Download Enzyme kinetics in downstream processing. and more Assignments Biochemistry in PDF only on Docsity!
1
Lecture 3: Enzyme kinetics
Fri 19 Jan 2009
Computational Systems Biology
Images from: D. L. Nelson, Lehninger Principles of Biochemistry, IV Edition, W. H. Freeman ed. A. Cornish-Bowden Fundamentals of Enzyme Kinetics, Portland Press, 2004 A. Cornish-Bowden Enzyme Kinetics, IRL Press, 1988
2
Summary:
- Simple enzyme kinetics
- Steady-state rate equations
- Reactions of two substrates
- Inhibition of enzyme activity
- pH dependence
- Biological regulation of enzymes
4
Basics
- Enzyme kinetics studies the reaction rates of enzyme-catalyzed
reactions and how the rates are affected by changes in experimental
conditions
- An essential feature of enzyme-catalyzed reactions is saturation : at
increasing concentrations of substrates the rate increases and
approaches a limit where there is no dependence of rate on
concentration (see slide with limiting rate V max)
- Leonor Michaelis and Maud Menten were among the first scientist to
experiment with enzyme kinetics in a “modern” way, controlling the pH
of the solution etc.
- The convention used for this slides is to use UPPERCASE for the
molecular entity: e.g. E is an enzyme molecule and italics lowercase for
the concentration: e.g. e 0 is the enzyme concentration at time zero
(initial concentration). Also square brackets can be used for
concentration, e.g. [E] = enzyme concentration.
For additional material: Fundamentals of Enzyme Kinetics, Athel Cornish-Bowden, 2004 or
Enzyme Kinetics, Athel Cornish-Bowden and C. W. Wharton, IRL Press, 1988
5
A simple view: E+A = EA as an equilibrium
- The mechanism: the first step of the
reaction is the binding of the substrate
(A) to the enzyme (E) to form and
enzyme-substrate complex (EA) which
then reacts to give the product P and
free enzyme E
- The concentrations: the total initial
enzyme concentration is e 0 , and the
complex concentration is x. The
substrate (A) concentration should too be
(a 0 - x) , but since the substrate
concentration is usually very high:
- The conversion is considered an
equilibrium with equilibrium constant K s
- The slowest step of the reaction is the
EA to E+P and hence the dominant rate
for all the reaction is the rate v = k 0 x or,
considering the form 3.
(e x) a x p
0
0
s
E P
k
EA
K
1. E A
a a 0 ( a 0 x )
(K )
or 3.
( )
[EA]
[E][A]
- K
s
0 0
s
a
e a
x
x
e x a
(K )
k
- v
s
0 0
a
e a
7
The steady-state assumption
- When the enzyme is first mixed with a large
excess of substrate there is an initial period,
lasting just a few microseconds, the pre-
steady state , during which the EA complex
concentration builds up
- The reaction quickly achieves a steady state
in which [ES] remains approximately constant
over time (Briggs and Haldane, 1925)
For additional material: Fundamentals of Enzyme Kinetics, Athel Cornish-Bowden, 2004 or
Enzyme Kinetics, Athel Cornish-Bowden and C. W. Wharton, IRL Press, 1988
8
A more general view: E+A EA as reversible
- The mechanism: the first step is now
reversible with a forward (k 1 ) and a
backward (k (^) - 1 ) constant
- The rate of change of the
concentration of intermediate (dx/dt)
is the difference between the rate at
which it is produced from E + A and
the sum of the rates at which it is
converted back into free E and A and
forward into free E and P
- Briggs and Haldane postulated that
dx/dt should be positive at the instant
of mixing of E with A (because at the
beginning there is no EA and then it builds
up) but then the removal rate of EA
would rapidly increase, until it
balances the rate of production. This
is the steady-state
(e x) a x p
E P
k EA
k
k
5. E A
0
0
1
k e x a k x k x
x
1 ( 0 ) - 1 - 2
dt
d
-
( ) - - 0
dt
d
- k 1 e 0 x a k 1 x k 2 x
x
10
- When a is much smaller than K m it can be ignored giving:
with first-order dependences on both enzyme and substrate,
or second-order kinetics overall.
- k 0 /Km is thus called the second order rate constant or, more
importantly, the specificity constant , as it is specific for each
enzyme type.
