Download The Role of Macrophages and the Innate Immune System in Inflammation and Immunity and more Study notes Biotechnology in PDF only on Docsity!
The Innate Immune System LB Nicholson F [email protected]
Lecture 2. Recognition and effector mechanisms (II) Integration
Lethal Toxin Dimeric toxin from anthrax delivering a proteolytic subunit to the cytosol C57BL/6 macrophages resistant, 129/S1 macrophages sensitive Single dominant locus on Ch Currently 5 different alleles in 18 mouse strains ( resistant; 3 susceptible) Protein is Nalp1b Caspase 1 deficient cells are protected
The inflammasome Inflammation involves the co-ordinated upregulation of a number of genes These are co-ordinated by specific transcription factors, particularly NF-κB Multiple different ‘danger’ signals are co-ordinated to a common final pathway This common final pathway has been called the inflammasome Immunity 27: 549-559 (2007) Current Opinion in Immunology 2007, 19:615–
Multiple activators for the inflammasome through Nalp and Nalp Common activation pathway leading to IL-1β secretion Genetic defects lead to inherited inflammatory conditions Treatment with anti-IL1R is effective
Co-ordinating the innate immune response Several cells have roles in the interface between innate and adaptive immunity. Neutrophils and macrophages are the predominant cell type in early inflammation. Here we focus on the macrophage, but also include some data from experiments on DCs.
Macrophages integrate innate immune signals
Extracellular signals
- PAMPs/MAMPs
- Danger
- Complement, proteolysis of extra-cellular matrix, chemotaxis, etc. acting as endogenous adjuvants
Leading to cellular responses
- Receptor recruitment of neutrophils and then macrophages in early inflammation
- MyD88 adapter protein is a key signalling molecule
- Leads to NF-kB driven gene transcription
Macrophages Large mononuclear phagocytic cells. Phagocytosis was described by Elie Metchnikoff whose first experiment was to introduce a splinter into a starfish larvae and to observe next morning that it was surrounded by mobile cells. In 1891 he proposed that this was important in human inflammation and immediately came under severe and protracted attack by the humoralists, who believed that immunity depended on soluble factors.
Macrophages express class II MHC and are therefore professional APCs. The other professional APCs are DCs, which share overlapping lineage, but are more associated with the ability to initiate immune responses.
the number of leucocytes in the blood, even though there is also a decrease in the mean half-life of the circulating cells.
Most macrophages and neutrophils at the site of acute inflammation derive from the circulation, although there is some division in situ at later time points. What are the mechanisms that draw cells to sites of inflammation and keep them there?
Release of soluble activators effecting endothelium e.g. chemokines Upregulation of integrins/addressins on endothelial cells Signals to retain cells in tissues Signals to the bone marrow
Signals at site of inflammation
Signals Dual receptor
Single receptor
Diffusible
Cognate interactions + Costimulation + ECM interactions +/- Pathogens + PAMPs + Endogenous Innate immune system ligands
Danger signals + Cytokines + Chemokines + NO; superoxides + +
Macrophages are very responsive to environment. External environment controls the gene programs executed by macrophages.
There are different responses to activation by Th1 or Th cytokines
IFNg, IL-12 M1 phenotype; classical activation IL-4, IL-13 M2 phenotype; alternative activation
But different to T cells: No clonal burst Integrating many other environmental cues, such as stimuli from pattern recognition receptors Degree of reversibility is uncertain
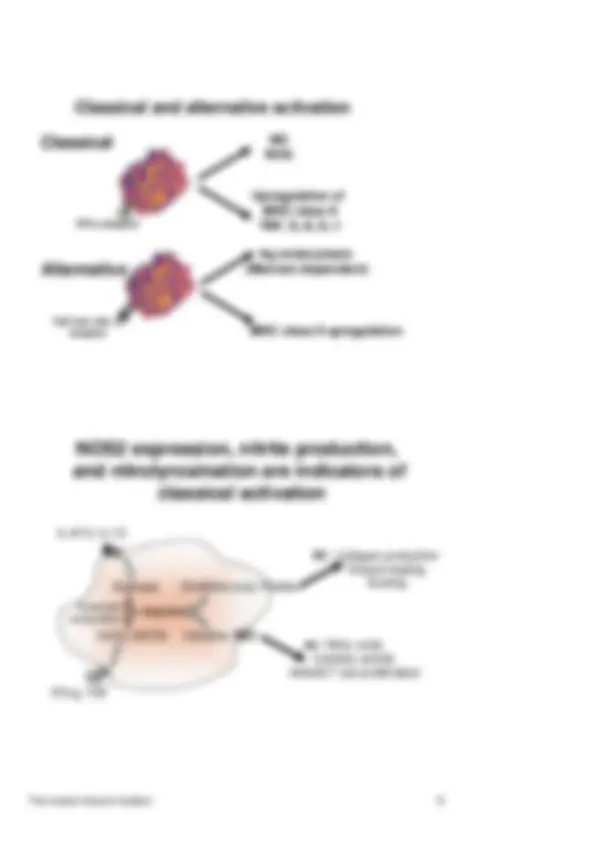
Classical and alternative activation
NO
ROS
Upregulation of MHC class II IFN- γ receptor TNF, IL-6, IL-
Classical
TGF-b/IL-4/IL- receptor
Alternative
MHC class II upregulation
Ag endocytosis (Mannan-dependent)
NOS2 expression, nitrite production,
and nitrotyrosination are indicators of
classical activation
Collagen production Wound healing Scaring
Nitric oxide Cytolytic activity Inhibits T cell proliferation IFN-g, TNF
IL-4/13, IL-
L-Arginine
Arginase
NOS2 (iNOS) Citrulline, NO
Substrate competition
OrnithineOrnithine ProlineProline
M
M
Innate immune signals induce IL-6 which ‘derepresses’ T regulatory cells
IL-
IFN-γ
IL-
IL-
TLRs (^) TLRs
Treg
Th
DC
Science. 299:1033-6, 2003
Innate immune signals in the presence of T regulatory cells which supply TGF-�, can lead to the development of T effector T cells which have potent pro-inflammatory effects.
Suppressing innate immune response can limit adaptive immune responses. In EAU, mice were treated with one or two doses of an antibody (BB5.1) that inhibits the proteolysis of C5. This treatment reduced ocular disease 14 and 21 days after immunisation.
Summary
It is increasingly recognised that innate immune signals play a role in setting the level of inflammation in a particular microenvironment. These signals act as natural ‘adjuvants’ which promote adaptive immune responses. This is good in the case of infection, but bad in the case of autoimmune disease.
Further reading:
S. Gordon. Alternative activation of macrophages. Nature Reviews Immunology 3 :23 - 35 2003 Nathan, C. Points of control in inflammation. Nature. 420 :846- 52 2002