Download biochem biochem EXP5.docx and more Exams Biochemistry in PDF only on Docsity!
Experiment 5: ISOLATION, PURIFICATION AND CHARACTERIZATION
OF LYSOZYMES FROM EGG WHITE
Introduction
The clear liquid of an egg is called egg white also known as albumin. It surrounds the yolk of the
egg providing protection and nutrition. It consists of 88% of water and only 11% of proteins and
remainder as carbohydrate, ash and 1% of lipids. There are a number of proteins that make up the
11% of albumin. Ovalbumin 54%, has three fractions A1, A2 and A3 that differ from each other
based on their phosphorus content. It is a storage glycoprotein and calcium binder that plays a
vital role in storage of amino acids around the body (Abeyrathne, Lee and Ahn, 2013).
Ovomucoid 11%, is an allergen, has trypsin inhibitory activity and inhibits growth of
tumour cells. Ovomucin 3.5%, has an alpha subunit and a β‐subunit that are bound by disulfide
bonds. The β‐subunit has cytotoxic effect on cultured tumour cell, suppressing the cell.
Ovotransferrin 12%, has antibacterial and antiviral activities, involved in transport of iron to
target cells and aggregates by heating resulting in milky white gel. Lysozyme 3.5%, is highly
stable in acidic solution and heats at 100°C for 1-2- minutes. Their stability is due to 4 disulfide
bonds. It catalyzes the hydrolysis of 1,4-glycosidc linkage between N-acetylmuraminic acid and
N-acetylglucosamine in peptidoglycan layer of bacterial cells walls specifically the gram-
positive bacteria. This activity can be enhanced by EDTA, tripolyphosphate and butylparaben.
These are the main proteins. Ovomacroglobulin 0.5%, glycoprotein has ability to inhibit
hemagglutination. Cystatin 0.05%, is a sulphydryl proteinase activity inhibitor used as
antimicrobial, insecticidal and antiviral agent as well as control of cancer cell metastasis and
prevention of cerebral haemorrhage. Ovoflavoprotein 0.8%, flavoprotein binds to riboflavin in
1:1 ratio in egg white and transfers it from blood serum to albumen of egg while. Avidin 0.05%,
is a strongly basic glycoprotein which combines with biotin forming a stable complex that is not
absorbed by intestinal tracts of animals. Ovoinhibitor 1.5% inhibits proteolytic enzyme and
inhibits bacterial serine proteinase, chymotrypsin and fungal serine proteinase. Ovoglycoprotein
1.0% is a foaming agent consisting of G2 and G3 globulins. (Stevens, 1996). The egg white
proteins can be extracted through various approaches. Proteins can be salted out based on their
ionic strength or the salt concentration such as ammonium sulphate (NH 4 ) 2 SO 4 salt or other salts.
The (NH 4 ) 2 SO 4 is a highly soluble salt that stabilizes protein structure by preferential solvation
and inhibits bacterial growth. Increasing the salt concentration decreases solubility of proteins
and at high ionic strength the protein will be precipitated out of solution. The dissociates of salt
NH 4
and SO 4
2- are attracted to charges on compound being purified preventing water molecules
from binding to it and allowing precipitation (Coen, Blanch, and Prausnitz, 1995). Proteins can
also be separated by their Ip as they are less soluble in their zwitterion form of zero charge
allowing them to be precipitated out. Polar solvents like acetone and alcohol will remove the
bound water from protein and aid in separation. Another form of approach is sodium dodecyl
sulfate polyacrylamide gel electrophoresis (SDS-PAGE) that separates protein based on their
MW. The SDS is an anionic detergent that makes proteins negatively charged by attachment and
denaturing the proteins to allow separation solely based on size of the fragments. All fragments
move towards positive electrode with heavier fragments moving slower than the lighter
fragments (Al-Tubuly, 2000). The proteins are most commonly isolated through column
chromatography techniques such as ion-exchange chromatography in which different
immobilized resins can be used to isolate proteins based on their charge. Diethylaminoethyl
(DEAE) a positively charged resin can be used to elute out negatively charged bounded proteins
by increasing salt concentration and displacing those proteins. Cation exchanger
carboxymethylcellulose (CMC) is negatively charged and is used for elution of positively
charged proteins (Coskun, 2016). Biuret assay is used for testing presence of the peptide bonds
where peptides bonds react with the Cu
2+ ions in alkaline solution forming a violet complex.