- As a increases and surpasses K m making it insignificant, 8.
becomes:
- Where V is the limiting rate (better definition than “V max ”, since the value is approached rather slowly, and never really reached)
- You can experiment with this equation on: http://bcs.whfreeman.com/lehninger/ clicking on Chapter 6: Enzymes “Living graphs” menu
The Michaelis-Menten equation:
effects of substrate concentration on a reaction maximum velocity
Images from: David L. Nelson, Lehninger Principles of Biochemistry, IV Edition, W. H. Freeman ed.
e a
k
0
m
0
)
K
- v (
- v k 0 e 0 V
11
Lineweaver-Burk equation
- The Lineweaver-Burk equation
is a rewriting of the Michaelis-Menten
equation that is often used for the (not
very accurate) determination of kinetic
parameters from the plot.
- It is useful for analysis of multi-
substrate and inhibited enzymatic
reactions (see next slides)
- You can experiment with this equation on:
http://bcs.whfreeman.com/lehninger/ clicking on
Chapter 6: Enzymes “Living graphs” menu
a
Km
V V
1
v
1
-
Double reciprocal plot:
Biochemistry, IV Edition, W. H. Freeman ed. Images from: David L. Nelson, Lehninger Principles of
13
Reactions of two substrates
14
- The most common enzyme mechanism involves a chemical group transfer
from one substrate to another.
There are multiple possibilities to be considered:
(A)
- Does the reaction involve transfer of the group from the donor (first substrate) to
the enzyme, followed by a second transfer from the enzyme to the acceptor
(second substrate)? [ Ping-Pong mechanism]
- Or does the transfer occur in a single step while both donor and acceptor are still in
the active site of the enzyme? [ Ternary complex]
(B)
- In this second case, is the order of binding of donor and acceptor random?
[ Random-order ]
- Or is it compulsory that one of the substrates enters the active site first?
[ Compulsory-order ]
- And, can we distinguish the mechanism from the kinetics properties of the
enzyme? Yes.
Types of enzyme mechanism for
reactions of two substrates
16
Two substrates mechanism: Ping-Pong
Images from: David L. Nelson, Lehninger Principles of Biochemistry, IV Edition, W. H. Freeman ed.
- Parallel lines at increasing concentrations of the second
substrate ([S 2 ])indicates a Ping Pong mechanism.
17
Reversible inhibition
of enzyme activity
19
Uncompetitive Inhibition (not very
common)
- An uncompetitive inhibitor (I) binds
at a site distinct from the substrate
active site
- You can experiment with this equation on:
http://bcs.whfreeman.com/lehninger/ clicking on Chapter
6: Enzymes “Living graphs” menu
Images from: David L. Nelson, Lehninger Principles of Biochemistry, IV Edition, W. H. Freeman ed.
20
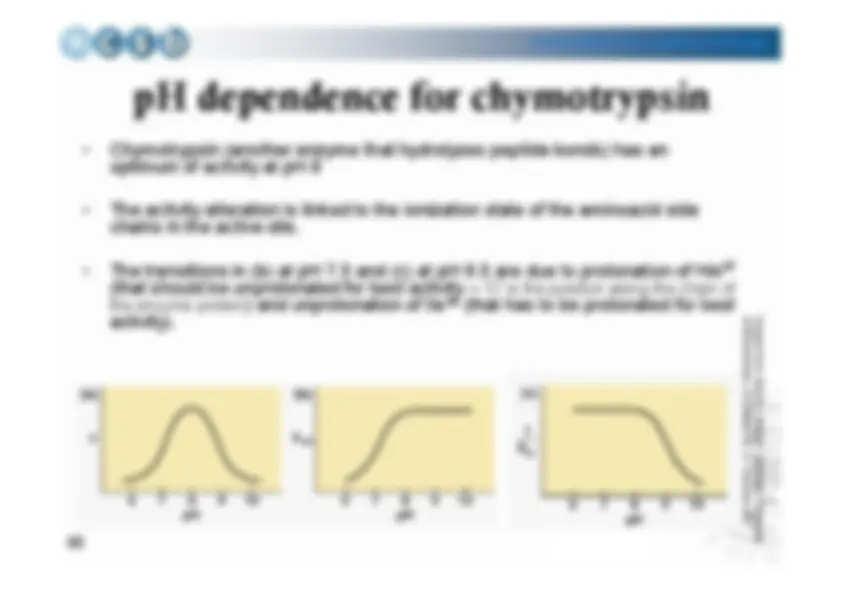
pH dependence
of enzyme activity