Download The mechanics of hearing: The middle ear and more Lecture notes Psychology in PDF only on Docsity!
The mechanics of hearing
Jonathan Ashmore
1 Introduction
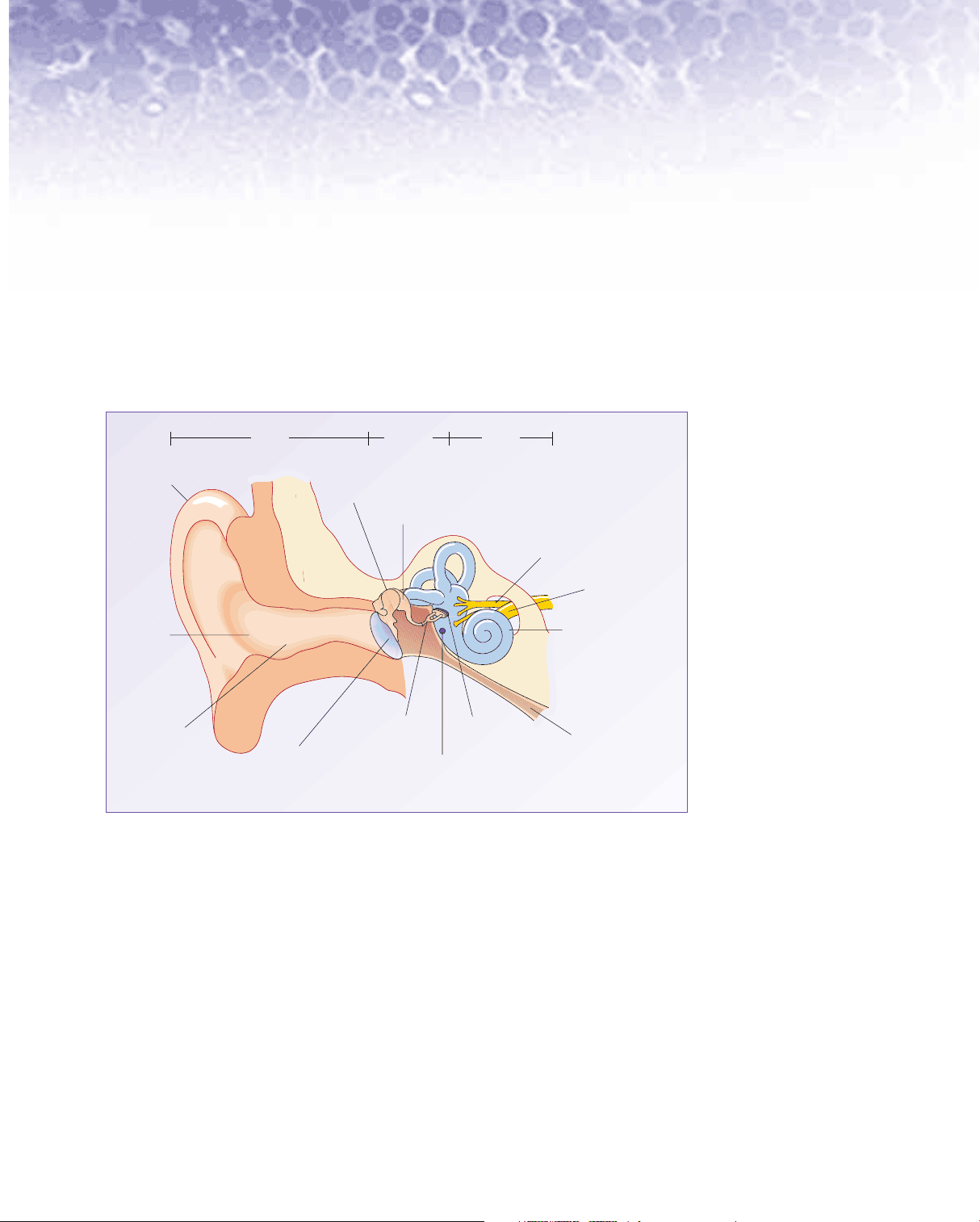
When we think of the ears, we think of the flaps that stick out from either side of our head. However these are only part of a complex physiological apparatus that enables us to hear all sorts of sounds and, more importantly, to respond appropriately. The most visible part of the ear is the outer ear , also sometimes called the external ear (Figure 1). The rest of the hearing organ, comprising the middle ear and the inner ear , is buried within the temporal bone on either side of the skull. The temporal bone can be felt just behind the outer ear. The inaccessibility of the structural components of hearing and, as we shall see, the small size of the important structures, makes the study of hearing particularly challenging.