Nitrogen’s in peptides form coordination bond with the metal ion. The number of bonds formed
depends on the number of peptide bonds. The proteins is then measured at 540nm to form
calibration curve to find protein concentration. It does not depend on amino acid composition of
protein and requires at least 1mg of protein and it has low sensitivity (Gornall, Bardawill and
David, 1949).
Objectives
This experiment is aimed to isolate, purify and characterize the lysozyme protein from egg white
sample as well as analyzing their biological significance. This is done through different
techniques by dividing experiment into three weeks. In week one the isolation of different
fractions of lysozyme is done through ion-exchange chromatography with CMC resin, pH
alteration by buffer, centrifuge and dialysis to obtain 4 fractions of lysozyme protein; A, B, C and
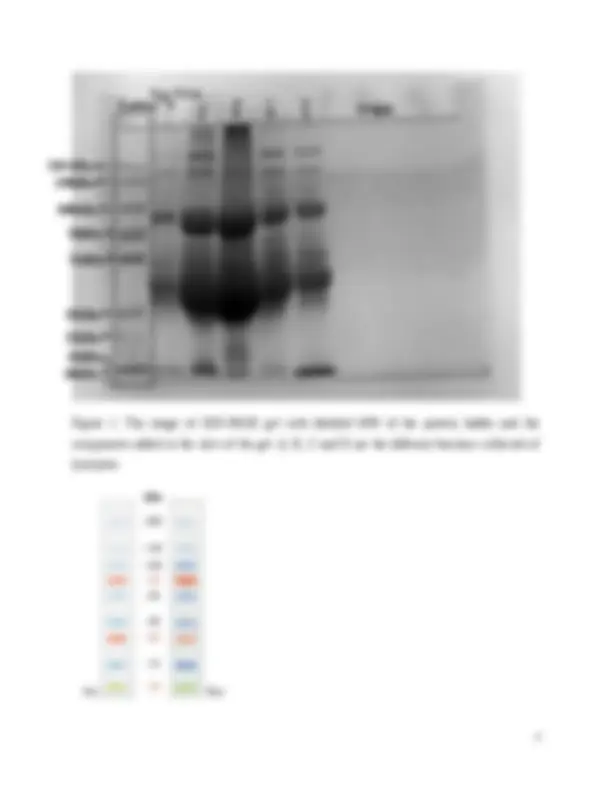
Figure 1: The image of SDS-PAGE gel with labelled MW of the protein ladder and the
components added in the slots of the gel. A, B, C and D are the different fractions collected of
lysozyme.
B C D
A
Egg White
Origin
10kDa
15kDa
25kDa
35kDa
55kDa
70kDa
100kDa
130kDa
250 kDa
Ladder
Figure 2: The MW of the protein ladder 26619 obtained from ‘Thermoscientific PageRuler Plus
Prestained Protein #26619’.
Table 2: The calculated distance travelled, the molecular weight and log of molecular weight for
the protein ladder #26619 used.
Band no.
Distance travelled
(cm)
Molecular weight
(kDa)
Log (molecular
weight) (kDa)
1 1.2 250 2.
2 1.6 130 2.
3 2.5 100 2.
4 3.3 70 1.
5 4.0 55 1.
6 5.1 35 1.
7 6.4 25 1.
8 7.2 15 1.
9 7.3 10 1.
[The distance measured through an enlarged image]
0.8 1 1.2 1.4 1.6 1.8 2 2.2 2.4 2.
0
1
2
3
4
5
6
7
8
f(x) = − 5.06 x + 12.
R² = 0.
Log of MW of protein ladder against the distance travelled by the bands
Log of molecular weight (kDa)
Distance travelled (cm)
Figure 3: The graph of the log of molecular weight of proteins ladder against its distance
travelled by the bands in cm.
[The distance travelled is measured through an enlarged image of gel] Sample calculation for the molecular weight of EW1 band: x = log of molecular weight, y = distance travelled by the fragment (cm)
- EW1 0.9 2.360 229. (kDa) - EW2 1.4 2.261 182. - EW3 2.8 1.985 96. - EW4 4.1 1.728 53. - EW5 4.9 1.570 37. - EW6 5.2 1.511 32. - EW7 5.4 1.471 29. - EW8 7.2 1.116 13. - EW9 7.3 1.096 12. - B - B - D - D - D - D - D - D - D - D - C - C - C - C - C - C - C - C - C - C - B - B - B - B - A - A - A - A - A - A - A - A - A - A - EW - EW - EW - EW - EW
- EW
- EW
- EW
- EW - A1 0.10 2.518 329. - A2 0.25 2.488 307. - A3 0.40 2.459 287. - A4 1.00 2.340 218. - A5 1.40 2.261 182. - A6 3.20 1.906 80. - A7 3.80 1.787 61. - A8 5.60 1.432 27. - A9 7.20 1.116 13.
