Download extraocular muscles: development, histologic features and more Summaries Logic in PDF only on Docsity!
488
THE ORIGINS AND INSERTIONS OF THE
EXTRAOCULAR MUSCLES:
DEVELOPMENT, HISTOLOGIC^ FEATURES,
AND CLINICAL SIGNIFICANCE*
BY David Sevel, MD, PhD
INTRODUCTION
THE TENDONS^ OF ORIGIN AND INSERTION^ OF THE^ EXrRAOCULAR^ MUSCLES SERVE^ TO attach these muscles to the bony orbit and to the sclera. By convention, the term "tendon" is used with extraocular muscles, where aponeurosis would be more correct: A tendon implies a rounded or oval cord, whereas
an aponeurosis is broad, dense, flattened connective tissue that gives
wide and extended purchase to the annulus of Zinn posteriorly and the sclera anteriorly. "Tendon" will be used in this dissertation, in keeping with presently accepted ophthalmic terminology. Little research has been carried out with reference to the tendinous
origins and insertions of the extraocular muscles. Previous investigations
have been confined to gross measurements of the tendon's length, width, and site of insertion.^ 1- The (^) purpose of this report is to describe the (^) development and histo- logic features^ of the^ tendons of^ origin^ and^ insertion^ of^ the extraocular
muscles. The evolution of tendons, from early in embryonic life through
different stages of development to term, is examined, as is the source of the embryonic tissue from which tendons are derived. The unique man- ner ofattachment of the tendon to the periosteum and sclera is described.
The detailed embryologic and histologic features of extraocular muscle
tendons from early development to^ term are^ used^ to^ explain superior
oblique tendon sheath^ syndrome and^ superior rectus^ muscle underaction
associated with^ ptosis.
*From the Division of Ophthalmology, Scripps Clinic and Research Foundation, La Jolla, CA. TR. AM.^ OPHTH.^ Soc.^ vol.^ LXXXIV, 1986
Extraocular Muscles 489
PRESENT STUDY
MATERIALS AND METHODS
Sixty-four human eyes from embryos and fetuses, ranging in size from
13.6 mm to term, were examined (Fig 1). Between 60 and 110 micro- scopic sections were examined, respectively, from each embryo and fe- tus. The normal development of the embryo/fetus was an essential pre- requisite, so the conceptus of septic abortions was not accepted for inves- tigation. If the mother had been exposed to teratogenic drugs during pregnancy, the^ embryo or^ fetus^ was excluded from the survey. Where salpingectomy was done (for (^) ectopic pregnancy), the fallopian tubes were
dissected with the aid of a dissecting microscope and the embryos re-
trieved. Usually these embryos were found within blood clots.
Age of EmbryolFetus
The maternal history with reference to duration of pregnancy was (^) found to be unreliable. A more satisfactory correlation was found between the clinical size of the uterus and the age of the embryo/fetus. Where pos- sible, however, a correlation was attempted between the clinical age of the (^) embryo/fetus and its (^) crown-rump length. A micrometer gauge was 350 1, -^ T I I. ADULT
I
I- z L1 J
z
3.J u
3uu 0 250 0 0 200 0 150
100 .0 ___
_ /iz (^) _== 50 0 0 5 10 1 5 20 25 30 35 40 EMBRYOIFETAL (^) AGE (WEEKS) FIGURE 1 Crown-rump length (mm) and^ age (weeks) of^ embryos and fetuses examined.
Extraocular (^) Muscles 491
C <^ ><U^ J^ ~~~~cl'cII co^ cl'^ cln^ c3^ C's^6 C's
< 0U
-U s
o E
z 0
H -~Q;C ~~ C
o 3~~~~~coI I^ C60000K1nK^6 C'^ C- c _
-e=^ OKDO
D~~~~~c V) C- .C):~^1 '^ _
H~~~~~~~~~~
z1 3 cl'z n)^ cln^ V) V)^ V)^ m^ C n
0
H 3 m
z z^0
..>XK < m O n N K K: (^) ~bt
1<Xv) _~~~lClJC-1 1 coJ V)C,)D
CLINICAL INVESTIGATIONS
In the Department of Ophthalmology at the University of California, San
Francisco, 46 patients who had had surgical treatment (bilateral medial
rectus recession) for nonaccommodative congenital esotropia were as-
sessed, with^ special reference to the sites of insertion of the medial rectus
muscles from the limbus and (^) the correlation between these sites of inser-
tion and the degree of esotropia (Table II).
