Download by Ibrahim Cissé and more Lecture notes Cell Biology in PDF only on Docsity!
n the Cissé lab, we develop high resolution imaging
approaches, to go inside living cells and look at the scale
of single biological molecules. We are particularly interested in the
remarkable molecular machines that decode the genetic informa-
tion from DNA into RNA. Here we discuss our discovery that
these molecules interact transiently to exhibit liquid-like proper-
ties, and how these new insights are shaping our view of biological
phase transition as a means for the cell to regulate the expression
of our genetic material.
fifl��fi�����������
I
Capturing
Our
Genome
70 ) cissé mit physics annual 2020
in Action
mit physics annual 2020 cissé ( 71
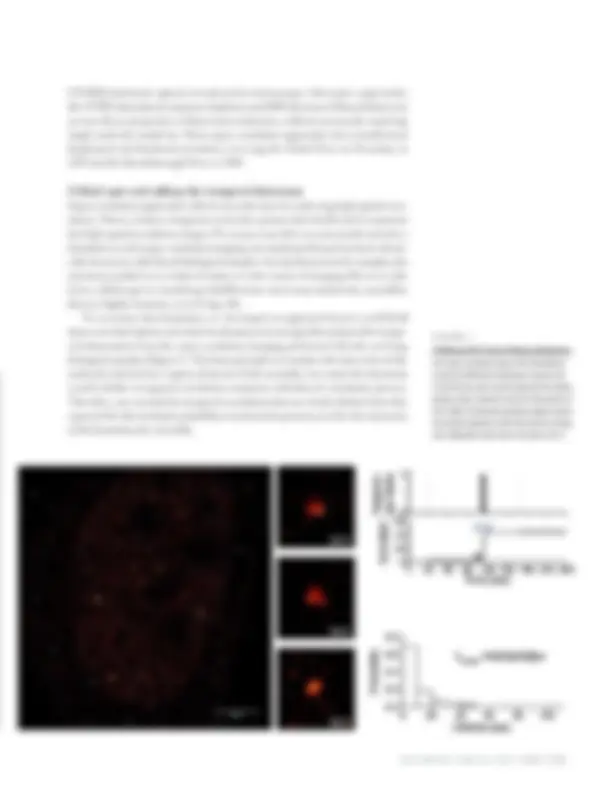
figure 1
tcPALM and Pol II cluster lifetime distribution. Left: Super-resolution image of Pol II distribution reveals sub-diffractive clustering in a human cell. A red-hot color code is used to represent the relative density. Center: Zoomed-in view of a few clusters of Pol II. Right: A temporal correlation analysis reveals the transient dynamics of the Pol II clusters in living cells. [Adaptation from Cissé et al Science 2013.] STORM (stochastic optical reconstruction microscopy). Alternative approaches like STED (stimulated emissions depletion) and SIM (structured illumination) rely on non-linear properties of fluorescent molecules, without necessarily requiring single molecule sensitivity. These super-resolution approaches have transformed biophysical and biochemical studies, receiving the Nobel Prize in Chemistry in 2014 and the Breakthrough Prize in 2018. A blind spot and adding the temporal dimension Super-resolution approaches effectively trade time for achieving high spatial reso- lution. That is, it takes a long time to localize points individually and reconstruct the high spatial resolution images. For many years this was not considered to be a limitation as such super-resolution imaging was mostly performed on dead, chemi- cally frozen (so-called fixed) biological samples. Or, if performed in live samples, the structures probed were relatively static over the course of imaging. However, this led to a blind spot in visualizing subdiffraction-sized macromolecular assemblies that are highly dynamic, as in living cells. To overcome this limitation, we developed an approach known as tcPALM (time-correlated photo-activation localization microscopy) that analyses the tempo- ral information from the super-resolution imaging performed directly on living biological samples (Figure 1). The basic principle is to analyze the time series of the molecules detected in a region of interest. If the assembly were static the detections would exhibit a temporal correlation consistent with that of a stochastic process. Therefore, one can look for temporal correlations that are clearly distinct from that expected for the stochastic pointillist reconstruction process, to infer the dynamics of the biomolecular ensemble.