- A10 7.40 1.076 11.
- B1 0.4 2.459 287.
- B2 1.4 2.261 182.
- B3 3.4 1.866 73.
- B4 5.9 1.372 23.
- B5 6.9 1.175 14.
- B6 7.4 1.076 11.
- C1 0.2 2.498 314.
- C2 1.0 2.340 218.
- C3 1.5 2.241 174.
- C4 1.7 2.202 159.
- C5 3.0 1.945 88.
- C6 3.6 1.827 67.
- C7 4.2 1.708 51.
- C8 5.3 1.491 30.
- C9 7.3 1.096 12.
- C10 7.5 1.056 11.
- D1 0.2 2.498 314.
- D2 1.0 2.340 218.
- D3 1.5 2.241 174.
- D4 2.8 1.985 96.
- D5 4.1 1.728 53.
- D6 5.0 1.550 35.
- D7 7.4 1.076 11.
- D8 7.6 1.037 10.
- y = -5.0639x + 12. From Figure 3,
- 0.9 = -5.0639x + 12.
x=
0.9−12.
−5.
=
Log of molecular weight for EW1 = 2.360 kDa
Estimated fragment size:
10
= 229.09 kDa
The above calculations are applied for all bands observed in the SDS-PAGE image.
Table 4: The mass and concentration of BSA and the volumes added before measuring
absorbance of BSA at 570nm by UV-Vis spectrophotometer.
Standard Mass
of
BSA
(mg)
BSA
concentra
tion (
mg / ml ¿
Volume
of BSA
added
(ml)
Volume of
water
added (ml)
Volume of
biuret
reagent
added (ml)
Total
volume
(mL)
Absorbance
measured at
570 nm
1 (blank) 0.0 0.0 0.0 1.0 4.0 5.0 0.
2 2.0 2.0 0.2 0.8 4.0 5.0 0.
3 4.0 4.0 0.4 0.6 4.0 5.0 0.
4 6.0 6.0 0.6 0.4 4.0 5.0 0.
5 8.0 8.0 0.8 0.2 4.0 5.0 0.
6 10.0 10.0 1.0 0.0 4.0 5.0 0.
Sample Calculation for concentration of BSA standard 2:
Concentration = mass/ volume
= 2.0mg/1ml
= 2.0mg/ml
Sample calculation for volume of BSA added and volume of water added for standard 2:
M 1
V 1
= M 2
V 2
Where M 1 is the initial mass and V 1 is the initial volume
M 2 is final volume and V 2 is the final volume.
M 1 = 10 mg, V 1 = x, M 2 = 2 mg, V 2 = 1 ml
Lysozyme A 0.150 1:10 28.90 2283.
Lysozyme B 0.572 undiluted 11.02 551.
Lysozyme B 0.152 1:10 29.29 1464.
Lysozyme B 0.019 1:100 36.61 1830.
Lysozyme C 0.411 undiluted 7.919 269.
Lysozyme C 0.048 1:10 9.249 314.
Lysozyme D 0.172 undiluted 3.314 99.
Lysozyme D 0.026 1:10 5.010 150.
Egg white 0.625 1:3 36.13 570.
Egg white 0.115 1:30 66.47 1050.
Sample calculation for Protein concentration in lysozyme A 1:10 dilution sample:
From figure 5; y=0.0519x where y= absorbance and x = concentration (mg/ml)
Absorbance for 1:10 of lysozyme A is 0.
0.150=0.0519x
x= 0.150/0.
x= 2.89mg/ml x10 (dilution factor) = 28.90mg/ml in 1ml
Sample calculation for total protein mass in lysozyme A 1:10 dilution sample:
Concentration in 1ml = 28.90mg/ml
Mass in 1ml= 28.90mg/ml x 1ml
= 28.90mg
Total protein mass of lysozyme A = 28.90mg x 79ml (total volume from table 1)
= 2283.2mg
Method of calculations above apply to all the fractions and egg white diluted and undiluted
samples in the table above.
Week 3:
Table 6: The absorbance of the four lysozyme fractions and egg white sample measured at
500nm using UV-vis spectrophotometer for 2 mins of 10 seconds interval.