In addition) in the Department of Ophthalmology at the University of
Sevel
California, San Diego, and at Scripps Clinic, San Diego, 21 patients who had had surgical treatment (medial rectus recession and lateral rectus resection) for nonaccommodative congenital esotropia were evaluated to
determine the site of insertion and^ the^ degree of esotropia was ascer-
tained. The^ length and width^ of the^ tendons^ were^ measured^ (Table III).
Special care^ was^ taken^ to^ identify abnormal^ adhesions, thickened^ tissue,
or extramuscular slips at the insertion sites.
MORPHOGENESIS OF ORIGINS OF EXTRAOCULAR MUSCLES Development of Annulus of Zinn The tendons of the rectus muscles are first^ observed to be attached to the perichondrium at^ the^ apex of the orbit^ at 26^ mm^ (Fig 2). The rectus muscles and their corresponding tendons mature, and the perichondrium at their sites of origin thickens in a ring-like configuration. As the cartilage at the apex of the orbit develops into bone (225 mm), the perichondrium becomes periosteum; this ring of thickened periosteum is called the annulus of Zinn, which is located over the optic foramen and the medial aspect of the superior orbital fissure. The origin of the lateral rectus muscle has both superior and inferior heads that^ straddle^ the superior orbital^ fissure.^ These^ superior and infe- rior heads are first discernible in the 54-mm embryo. At their origins, the superior and medial rectus muscles fuse with the dura of the (^) optic nerve. The fusion of the medial rectus muscle to the anlage of the dura of the optic nerve is first observed in the 22-mm embryo (Fig 3). The superior rectus muscle is seen to merge with the dura of the optic nerve in the 70-mm fetus. At term, the superior portion of the annulus (tendon of Lockwood) is attached to the body of the sphenoid bone, straddles the superior orbital
fissure, and^ also^ attaches^ to a^ bony spur, the spina recti^ lateralis. The
superior rectus muscle and the superior portions of the medial and lateral rectus muscles originate at the tendon of Lockwood. The inferior portion of the annulus (tendon of Zinn) is attached to the lesser wing of the
sphenoid and to the infraoptic tubercle. The inferior rectus muscles and
part of the medial and lateral recti arise from the tendon of Zinn. The origin of the superior oblique muscle is continuous with the origin of the medial rectus muscle at the apex of orbit and is noted first in the 22-mm embryo. By term, with the^ development and enlargement of the
orbit, the^ tendinous^ origin of^ the superior oblique muscle sequestrates
from the annulus of Zinn and takes (^) origin from the (^) region of the fronto-
ethmoidal suture immediately superior and medial to the origin of the
medial rectus muscle.
492
Sevel
TABLE III: CORRELATION BETWEEN SITE OF INSERTION OF MEDIAL RECTUS MUSCLES AND DEGREE OF ESOTROPIA IN 21 PATIENTS TENDON MEASUREMENTS (mm)* PATIENT PATIENT MEDIAL RECTUS LATERAL RECTUS ESOTROPIA NO AGE (MO) (DIOPTERS) L W^ LIMB L W LIMB (^1 6 2) 4.0 3.0 4.5 5.5 3.0 60 (^2 7 2) 4.5 3.0 4.5 5.5 3.5 50 3 7 2.5 4.5 (^) 3.5 4.5 5.0 3.0 50 4 8 2.5 6.5 3.0 4.0 6.0 3.0 (^50) 5 9 2.5 5.5 3.0 4.0 5.5 3.5 60 6 9 2.0 5.5 3.0 5.5 5.5 3.5 40 7 10 2.5 6.0 4.0 5.0 6.0 3.5 45 8 10 2.5 6.5 3.0 6.0 6.5 4.0 50 9 10 3.0 6.5 3.0 5.5 6.5 4.0 50 (^10 11) 2.0 6.0 3.5 6.0 6.5 4.0 60 (^11 11) 2.5 7.0 3.5 6.5 7.0 4.0 50 12 12 2.0 (^) 6.5 4.0 6.5 6.0 4.5 60 13 12 2.0 8.0 4.5 (^) 6.5 6.5 3.5 50 14 12 2.5 8.0 5.0 (^) 7.0 7.5 5.5 40 15 12 3.0 8.0 5.0 6.0 6.5 4.5 (^40) 16 15 3.0 8.0 5.0 6.0 7.0 5.5 (^50) 17 18 3.0 8.5 5.0 7.0 6.5 5.5 45 18 18 2.5 8.5 5.0 7.5 7.5 6.0 40 19 18 3.0 8.5 5.5 8.0 8.0 5.5 55 20 24 3.5 9.0 5.5 8.5 8.5 6.5 60 21 24 3.5 9.0 5.5 8.5 8.5 6.5 60 *L, length; W, width; LIMB, site of insertion from limbus.