stapes
round window
incus
malleus
oval window
cochlear nerve
Eustachian tube eardrum (tympanic membrane)
external auditory canal (meatus)
concha cochlea
pinna
middle ear
outer ear
inner ear
vestibular nerve
Figure 1 A cross section of the human ear.
We can hear because a precise sequence of events takes place in the ear. We shall explore these events in this and succeeding chapters. In this chapter, we shall trace the sequence of events from initial receipt of the signal to the stimulation of the cochlea. We use the word ‘ear’ as the collective description of all the structures and not just the outer ear. Normally when we hear a sound it is as a result of a physical disturbance transmitted through air, through the external auditory canal, through the middle ear and into the inner ear structures where it may be detected, if it is intense enough and of the right frequency. If you tap the temporal bone with your finger you can hear the percussive sound transmitted directly to the inner ear. In this case sound is said to be transmitted by bone conduction. This simple test is one of those used to identify whether hearing losses come about because sound is not being transmitted properly through the external and middle ear to the hearing organ of the inner ear or whether it is the inner ear itself that is working incorrectly.
4 SIGNALS AND PERCEPTION: THE FUNDAMENTALS OF HUMAN SENSATION
2 The sensitivity of the ear
It is usually most convenient to describe the sensitivity of the ear in terms of a threshold. This threshold is the amplitude of a sound pressure wave that can just be heard when presented to a listener. To determine such a threshold it is important that the sound is delivered in completely quiet surroundings as any background sound can interfere with the measurement. Experiments have shown that the minimum amplitude of a sound wave that can be detected under optimal conditions for a normally-hearing individual is 20 μPa (micropascals). For comparison, atmospheric pressure is approximately 100 kPa, and hence a disturbance 2 × 10 −^10 times atmospheric pressure is detectable by the ear. An equivalent comparison is to consider the situation where you blow up a normal party balloon and then start pushing the tip of a pencil into the side. When the pencil indents the side by 1 mm, the pressure inside will have increased by about 20 μPa. Our ears, like microphones, are sensitive to sound pressure. A sound stimulus can be determined by physical measurements of the sound wave at the entrance to the external auditory canal using calibrated probe microphones. Sound is measured in units of decibels (dB SPL) where SPL stands for sound pressure level , and is defined as follows using a logarithmic scale:
sound stimulus (in dB SPL) = 20 log 10 ( P / P ref )
where P ref = 20 μPa. This particular definition means that each increment of the sound wave amplitude by a factor of 10 will increment the sound level by 20 on the decibel scale. The definition is a consequence of sound energy being proportional to the square of the sound wave amplitude. The logarithmic scale is useful because of the very wide range of human hearing, where the amplitude of the sound varies by more than a million times, from the very softest that can be heard to the loudest, which can begin to produce damage. The energy per unit area (referred to as intensity) that passes into the ear from a sound wave with amplitude P is proportional to P^2 and has units of W m−^2. In air the threshold of hearing corresponds to 10−^12 W m−^2. The threshold sound stimulus depends on the frequency. Figure 2 shows the auditory threshold curve (or audiogram ) for a normal subject. The threshold rises at both lower and higher frequencies. Below about 20 Hz, the sound has to be so intense that it is possible to say that there is no hearing below this frequency. Depending on the history and age of the individual, the auditory threshold also rises at higher frequencies. The range over which we hear is, optimally, 20 Hz to 20 kHz, but can be very different from individual to individual. In particular, the upper limit of hearing is severely reduced in age-related deafness. The manner in which the measured threshold curve differs from the curve obtained from a population of normally- hearing individuals is thus a measure of auditory performance.