mit physics annual 2020 cissé ( 73
Pol II clusters too short-lived In the first demonstration of tcPALM, we studied the dynamics of the enzyme Pol II, which is the molecular machine that is responsible for reading the information in DNA and converting it into a related polymer called RNA. This RNA molecule is a transient message that the cell uses to make proteins, which perform most of the work of the cell. Using tcPALM, we discovered that Pol II forms subdiffrac- tive clusters that previously went unnoticed in living cells imaged by conventional fluorescence microscopes. Pol II had been hypothesized to form clusters in previous studies, but those observations were often dismissed as a possible artifact of the harsh chemical fixation used in those earlier studies, as there had been no evidence for such clusters in conventional live cell imaging. In addition to revealing the exis- tence of such clusters in living cells, tcPALM could also measure their dynamics, and therein lay the surprise to us. Instead of being “stable factories,” as previously hypothesized, we found that clusters of about 100 Pol II molecules assemble and then disassemble within a few seconds. And while a few seconds could sound like a long time, it is orders of magnitude faster than the time scale of Pol II activity. Pol II is a complex molecular motor that assembles and tracks on DNA, unwinding the double helix and copying it one basepair (bp) at a time into RNA transcripts. The speed of Pol II is ~2,500bp per minute. A portion of DNA that Pol II copies is called a gene, and in mammalian (human and mice) cells that we study in our laboratory a typical gene size is about 10,000 bp. So it takes several minutes for Pol II to finish even one transcript. It was therefore unclear whether the very transient, seconds-long clusters had any significant role in the biological function of Pol II. However, given that this stems from studying a fundamental process at unprecedented spatiotemporal resolutions in living cells, we suspected that perhaps these puzzling transient cluster dynamics could be pointing to previ- ously hidden mechanisms of gene regulation. Dynamics predict the number of transcripts made We next sought to test the putative function of transient Pol II clusters in living cells. Toward this end we developed dual color super-resolution imaging in living cells. One color would be used to perform tcPALM on Pol II as we had done earlier, and the second color would be used to look at the RNA transcripts that are the product of Pol II activity at a specific gene of interest. This idea worked wonderfully (Figure 2). We were able to capture transient Pol II clustering, precisely at the locus of a gene that is actively being transcribed, and correlate those measurements directly with the gene output—that is, nascent RNA transcripts inside the living cells. We discovered that the dynamics of clustering predictively correlate with how many RNA copies are made. The clusters form in the early stages, as the gene transcription process is initiated. The cluster likely helps to increase the local concen- tration of Pol II enzymes at the gene to be transcribed. In a biochemical reaction, a high concentration increases the collision rate between enzyme and substrate.
74 ) cissé mit physics annual 2020
figure 3
Mediator and Pol II colocalization. Left: Multiple cells can be seen in the field of view showing that Mediator clusters (magenta) co-localize with Pol II (green) clusters. Right panels: Zoomed-in view of Pol II & Mediator clusters colocalization (green
- magenta = white indicates the two signals overlap). [Adaptation from Cissé Lab ms.: Cho et al Science 2018.] Pol II partner, Mediator, also clusters To understand how the clusters form and what could control their dynamics, we looked at the molecular partners of Pol II. One such partner is the so-called Media- tor complex. Mediator binds at regions on the DNA that are called an enhancer , which are distant from where Pol II binds and starts transcribing (which is called a promoter ). The DNA is then believed to loop around so that the Mediator can directly contact Pol II. But when these models of Mediator – Pol II contacts were developed, it was believed that only a single Pol II complex would be present, not a cluster of 100 Pol II. We therefore wondered whether, in fact, Mediator also forms clusters similar to the Pol II clusters. We found that indeed Mediator forms clusters similarly to Pol II. In fact, Mediator and Pol II clusters co-localize in the same loci in living cells (Figure 3). Therefore, the dynamic clustering we discovered with Pol II may be general to many other regulatory molecules beyond Pol II.
76 ) cissé mit physics annual 2020
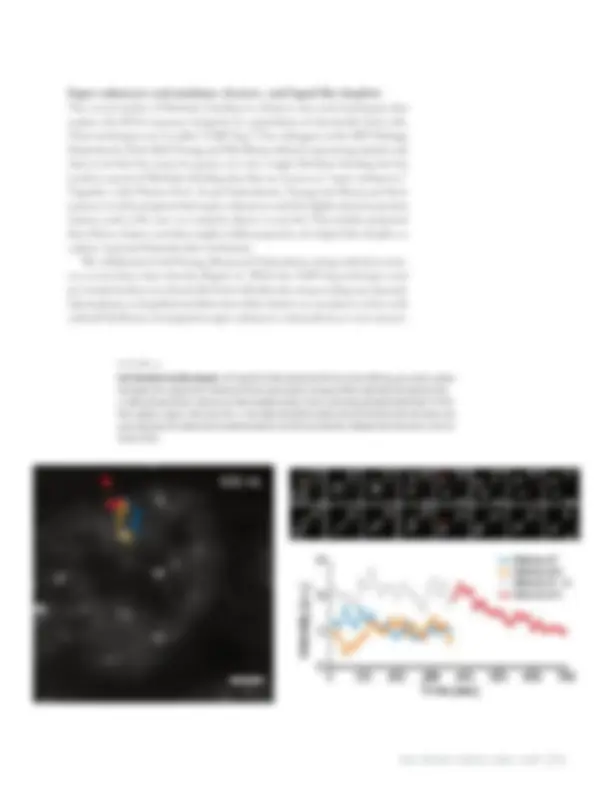
figure 4
Pol II-Mediator fuse like droplets. Left: Snapshot of video showing that the two clusters diffusing, upon contact, coalesce like droplets into a single cluster. Trajectory of the two clusters drawn in orange and blue respectively; the trajectory of the co-diffused merged cluster is drawn in red. Video available at https://science.sciencemag.org/highwire/filestream/711819/ field_highwire_adjunct_files/3/aar4199_s1.mov. Right: Quantitative analysis shows the intensity of the two clusters, and upon coalescence the merged cluster instantaneously has sum of the two intensities. [Adaption from Cissé Lab ms.: Cho et al Science 2018.] Super-enhancers and mediator clusters, and liquid-like droplets The recent studies of Mediator binding at enhancer sites used techniques that analyze the DNA sequence footprint in a population of chemically fixed cells. These techniques are so-called “ChIP-Seq.” Our colleagues in the MIT Biology Department, Profs. Rick Young and Phil Sharp, utilized