Time
(seconds)
Absorbance
of Lysozyme
A
dilution) at
500nm
Absorbance
of Lysozyme
B at 500nm
Absorbance
of Lysozyme
C at 500nm
Absorbance
of Lysozyme
D (1:
dilution) at
500nm
Absorbance
of Egg white
(1:
dilution) at
500nm
0 0.348 0.348 0.348 0.348 0.
10 0.180 0.333 0.343 0.164 0.
20 0.137^ 0.314^ 0.339^ 0.129^ 0.
30 0.125 0.304 0.318 0.120 0.
40 0.103 0.288 0.314 0.102 0.
50 0.084 0.287 0.305 0.085 0.
60 0.080 0.273 0.296 0.083 0.
70 0.072 0.257 0.289 0.078 0.
80 0.074 0.252 0.286 0.069 0.
90 0.074^ 0.254^ 0.280^ 0.067^ 0.
100 0.069 0.244 0.270 0.063 0.
110 0.063 0.232 0.263 0.059 0.
120 0.060^ 0.218^ 0.256^ 0.059^ 0.
Lysozyme B 1.8× 10
Lysozyme C
5.5× 10
Lysozyme D
(1:
dilution)
0.0186 1.
22.32 4.455 669.
Egg white
(1:
dilution)
0.0148 0.
17.76 0.267 280.
[Mass in 1ml is taken from table 5, egg white dilution 1:30 protein concentration and total
protein mass used and undiluted values for lysozyme B and C calculations in this table]
Sample calculation for initial slope of lysozyme A:
Slope= rise/run
= (0.35-0.18)/10s = 0.017abs unit/s
Sample calculation for enzyme activity of lysozyme A:
Enzyme activity = 0.017 x 60s = 1.02 abs unit/min in 1:10 diluted sample
Sample calculation for enzyme activity min/ml in lysozyme A:
Enzyme activity in 1:10 dilution = 1.02 abs unit /min
Originally = 1.02 x 10 = 10.2 abs unit/min
Lysozyme activity per ml = 10.2/ 0.5ml
= 20.4 abs unit/min/ml
Sample calculation for specific enzyme activity (abs unit/min/mg) of lysozyme A:
Specific enzyme activity =
enzyme activity
total proteinmass ∈ 1 ml
=
= 0.706 abs unit/min/mg
Sample calculation of total activity of lysozyme A:
Total protein mass of lysozyme A 1:10 sample after correcting it to its original value is 2283.2mg
Total activity = specific enzyme activity x total protein mass
= 0.706 x 2283.3 = 1612 abs/min
The method of calculation is applied to all other fractions and egg white sample in the table
above
Lysozyme purity
Table 8: The tabulation of lysozyme purity readings for all lysozyme fractions compared to egg
white.
Fraction Protein
concentrati
on (mg/ml)
Volume of
fraction
(ml)
Total
protein
(mg)
Specific
enzyme
activity
(abs/min/
mg)
Total
enzyme
activity
(abs/min)
Percenta
ge
recover
(%)
Fold purity
Lysozyme
A
A
10.87 79 858.5 0.706 1612 81.74 2.
Lysozyme
B
B
11.02 50 551.1 0.0196 10.80 52.50 0.
Lysozyme
C
C
7.919 34 269.3 8.33x
Lysozyme
D
D
3.314 30 99.4 4.455 669.6 9.46 16.
Egg white 66.47 15.8 1050.3 0.267 280.4 100 1
Sample calculation for percentage recovery of lysozyme A:
% recovery =
total proteinmass of egg
total proteinmass of fraction
¿
¿
=
x 100
= 81.74%
and be collected as pellet after centrifuge. Hence the high percentage recovery of 52.50%
lysozyme B with presence of other egg white proteins in the fraction.
The pellet from centrifuge is treated with 35ml of bicarbonate buffer to allow further aggregation
of proteins of selected proteins according to their pI and the pH of buffer and the solution
undergoes centrifuge again with supernatant providing lysozyme C fraction. The percentage
recovered 25.64% is lower than lysozyme A and B as most of the egg white proteins have been
collected in fraction A and B. This lysozyme C fraction again is contaminated with other egg
white proteins so further isolation is done to extract lysozyme from the pellet. Sodium carbonate
buffer (pH 10.4) is added to CMC residue and centrifuged to obtain lysozyme supernatant that
undergoes dialysis against running tap water to remove the excess alkali. After that the sample is
used as fraction D of lysozyme. This should ideally provide 3.5% theoretical yield but
experimentally 9.46% was recovered. This may not only be due to contamination by other egg
white proteins but could also be accounted to the presence of the traces of other components of
the egg white such as carbohydrate and lipids. Additionally some of the original egg white
sample in week 1 was spilled that may have contained lysozyme and thus this percentage
obtained may consist more of the other contaminant proteins than the interest protein lysozyme.