3 Components of the ear: overall considerations
Figure 3 shows the sequence of sound processing components in the ear. As a functional entity, these components constitute the peripheral auditory system. The function of the outer and middle ear is to transform sound from a wave travelling in air to one travelling in the fluids of the inner ear. This idea was first developed by the German scientist Hermann von Helmholtz in 1877 (Figure 4).
6 SIGNALS AND PERCEPTION: THE FUNDAMENTALS OF HUMAN SENSATION
The effectiveness of the transmission in each structure of the ear is under central nervous system control. The outer ear can be oriented towards (or away from) a sound by movement of the head, and thus the intensity of sound entering each ear can be controlled. The middle ear, responsible for transmitting sound to the inner ear, contains two muscles that control the sound passing through. The inner ear itself is the target of a neural pathway (the efferent system) that can control the sensitivity of the sensorineural cells. Together these three subsystems determine the codes sent along the auditory nerve to the first relay nuclei of the auditory pathway. The descending control from the higher central nervous system therefore acts both to limit and, if necessary, to enhance the sound stimulus reaching the sensory structures of the inner ear and shows the importance of maintaining the correct level of stimulus.
4 The outer ear
The outer ear includes the visible flap ( pinna ), the funnel-like inner portion ( concha ) and the external auditory canal ( meatus ) (Figure 1). The shape of our outer ears (and those of many mammals) is approximately that of a tapered tube with the larger end open to the outside of the head. The consequence of this shape is that sound is less effectively transmitted at frequencies below a critical frequency that depends on the length and cross-sectional areas of the meatus and concha. For the human ear this frequency is near 1–2 kHz. In addition, there is sometimes a slight sympathetic vibration in the canal itself near 5–7 kHz, when the canal behaves like an organ pipe and slightly increases acoustic transmission at these frequencies. Some animals can move their pinnae to orientate them towards a sound. In humans, with a relatively static pinna, the acoustic shielding effect of the head between the two ears and the diffraction of the sound around the head provides the major clue for the brain to work out where a sound is coming from. Clues about the elevation of a sound source are also provided by the reflection of sounds from the curved surfaces of individual pinnae, which we learn to use by processing the auditory signals.
5 The middle ear
The middle ear is a cavity interposed between the eardrum (the tympanic membrane that closes off the ear canal), and the membranous oval window that opens into the cochlea of the inner ear but retains the fluid within it (Figure 1). In construction, the middle ear on each side of the head contains the ossicular chain of three bones (the malleus , the incus , and the stapes , collectively called the ossicles ), that connect the external and inner ear structures (Figure 5). The middle ear contains (moist) air: its communication with the pharynx through the Eustachian tube allows the pressure on either side of the eardrum to be equalized to atmospheric pressure, as most airline passengers know. The middle ear acts, functionally, as a device that matches the acoustic impedance of the medium through which sound travels to the fluid in the inner ear (see Box 1, p. 9). This transformer action allows a sound wave travelling in air (in the outer ear) to become a sound wave travelling in fluid (in the inner ear) without reflection at the interface. The transformer action of the middle ear allows more than 60% of the incident sound energy reaching the eardrum to be transmitted faithfully to the inner ear structures. Without the middle ear, the efficiency of sound transmission into the inner ear would drop and the sensitivity to sound would be reduced more than thirty
ASHMORE THE MECHANICS OF HEARING 7
times. This situation occurs when the middle ear is prevented from working properly, for example, when it becomes filled with mucous fluid during an infection.
There are two main mechanisms that allow the middle ear to match acoustic impedance.