The origin of the inferior^ oblique muscle^ is^ attached^ to^ the^ periorbita
adjacent to^ the^ bony opening of the nasolacrimal duct. This^ origin is
muscular in the 38.2-mm embryo, becomes argyrophilic, and at term is
attached to the perichondrium and later the periosteum via a tendon
adjacent to the lacrimal fossa just posterior to the orbital margin.
The levator palpebrae superioris muscle, while playing no role in the
movement of the eye, is^ embryologically and^ developmentally closely
associated with the superior rectus^ muscle, and^ both muscles share^ a
common epimysium.
Histologic Features in Development of Origins of Extraocular Muscles
The features of developing muscle8-early myoblast, myoblast, multinu-
cleated myoblast, myotube (with longitudinal and cross-striations), and
mature muscle cells-are not observed in developing tendons. At the
proximal and distal ends of the extraocular muscles, mesenchymal cells
contain minimal cytoplasm, become elongated, and are more compact
than the cells of the muscle (36 mm).
Muscle endings are tapered, rounded, or split (Fig 4), with three forms
494
Extraocular Muscles 495
,.
M
r;--................. *
*.:.....::.;:-b:^ '.::.^ ....
'a. :. .:.
i:::;::::.*.:
o. _L. ; wF S
p.,
I: a..
_9.
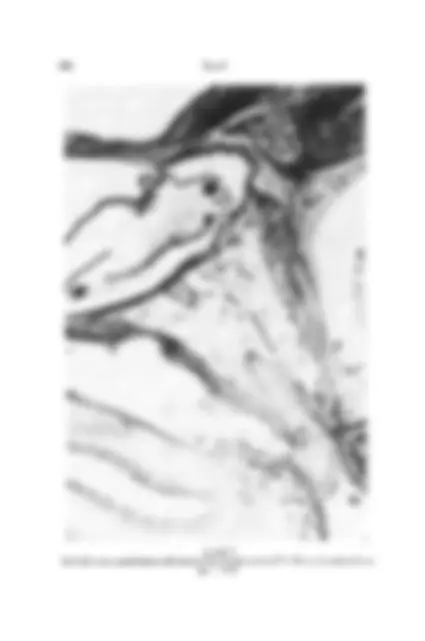
FIGURE 2 At annulus of Zinn (^) (arrow), tendon (^) (open arrow) fuses with cartilage (short arrow) at apex of orbit (^) (2/72, 88.6 (^) mm, Masson trichrome, x (^) 40).
-^ A
C.,
Extraocular Muscles
being found in any one muscle. Furthermore, in serial sections, what is
interpreted as being tapered may in deeper sections be rounded or even
split. The distance between the muscle fibers and the tendon extending from the annulus of Zinn varies at different levels of section. The ends of muscle fibers in the center of the muscle are closer to the tendon of the annulus of Zinn than the more (^) peripheral fibers. Argyrophilic fibers form "caps" at the ends (^) of muscle fibers where thickened fibers arise from the convex surface (^) (Fig 5). These (^) argyrophilic
tissue caps are continuous with the endomysium that encases each muscle
fiber. The argyrophilic cap and the loose connective tissue at the origin
are first observed in the 36-mm embryo. In certain sections, muscle caps
are derived from a thickening of the epimysium. If the distance from the muscle fiber to the tendon of the annulus of Zinn is (^) large, then the (^) argyrophilic fibers thicken and these fibers become collagenous and lose the silver (^) staining characteristics. If, however, the
distance between the muscle and the tendon is short, these fibers may
remain argyrophilic. Fine, argyrophilic fibril invaginations occur (^) between
the muscle fibers. These may be straight and slender, arrowhead, or
spiral in shape. These projections are best observed in oblique sections of
the musculotendinous junction. Occasional fibroblasts are seen within the dentate muscular extensions.
