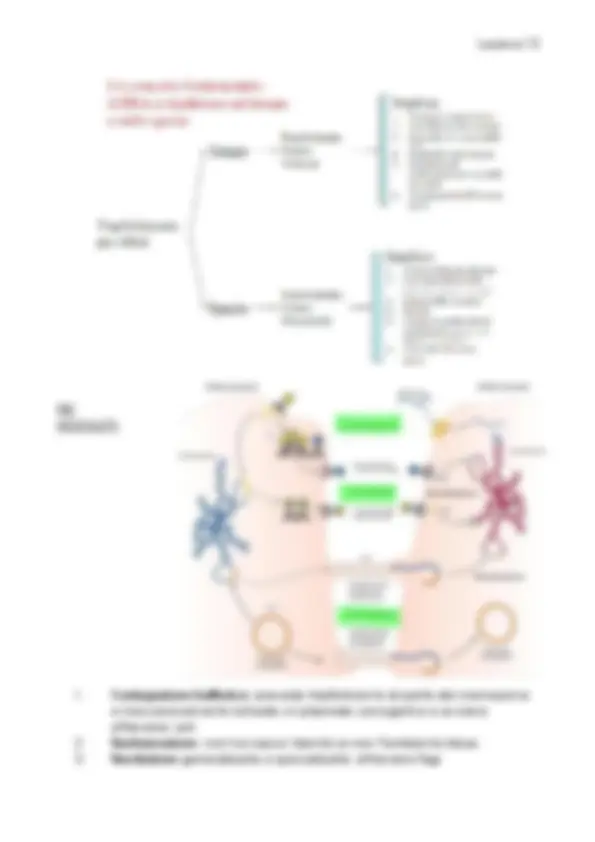
- monotrichi: singolo flagello
- anfitrichi → un flagello ai due pori
- lofotrichi → ciuffo di flagelli ad un polo
- peritrichi → numerosi flagelli che protrudono da tutta superficie Struttura:
- corpo basale → ancora il flagello all’involucro cellulare e costituisce il motore flagellare, ed è formato da anelli cilindrici, i bastoncelli ❖ anello L: al livello della membrana esterna (lipide A) ❖ anello P: peptoglicano ❖ anello MS: va dalla membrana plasmatica fino al periplasma ❖ anello C: citoplasma, è il rotore del motore flagellare, mette in movimento ed è formato da 3 proteine – flig G/M/N La componente non rotante: statore → formata da proteine integrali: ❖ MotA: interagisce in modo diretto con le subunità flig ❖ MotB: ancorata al peptidoglicano.
- Uncino: struttura flessibili e corta che connette corpo basale al filamento
- Filamento: struttura formata da 20000 subunità di una proteina detta flagellina
Movimento: rotazione oraria/antioraria del filamento flagellare grazie all’energia proveniente dal gradiente elettrochimico. Il flusso di protoni attraverso proteine MotA e MotB caus un cambiamento conformazionale in MotA che muove flig , permettendo uno spostamento in senso antiorario → la forza rotatoria è trasmessa dall’anello C, mediante un anello MS e dunque il movimento si propaga da: bastoncello – uncino – filamento flagellare. La proteina del rotore fliM , interagento con CheY del sistema di chemiotassi, determina il cambiamento di rotazione in senso orario. Si differenzia a seconda del tipo di batterio:
- Batteri monotrichi → flagello fa muovere il batterio: ❖ Avanti: quando muove in senso anti-orario ❖ Indietro: quando muove in senso orario
- Batteri peri/lopotrich ❖ Senso orario: apertura dei fasci: capovolgimento casuale del batterio - capriole ❖ Senso antiorario: movimento di avanzamento – corsa, che permette l’unione di fasci compatti N.B. Negli Archaea: pochi studi sulla loro motilità, apparentemente simili a quelli dei batteri → strutture rotanti con:
- Filamento – più sottile dei batteri
- Uncino La loro direzione è controllato dai sistemi di chemiotassi, ma le modalità di biosintesi dei due organelli cambiano. Il movimento rotatorio avviene utilizzando l’energia a partire dall’idrolisi di ATP, ma ancora non si sa quale sia il motore che genera la forza. La motilità permette al batterio di:
- spostarsi verso ambienti favorevoli
- allontanarsi da ambienti ostili