1 Area ratios
The area of the eardrum in the human ear is 17 times that of the stapes footplate, the region of the stapes in contact with the oval window of the cochlea in the inner ear. This allows the middle ear to act as a pneumatic lever: the pressure and flow velocity at the tympanic membrane is converted to a larger pressure (but a smaller flow velocity) at the cochlea:
pressure at cochlea =
area of tympanum = 17 pressure at eardrum area of stapes footplate
2 Ossicular lever
The three small bones of the middle ear, the malleus (Latin for ‘hammer’), the incus (‘anvil’) and the stapes (‘stirrup’) link the eardrum to the flexible membranous oval window of the cochlea, which contains the fluid of the inner ear. These small bones are attached together by cartilage, but essentially move as a single unit. The axis of rotation of the ossicles runs through the point where the malleus and incus are fused together. Thus the ossicles behave like a lever system:
force at cochlea =
length of malleus = 1. force at eardrum length of incus
At higher frequencies the motion is more complicated than a simple lever mechanism. More complex modes of vibration occur. The pneumatic lever and the ossicular lever schemes provide the simple basic theoretical framework for understanding how the middle ear operates. Taken together, the two mechanisms should enhance, (as an ideal transformer), the sound pressure at the cochlea ( P C) over that of the eardrum ( P T ) by a factor of 20, independent of frequency.
P C / P T = 17 × 1.2 = 20
This figure is hard to verify precisely in humans, but can be studied in animals. Figure 6 (overleaf) shows the data for the middle ear of the cat where, because of the
Figure 5 Structure of the middle ear. The diagram shows the eardrum, and the ossicular chain (stapes, malleus and incus). The axis of rotation of the ossicles is shown.
axis of rotation malleus incus stapes footplate
stapes superstructure
tympanic membrane (^) tympanic membrane piston
rigid lever system
footplate piston
axis of rotation
ASHMORE THE MECHANICS OF HEARING 9
prevent free movement of the middle ear mechanics. The functional effect is therefore to remove any impedance-matching advantage of the middle ear and to reduce the transfer of sound energy to the inner ear. Activation of the middle ear muscles occurs by a reflex pathway and constitutes the middle ear reflex. Activation certainly occurs when loud sounds are presented to the ear (i.e. when sound levels exceed about 70 dB SPL). The middle ear muscle reflex thus protects against loud sounds, although because of the time taken to contract the muscles (typically at least 40 ms), the middle ear reflex will not protect against short percussive stimuli such as gunshot noises. There is some evidence that the middle ear reflex is also activated during speaking and other vocalization. This prevents you being deafened by the sound of your own voice.
Box 1 Acoustic impedance
If we had no outer or middle ear and the oval window of the inner ear were directly exposed to sound waves transmitted through the air, only 0.1% of the sound energy would be transmitted through to the cochlea and the other 99.9% would be reflected. As a consequence, our hearing would be less sensitive by about 30 dB. The reason is that the inner ear is filled with fluid and consequently it has a much greater acoustic impedance than air. Acoustic impedance is a measure of how readily the particles of the conducting medium can be displaced by the sound waves, which is much more difficult for water than for air. The acoustic impedance is equal to the ratio of the sound pressure to the flow velocity of the particles of the transmitting medium. (It is analogous to electrical impedance, which is the ratio of voltage to current flow.) At an interface between one medium and another the amount of sound reflected, and hence the amount transmitted, is determined by the difference between the two acoustic impedances. The larger the difference, the greater the proportion reflected. The acoustic impedance of water is a factor of around 3750 times that of air, which is why 99.9% of the sound energy is reflected, and only 0.1% transmitted. This explains why, when swimming underwater in a swimming pool, you can only hear ambient sounds very faintly. The main function of the middle ear is to act as an impedance-matching device to counteract this large difference in acoustic impedances, which it does by the two mechanisms described in the text.
6 The inner ear
The inner ear is a structured fluid-filled cavity within the temporal bone. It contains the organs of hearing and of balance. In this chapter we shall consider only that portion which is concerned with hearing. This is the cochlea (Latin for ‘snail’), the organ of hearing (Figure 1). The cochlea is the site where sound is converted into a neural signal.