Myofibrils appear to end at the end of the muscle fibers and do not
extend beyond it.^ The myofibrils with conical ends do not converge
toward the center of the cone. The cross-striations and subsarcolemmal nuclei (^) extend as far as the end of the muscle fiber or end at a (^) varying distance from it. Internal nuclei, characteristic of embryonic muscle, are located be- tween muscle fibers. In mature extraocular muscle, occasional internal nuclei are present at the origin, particularly if the muscle fibers are split. If muscle is sectioned in the line of the long axis of the muscle, muscle
fibers appear to be^ continuous rectilinearly with the tendon. Depending
on the direction of the (^) section, the (^) angle with which the muscle meets the tendon is either acute or obtruse. The mature tendon (165 mm) is (^) composed of thick, closely packed
parallel collagen bundles with distinct longitudinal striations that in cross-
section appear as fine dots. The tendons are devoid of elastic fibers. Few fine capillaries are found in the epitenon.
General Disposition of Insertions of Extraocular Muscles
The tendinous insertions of the extraocular muscles are not identifiable before 80 mm because of their gelatinous nature. Up to 54 mm, the
497
498 Sevel
'd0y¢tiXilSSiD?2 i.f0'it- ifl't,;RSS~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~
L;iMe t' x,,(, ,~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~
X (^) X W( ;,' 0 g z; i 0, ^, ^, ;'( ( ' j~~~~~~~~~~~~~~~.........
.g, 'w0,'r(1'?i':'0: iWEqi........ FIGURE~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~.. Muslefier a ste o oigi my e apre (edca rctu, rrw) runed(speio rectus, duble arro), or^ splt^ (latera^ rectus,small^ arrw) (82 mm^ 36/72,^ M^ sson.tri ..r.m.. .....).
tendons are indistinguishable microscopically from the developing epi- sclera and sclera; only after 165 mm are they macroscopically identifiable as tendons and their sites of insertion can be measured with any accuracy (Table I). Up to 210 mm, the rectus muscle insertions are^ found^ to be equidistant from the limbus. At^ term, however,^ the insertions of the rectus muscles vary^ in^ shape and^ site^ of^ insertion.^ A^ continuous^ line joining the^ sites^ of^ insertion^ of the^ rectus^ muscles^ is^ called^ the^ spiral^ of Tillaux. 10 Histologic Features in Development ofInsertions ofExtraocular Muscles The tendons of insertion and the muscular portion of the extraocular muscles develop simultaneously. For a more lucid comprehension of the
development of the tendinous insertions, it is necessary to understand the
morphogenesis of the corneoscleral area. In 13.6- and^ 28.2-mm^ embryos,^ the corneal^ stroma^ and^ sclera merge imperceptibly and^ cannot^ be^ differentiated^ microscopically.^ From^ the 13.6- to^ the^ 22.0-mm^ stage^ of^ development,^ fibroblasts^ in^ the^ region of
insertion of the recti muscles become elongated and closely packed to-
gether. The cytoplasm is pale and basophilic. Processes that extend from the cytoplasm in a stellate fashion are less basophilic staining. The nuclei are oval and contain fine chromatin and observable nucleoli. As^ these
elongated cells mature, less^ and^ less^ cytoplasm is^ demonstrable,^ the
nuclei become flattened and^ oval,^ and^ fibrocytes^ develop.^ Collagen^ fi-
brils, which^ develop between^ the fibroblasts and^ fibrocytes,^ become
orientated in^ the^ long axis^ of the future^ tendon.
In the 26-mm specimen, at the anatomic site of the future limbus, there is a condensation of mesenchymal tissue that is orientated at right angles to the cornea and sclera. The adjacent scleral region is divided into two
distinct areas, a superficial spongy area and a deep, more compact area.
The spongy area is the location of the^ future^ episclera, while^ the more compact area is^ destined^ to^ become the^ sclera. In the 36-mm (^) embryo, sclera is distinguishable from the cornea. The
characteristic corkscrew-like^ appearance of^ the^ immature^ collagen^ of the
scleral fibers^ is^ different from^ that^ of the corneal stromal^ fibers,^ which^ are
regularly disposed and^ orientated^ in^ the^ long^ axis^ of the^ cornea.^ In^ the
38.2-mm embryo, there is a condensation of mesenchymal tissue on both
the outer and inner surfaces of the sclera. The outer mesenchymal cells
mature to become fibroblasts which in turn form the tendon of the rectus
muscle (Fig 6). The undulating tendon extends^ anteriorly and^ terminates
at a dimple where the corneal^ epithelium merges with the^ conjunctival
epithelium (Figs 7 and^ 8). The^ superficial and^ deep scleral^ condensations
noted in the 38.2-mm^ embryo are^ more^ accentuated^ in^ the^ 54-mm em-
(^500) Sevel
Extraocular Muscles
bryo. In the 78-mm embryo, the demarcation line between the corneal stroma and the sclera is more discernible and the distinct dimpling in the epithelium (vide supra) is^ less^ noticeable. The merging of^ the^ tendon^ with the^ sclera differs^ slightly^ from^ that noted at the^ origin. Muscle fibers become^ attenuated^ and^ taper toward the tendon. Distinct argyrophilic caps are not observed at the insertional
ends of the muscles. A distinct dividing line is not observed between the
muscle and the tendon, but rather a gradual merging and blending of the two tissues occur. While fine attenuated argyrophilic fibrils^ are present^ between^ the
muscle fibers, distinct^ argyrophilic connecting tissue^ between^ muscle and
aponeurosis (as observed^ at the^ origin) is^ not seen.^ Cross-striations^ may be (^) present up to the end of the muscle fiber or terminate some distance from it. Subsarcolemmal nuclei extend as far as the end of the muscle fiber,
stopping short of the muscle end, and no internal nuclei are discernible.