During the experiment some of the pellet from centrifuged suspension may also had been
transferred to sample with the supernatant when collecting lysozyme B and C fractions, leading
to increased contamination and high percentage recovery and inaccuracy of the results. Avidin
protein has pI of 10 (Abeyrathne, Lee and Ahn, 2013) so it may also contaminate the lysozyme D
as it will attach to CMC resins and be collected in lysozyme D fraction.
2-Comment on SDS-PAGE separation of lysozyme A,B,C and D – which fraction is pure? The MW
of the purest lysozyme and compare the experimental value with theoretical MW.
SDS-PAGE uses polyacrylamide cross-liked with the N,N dimethyl bis-acrylamide to separate
proteins and nucleic acids according to their molecular weights. The pore size of the gel can be
manipulated to allow larger samples and smaller samples to be separated from each other. This is
possible by alternating the concentration of the acrylamide and bis-acrylamide. The size of the
pores decrease with increasing acrylamide concentration after polymerization of the acrylamide.
Prior to addition of protein samples into the gel slots, the samples are treated with SDS to
denature proteins into same linear shape and allow all proteins to be negatively charged so
eliminate the influence of factors like length, charge and shape from effect the efficiency of the
results. The SDS is an anionic detergent that attaches to the proteins making them all negatively
charged and denaturing by the three-dimensional structure of proteins by disrupting the non-
covalent bonds, hydrophobic bonds, ionic bonds and the hydrogen bonds. Doing so allows the
proteins to be separated based solely on their molecular weights. All the negatively charged
proteins put through the PAGE allow them to migrate towards the positive electrode with heavier
protein moving slower and lighter proteins moving faster (Al-Tubuly, 2000). Estimation of the
distance travelled by represented bands of the proteins allow approximate MW of proteins to be
calculated when compared to a ladder with known molecular weights of bands. The weight of the
representing bands can be compared to the theoretical weight of the interest protein to determine
the purity of the sample being tested. The closer the measured weight of the band is to the
theoretical weight the pure the interest protein sample will be.
The theoretical MW of lysozyme is 14.4 kDa (Abeyrathne, Lee and Ahn, 2013). The MW of the
bands from the 4 fractions of lysozyme and egg white closest to the theoretical MW of lysozyme
is lysozyme A9 13.05kDa, lysozyme B5 14.96kDa, lysozyme C9 12.47kDa, lysozyme D
11.92kDa and egg white EW8 13.06kDa. The difference in the MW of the fractions and egg
white can be due to the subjectivity of the measuring of this MW by the distance travelled by the
bands, the scale used to determine the MW, the enlargement of the image in order to determine
the MW and the smearing and overlapping of the bands that further complicate the estimation of
the molecular weight by adding inaccuracy in the results. The smearing of the band occurs when
too much sample of the protein is loaded into the wells of gel or the sample is too concentrated
and requires dilution. The samples may also contain many proteins that have similar range of
molecular weight leading to overlapping bands (Mikkelsen, Cort and Corton, 2016).
The lysozyme D and egg white results on the gel are the only ones with least amount of smearing
and overlapping of the bands so are assumed to be the purest for estimation of MW of lysozyme
protein. The egg white has band EW9 MW of 13.06kDa which is closer to the theoretical value
than the 11.92kDa of lysozyme D7 band. Although D7 is clearer than EW9 so it must have
higher lysozyme content than the other. Lysozyme A and C fraction had the highest number of
bands viewed on PAGE that could be due to the presence of more of the other egg white proteins
in the fractions compared to the other fractions and the egg white sample. Though handling of
above the pI of the protein the protein carries a net negative charge and if the buffer pH is below
the pI, the protein carries a net positive charge. The IEC technique makes use of the amphoteric
nature of proteins to manipulate the charge on the protein in order to isolate it. When a positively
charged ion-exchanger is used in the column where the solution is run through and the protein
sample is supplied with buffer of pH above the pI of the protein, the proteins become negatively
charged. When these negatively charged protein sample is run through the column with
immobilized positively charged resins such as DEAE, the negatively charged proteins attach to
the opposite charged resins while the positively charged proteins are eluted out first. The
negatively charged proteins are then treated with high ionic strength counter-ions (such as Cl
that can displace the proteins so the interest fraction of proteins can be collected. A negatively
charged ion-exchanger such as CMC cation-exchanger allows isolation of positively charged
proteins. After the attachment of positive proteins on to the CMC the negatively charged proteins
are eluted out and the column is treated with high ionic strength ions such as Na
or K
that
displace the immobilized positively charged proteins and allowed them to be eluted out and
isolated. Thus IEC makes use of the affinity of the proteins and their charge in order to separate
them (Coskun, 2016).