6.1 Overall organization of the cochlea
The cochlea is a fluid-filled tube that forms a part of the ‘bony labyrinth’. The term labyrinth emphasizes the nature of the inner ear as a series of convoluted compartments within the bone, even though we tend to think of the structure dissected out from the surrounding hard tissue. The cochlea is coiled to save space.
10 SIGNALS AND PERCEPTION: THE FUNDAMENTALS OF HUMAN SENSATION
Were the coiled turns straightened out, the tube would be about 34 mm long. In other mammals, the uncoiled length varies from about 10 mm in a mouse to about 60 mm in some whales. The cochlear tube (or duct) is closed at one end. This end is termed the apical cochlea. The other end, the basal cochlea, is linked to other compartments of the inner ear allowing fluid continuity. The basal end of the cochlea contains two flexible membranes, the oval window, on which the stapes sits, and the round window , which acts like a pressure release surface. The main structural feature of the cochlea is the basilar membrane (Figure 7). The basilar membrane is an acellular membrane mainly composed of radially-oriented collagen fibres, and provides the support membrane for the sensory cells of the inner ear. The basilar membrane divides the cochlear tube into an upper and a lower compartment, the scala vestibuli and scala tympani respectively. The two are freely connected at the cochlear apex by the helicotrema , an opening that allows the pressures in the two scalae to equalize readily. Therefore, only rapid pressure changes produced in the scala vestibuli by movement of the stapes footplate act across the basilar membrane. The cochlear tube contains a third compartment, the scala media. The scala media is a sub-compartment of the scala vestibuli and is bounded on one side by the basilar membrane. It runs the full length of the cochlear duct, and provides a special environment for the organ of Corti , the structure that contains the sensory cells of the cochlea (see below). The scala media moves with the basilar membrane so does not contribute essentially to the mechanics of the cochlea.
scala media
organ of Corti
scala tympani
cochlear nerve
scala vestibuli
cochlea
basilar membrane
spiral ligament
Figure 7 The cochlear duct showing the position of the basilar membrane and the three fluid compartments, the scala vestibuli, the scala tympani and the scala media. Uncoiled, the duct would be a tube about 34 mm long, with the basilar membrane dividing it along its length.
12 SIGNALS AND PERCEPTION: THE FUNDAMENTALS OF HUMAN SENSATION
6.4 Basilar membrane mechanics
The key element in the design of the mammalian cochlea is the basilar membrane. Through its mechanical design it responds to sounds by vibrating in a pattern that depends uniquely on the intensity and frequency of the incoming sound. The inner hair cells relay the information about this pattern to the auditory nerve and to the auditory brainstem. It is clear that more intense (i.e. louder) sounds will produce a larger pressure difference across the basilar membrane. Therefore the displacements of the hair cell stereocilia will be greater. How different tones are encoded within the whole auditory nerve is less obvious. The fundamental property of the cochlea that enables us to hear fine differences in frequency depends on the specialized mechanics of the basilar membrane. In summary the cochlea behaves like a mechanical spectrum analyser. Different frequencies excite different populations of hair cells along the length of the organ of Corti. The design of the cochlea ensures that each tone within the auditory range selectively excites only a subpopulation of hair cells along the cochlear duct. The frequencies are spread so that high frequencies excite cells at the basal end of the cochlea near the stapes, and frequencies at the low end of the auditory range excite cells at the apical end of the cochlea nearest the helicotrema. The conversion of sound frequency to coding as position of excitation is referred to as a tonotopic mapping (from the Greek, τoνoσ = sound, τoπoσ = place) (Figure 10). How do the mechanics of the basilar membrane determine the tonotopic map? The main experimental observation was made by the Hungarian-American Georg von Békésy (Figure 11) in the 1930s. He observed that the basilar membrane presents mechanical stiffness to a probe placed perpendicularly against it, the stiffness being
Figure 9 The organ of Corti. (a) A scanning electron micrograph of a section, with the view taken looking radially from the lateral side towards the spiral centre. The three rows of V-shaped stereocilia of the outer hair cells (OHC) and the single row of stereocilia of the inner hair cells (IHC) are apparent. The micrograph also shows the overlying tectorial membrane (TM) and non-sensory cells (Deiters’ cells, DC). The hair cell bodies, beneath the stereocilia, are 10 μm wide. (b) A diagrammatic representation showing the basilar membrane, the hair cells and the overlying tectorial membrane. The non-sensory cells maintain the physiology of the organ.