The collagenous fibers of the tendon intertwine, interweave (Fig 9), and interlock with the scleral fibers. Superficial fibers extend more dis-
tally than do the deeper fibers, thus^ giving a^ wider and^ more^ extensive
surface area^ of^ attachment.
Until late in development (210 mm), the tendinous insertions of the
rectus muscles have wide attachments to the sclera. These areas of at- tachment extend from the equator of the eye to the limbus (Fig 10). As the eye increases in size, there is posterior recession of the anterior
attachment of the tendon and, concurrently, differential degeneration of
the tissue between the tendon and the sclera. This degenerative process,
which commences at the equator and extends^ anteriorly, is^ characterized by vacuolation, absence of^ nuclei, and^ less^ intense^ staining with^ hematox-
ylin-eosin and^ Masson^ trichrome.
In the full-term fetus the tendons are located between the adult site of insertion and the limbus. Mature (full-term) tendons of insertion of the
rectus muscles and of the superior oblique muscle are made up of tendin-
ous fascicles, hexagonal in cross section, and consisting of closely packed
collagen fibers. These collagen fibers have been produced by fibroblastic
tendon cells that remain closely attached. The fascicles are bound to- gether by connective interfascicular tissue (endotenon), which is continu- ous with the perimysium of^ the^ muscle and^ the^ periosteum of the bone. The (^) closely packed collagen bundles are tightly bound together, lie
parallel to^ one^ another, and^ show^ distinct^ longitudinal striations.
Applied to the collagen fibers are the tendon cells (fibroblasts) that
produce them. These stellate-shaped fibroblasts occur in rows, are situ-
Extraocular Muscles
.;.:..'..: .. ...: ..: ::..:::. ...... ............ ...
.... ....
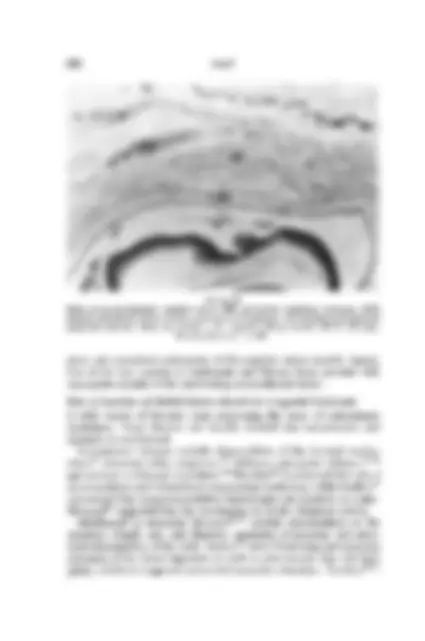
FIGURE 7 Medical rectus tendon extends to^ limbus (^) (arrow) (19/72, 26.1^ mm, (^) hematoxylin-eosin, x 15).
503
..,
if
Sevel
:.. :....: .. 9
FIGURE 8 Tendon of medial rectus muscle causing dimpling (arrow) of^ epithelium at^ limbus^ (19/72, 26.1 mm, hematoxylin-eosin, x^ 120).