According to Abeyrathne, Lee and Ahn (2013) the pI of lysozyme is 10.7, ovalbumin pI is 4.5,
ovotranferrin is 6.1 and avidin is 10. All egg white proteins except avidin and lysozyme have pI
within 3.9-6.1. The buffers used in the experiment are sodium bicarbonate buffer with 9.0 pH
and sodium carbonate buffer with 10.4 pH. In the presence of sodium bicarbonate buffer the
lysozyme becomes positively charged and attaches to the negatively charged resins CMC in the
flask with the buffer. So it is collected as pellet after centrifuge. When treated with sodium
carbonate buffer of pH 10.4 the lysozyme proteins are displaced and undergo centrifuge to be
collected as supernatant and put through dialysis to achieve pure sample of lysozyme and remove
excess alkali.
5-Explain the underlying mechanism of the turbidimetric assay for lysozyme activity – measure the
clearing of suspension of dead bacterial cells when their cell wall break down.
A turbidimetric assay is used to quantify bacterial density by the lysozyme activity on the
bacterial cell walls (Li, Nix and Schentag, 1993). Gram positive bacteria have thick
peptidoglycan layer in their cell wall that surrounds the single lipid membrane of bacterial cell.
Gram negative bacteria have a thin layer of peptidoglycan between the cytoplasmic and outer
lipid membrane. Disruption of bacterial cell wall can help fight against the invasion of these
pathogenic bacteria. Lysozyme hydrolyzes the 1,4-β-glycosidic linkages of N-acetylmuramic acid
(NAM) and N-acetylglucosamine (NAG) in the peptidoglycan layer of the bacteria that provides
protection and structural integrity to bacterial cells. The lysozyme binds to the glycan strands of
repeated disaccharide motifs NAM-NAG through its two active sites to the binding sites within
prominent cleft between the two domains in the peptidoglycan layer producing oxo-carbonium
intermediate. (Primo et al. 2018). In this experiment M. lysodeikticus a gram positive bacteria is
used to investigate the lysozyme activity. The absorbance of the fractions measured through
spectrophotometer at 500nm in the presence of this bacteria allows estimation of the lysozyme
activity. Lysozyme fraction D is diluted 1:10 and has the lowest absorbance reading of 0.059 at
two minutes that implies it has contains the highest amount of lysozyme in its suspension.
Lysozyme A 1:10 diluted has the second most with absorbance of 0.060 and lysozyme B and C
are undiluted showing 0.218 and 0.256. Lysozyme C has the least amount of lysozyme activity
hence its concentration is lowest in lysozyme C fraction. Egg white 1:10 diluted sample has
absorbance of 0.150 at two minutes so it has moderate amount of lysozyme present in it.
Therefore, slow decrease in absorbance of lysozyme B, C fractions and egg white imply low
lysozyme activity in the samples. Lysozyme A and D have the highest lysozyme activity and the
clearance rate so they contain high lysozyme concentration. Lysozyme contributes to the degree
of optical clearance of suspension.
During the 2 minutes of absorbance recording for each fraction of lysozyme and egg white in 10
seconds intervals, the absorbance decreases with the increasing time for all samples. The lysis of
the bacterial cell wall decreases the turbidity and the absorbance readings, as less molecules are
detected with increasing time in the suspension of bacteria and lysozyme fraction. The
decreasing absorbance is an indicator for the active lysozyme activity. Thus it can be said that it
is present in all the fractions with varying concentration. The clearance rate can also be measured
to identify the concentration of the lysozyme present in each fraction (A-D) and the egg white
sample. Lysozyme found in human fluids like tears, mucous and saliva acts like an antibiotic in
human body as it can hydrolyze mucopolysaccharides of the gram-positive bacterial cell walls.
Lipopolysaccharides (LPS) present in the gram-negative bacterial cell walls induce a strong
immune response in the body. The different composition of the gram-negative bacterial cell wall