basilar membrane
scala vestibuli
tectorial membrane
scala tympani
scala media
TM
IHC
OHC
DC
(a) (b)
Reissner’s membrane
stria vascularis
inner hair cell
Deiters’ cells
outer hair cells
ASHMORE THE MECHANICS OF HEARING 13
100 200 (^400300)
500
(^600 )
800
900
1000
2000
3000
4000
5000
6000 8000 7000 10 000 9000
15 000
20 000
basilar membrane
greater at the middle-ear end of the cochlea than at the helicotrema end. The stiffness is graded monotonically along the duct and as a result distinct sound frequencies cause vibration of distinct sections of the basilar membrane. *
The mechanical coupling between each section of cochlea depends both on the surrounding fluid and on the cellular components of the organ of Corti. When the pressure wave enters the cochlea from the middle ear, the sound wave will propagate through the fluids of the duct. Each component of the basilar membrane will start to respond, but only those ‘tuned’ to the appropriate frequencies will vibrate maximally. The net effect is that a wave of mechanical motion (the travelling wave ) propagates along the basilar membrane and reaches a peak at the position where the frequency of the sound wave and the place frequency correspond. The estimated velocity of the cochlear travelling wave is approximately 15 m s−^1 , considerably slower than the velocity of sound in water. The wave carries the information about the frequency, intensity and temporal envelope of the sound.
6.5 The cochlear amplifier
A cochlea constructed as described would lack sensitivity and be unable to separate frequencies in an input sound. The existence of the travelling wave was deduced by Békésy from measurements made on cadaver cochleas at sound levels high enough to produce visibly detectable movements. However, scaling down to the expected displacement of the basilar membrane at the threshold of hearing produced a value that was much too small. Békésy’s data indicated that at threshold the displacement
Figure 10 The basilar membrane showing where frequencies are mapped. Only the first 2.5 turns are drawn to scale. Note that approximately equal distances are assigned to each octave (i.e. doubling of frequency) of the auditory range.
- (^) A similar conclusion, but for different reasons, was reached by Helmholtz in the mid-nineteenth century. He studied the early anatomical measurements of the cochlea made by Corti and others and thought that the basilar membrane resembled a set of piano strings, with the shorter, more tensioned treble strings at the basal end of cochlea.
Figure 11 Georg von Békésy (1899–1972). Békésy was employed by the Hungarian telephone industry before moving to the USA in 1949. His experimental work on the cochlea developed the idea of the cochlear travelling wave, for which he was awarded the 1961 Nobel Prize in Physiology or Medicine.
ASHMORE THE MECHANICS OF HEARING 15
7 Summary
Sound stimuli entering the ear are transformed by the peripheral auditory system into a series of signals sent along the fibres of the auditory nerve. The minimum detectable level of sound corresponds to an energy flow (or intensity) of 10−^12 W m−^2 in a sound pressure wave. The outer ear and middle ear structures ensure that sound energy is delivered to the inner ear where it causes a pressure wave to propagate in the fluids of the cochlea. The sound wave causes the basilar membrane stretched along the cochlear duct to vibrate and the many frequencies in a complex sound can be separated out as a result of this membrane’s mechanical properties. The vibrations at each site along the membrane deflect the stereocilia of the sensory hair cells of the organ of Corti, and lead to the signals sent to the brain. A sub-population of hair cells in the cochlea is also responsible for mechanically amplifying the movement of the basilar membrane.