504
comes semilunar while the cells of the tendon are (^) elongated and (^) closely packed. In the region of the trochlea, the mesenchymal cells become rounded (30-mm embryo) and their cytoplasmic processes are retracted. These
chondroblasts lie in amorphous intercellular substance. Chondrocytes are
noted in lacunae in the 83-mm embryo, and cell nests develop in the 210-mm fetus. By the 26-mm stage of development, the mesenchymal cells in the region of the^ tendon become^ elongated and^ closely packed. The nuclei
become ovoid and darkly basophilic staining. As the tendon traverses the
trochlea, it is rounded and the cells are so closely packed that no inter- vening cytoplasm is present. Distal to the trochlea the tendon fans out. The cells in this location remain elongated and are not closely packed, and a distinct rim of cytoplasm is present. The tendon distal to the trochlea becomes a flattened aponeurosis that fuses and interdigitates with the scleral fibers. Differential (^) degeneration occurs between the (^) developing tendon and
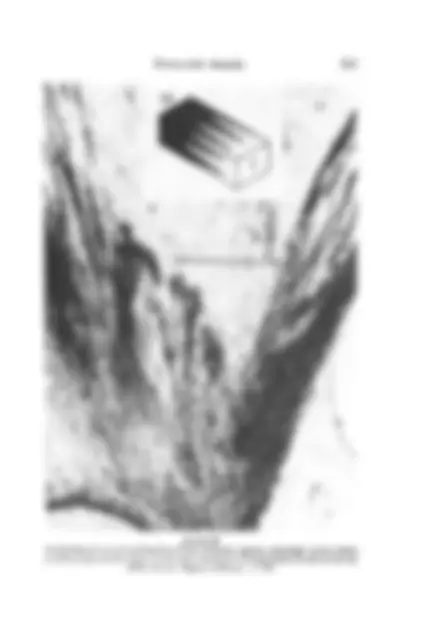
the trochlea. In the 59.5-mm embryo, the mesenchymal cells in this
region become vacuolated, their cell walls and nuclei break down, and spaces filled with lightly staining eosinophilic material are present. In the 83-mm embryo, numerous well-defined septae extend from the trochlea to the tendon (Figs 11 and^ 12). In^ the 210-mm^ fetus, these septae are less numerous and^ also^ are^ attenuated.^ Fine^ septae are located between the
tendon and^ the trochlea^ in^ the^ term^ fetus; in^ the^ adult, septae are also
present, but^ are^ fewer^ in^ number, between^ the^ capsule and the tendon
just proximal and distal to the trochlea. At (^) term, the superior oblique muscle inserts as a flattened (^) tendon into the superolateral and posterior aspect of the eye.
Insertion of Inferior Oblique Muscle
The distal end of the inferior oblique muscle is first observed in the 22-mm embryo. The insertion is muscular and fuses directly with the sclera. At the site of insertion, all the stages of muscle development (ie,
myoblast, multinucleated myoblast, myotube, and mature muscle cell)
are observed.8 No distinction can be made between muscle formation in the belly and muscle^ formation at the insertion of the inferior oblique muscle. No tendinous fibers are detected at the site (^) of insertion of the muscle. The muscle approaches the sclera tangentially. The superficial fibers extend beyond that of the deeper fibers, widening the area of insertion. The shapes of the ends of the muscle fibers are tapered, rounded, or split.
Neither argyrophilic fibers nor argyrophilic caps are present at the ends of
506 Sevel
Extraocular Muscles
the muscle^ fibers.^ Cross-striations^ extend^ as^ far^ as^ the^ end^ of the muscle fibers to the scleral insertion. At term, the inferior oblique muscle inserts via muscular fibers at the inferolateral and posterior aspect of the globe over the region of the macula.
Connective Tissue of Extraocular Muscles
Development of the connective tissue is^ inextricably^ associated^ with development of^ the^ extraocular muscles and their tendons.^ Up to 36 mm, the individual^ muscle^ cells^ are^ surrounded^ by mucopolysaccharide^ stain- ing material^ and stellate^ mesenchymal^ cells. By^54 mm,^ argyrophilic
fibrils are^ present between the^ muscle^ cells,^ to^ be^ replaced by^ fibroblasts
at about the 80-mm stage of development. Mature connective tissue is present at about 120 mm both within and surrounding the muscle. The intramuscular connective tissue fibers (internal perimysium) are uniformly distributed and consist of fine argyrophilic and elastin fibers.
There is minimal mucopolysaccharide staining material^ in^ the^ intramus-
cular connective tissue by 225 mm.^ This^ tissue^ merges with the^ capsule or
sheath (external perimysium or^ epimysium) of^ the^ extraocular^ muscles.
FIGURE 11 Mesenchymal remnants: trabeculae^ (long arrow) between^ superior oblique tendon and trochlea (short arrow) (33/73, 59.5^ mm, Masson^ trichrome,^ x^ 15).
507