sequencing analysis and discovered that for many key genes, it is not a single Mediator binding site but tandem repeats of Mediator binding sites that are known as “super-enhancers.” Together with Physics Prof. Arup Chakraborty, Young and Sharp and their trainees recently proposed that super-enhancers could be highly dynamic protein clusters, such as the ones we routinely observe in my lab. They further proposed that if these clusters exist they might exhibit properties of a liquid-like droplet, or a phase-separated biomolecular condensate. We collaborated with Young, Sharp and Chakraborty, along with their train- ees, to test these ideas directly (Figure 4). While the ChIP-Seq technique used previously had been in chemically fixed cells (thereby not providing any dynamic information), we hypothesized that some of the clusters we can observe in live cells with tcPALM may correspond to super-enhancers. And indeed, we were success-
mit physics annual 2020 cissé ( 77
living cells. Biomolecular condensates are in fact becoming ubiquitous (Figure 5) in many different aspects of cell function besides Pol II transcription. And there are likely general physical mechanisms that govern these phase transitions inside living cells. We recently discovered that classical nucleation theory describing the conden- sation of a system in a steady state, super-saturation system describes how some diffraction-sized liquid-like condensates form and grow in living cells. Classical nucleation theory: a first order phase transition mechanism for biomolecular condensates formation in living cells Briefly, classical nucleation theory describes how a super-saturated system, say water vapor at high relative humidity, condenses into liquid droplets. It predicts that small clusters or drops will spontaneously form but rapidly dissolve due to a high surface tension. If the vapor concentration is higher than a critical (saturation) concentration, there emerges a maximum in the free energy, that is a critical size above which a nucleating cluster is stable enough to grow spontaneously. However, such a free energy is thought nearly impossible to measure experimentally: that is because the super-saturated state is a very transient metastable state. As soon as a few clusters reach the critical size, the clusters rapidly absorb monomers to grow, the ambient vapor concentration decreases rapidly, and the free energy maximum vanishes. Nonetheless, we suspected that with our high spatiotemporal resolution in living cells we might have a shot at capturing the small nucleating clusters, and perhaps measure for the first time directly the free energy of a super-saturated system in the living cells. Szilard’s demon and steady state super saturation A toy model proposed by Hungarian-American physicist Leo Szilard suggested that super-saturation could be maintained in steady state, in theory, if one could imagine a “Maxwell’s demon” that surveys nucleating clusters and removes those that reach the critical cluster size. However, Szilard’s model was thought never to exist in natural systems where phase transitions were more commonly observed in physics. Nonetheless, we posited that in biological phase transitions one could conceive that, through evolution, living cells evolve such a Szilard demon as a regulation mechanism, especially in cases where large clusters can be detrimental to the cell function. We found this to be exactly the case in clusters associated with Parkinson’s disease, a neurological disease. We took a high-resolution snapshot of the clusters associated with Parkinson’s, thereby measuring the probability distribution of cluster sizes. From that distri- bution, which the theory predicts should be Boltzmann’s distribution for cluster sizes below criticality, we can measure the associated free energy function. To our surprise, the free energy was exactly as predicted in classical nucleation theory with a term driven by surface tension (which dominates at very small cluster sizes), balanced by a second term attributable to relative super-saturation of the system.
mit physics annual 2020 cissé ( 79
We could therefore measure the energy barrier for nucleation (7 kBT); the critical cluster radius (160nm); bounds for the surface tension, and more. Importantly for disease research, we discovered that the pre-nucleating cluster is in fact a liquid- like biomolecular condensate, and we uncovered Szilard’s mechanism, involving a protein that seems to preferentially remove clusters above the critical size (Figure 6). Moving ahead Armed with the excitement that such biomolecular phase transition mechanisms can be quantitatively addressed by our experimental approaches, we are hopeful that we can ultimately uncover the hidden steps by which Pol II condenses into clusters, and how cells might have evolved to regulate these clusters. We suspect this is a place where cells have evolved to utilize a hidden and complex mixture of physics, biology and chemistry to control one of the most fundamental processes of life: decoding the hereditary information we inherit in the form of DNA.
figure 6
Classical nucleation theory. Left: Theoretical Free Energy function for a condensation (droplet formation) of a super-saturated system in first order phase transition. Right: Data from quantitative high-resolution microscopy of the clusters in human cells show precisely the scaling and free energy function expected from theory. This allows for the first time a direct read out of nucleation barrier and critical cluster size, which have been difficult to in a laboratory setting in other natural, phase transition systems (e.g., outside of biology). [Adaptation from Cissé Lab ms.: Narayanan et al eLife 2019.]
80 ) cissé mit physics annual 2020