Scarica Appuni su citoscheletro e più Appunti in PDF di Biologia solo su Docsity!
IL CITOSCHELETRO: è costituito da un insieme di polimeri proteici che formano strutture filamentose che sostengono e danno una forma alla cellula. Il citoscheletro è una struttura dinamica che permette alla cellula di cambiare forma. Es: in interfase il citoscheletro ha la configurazione che conferisce alla cellula la sua forma (forma che può essere diversa per ogni tipo di cellula e lontana dalla forma sferica). In fase M, invece, il citoscheletro si disassembla perché le sue componenti vengono usate per la divisone cellulare e per la formazione del fuso e la cellula assume la forma sferica, per poi ritornare alla sua forma caratteristica alla fine della divisione. Il citoscheletro è costituito da:
- Microfilamenti : polimeri di actina, si trovano a ridosso della membrana plasmatica
- Microtubuli : polimeri di tubulina, si irradiano dal centro della cellula verso la periferia
- Filamenti intermedi : polimeri di diverse proteine, hanno una distribuzione reticolare e sono più densi attorno al nucleo. NB: microfilamenti e microtubuli costituiscono dei binari sui quali avviene il trasporto di vescicole e organuli grazie a proteine motrici dette motori molecolari.
MICROFILAMENTI: sono polimeri di actina.
NB: L’actina polimerica è una proteina globulare (G- actina) e presenta un sito di legame e idrolisi di ATP. Può quindi trovarsi in vari modi:
- Senza nucleotide legato
- Con ATP o ADP legato-> nella cellula prevalgono queste forme. I monomeri di actina si possono associare per formare polimeri costituiti da 2 filamenti avvolti ad alfa elica (F-actina).
Caratteristiche fondamentali dei microfilamenti di F-actina:
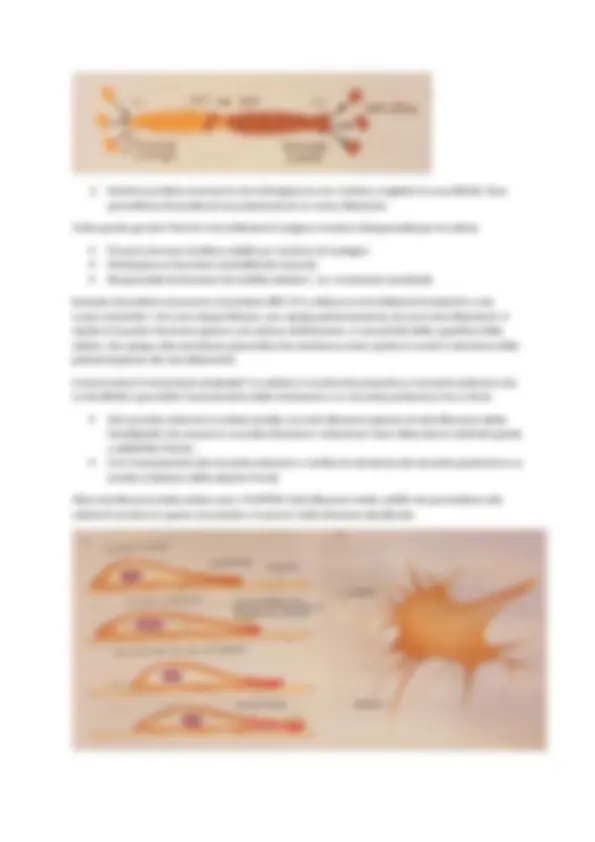
1. Polarità intrinseca: le estremità dei filamenti differiscono l’una dall’altra. Perché? Se osserviamo al microscopio una preparazione ottenuta con actina filamentosa e miosina II (ha elevata affinità per l’actina) notiamo che il filamento di actina viene ricoperto dalle teste della miosina II che hanno tutte lo stesso orientamento> la loro disposizione dà luogo a un aspetto a punta di freccia: Da una parte abbiamo le punte delle frecce- ESTREMITA' A PUNTA. Da una parte abbiamo i barbigli (prolungamento laterale della freccia) - ESTREMITA’ A BARBIGLI La polarità è utile per le proteine motrici che si muovono sui filamenti in modo direzionale (alcune verso una estremità, altre verso l’altra, ma in maniera specifica) e costituiscono sistemi di trasporto diretti dal centro alla periferia e viceversa. 2. Dinamiche di polimerizzazione e depolimerizzazione: L'associazione di due monomeri a formare un dimero ha una probabilità molto bassa, quindi, affinché possa avvenire la polimerizzazione è necessario un trimero> esso è più stabile e comporta interazioni: testa-coda con altri monomeri laterali a opera del monomero centrale (forma la struttura a doppio filamento)
- Esistono proteine accessorie che interagiscono con l’actina e regolano la sua attività. Esse permettono di accelerare la produzione di un nuovo filamento. Tutto questo perché? Perché i microfilamenti svolgono funzioni indispensabili per la cellula: Possono formare strutture stabili con funzione di sostegno Partecipare ai fenomeni contrattili del muscolo Responsabili di fenomeni di motilità cellulare > es: movimento ameboide Esempio di proteina accessoria: la proteina ARP 2/3 si attacca ai microfilamenti esistenti e crea nuove estremità + che sono disponibili per una rapida polimerizzazione di nuovi microfilamenti> il ripetersi d questo fenomeno genera una estesa ramificazione, in prossimità della superficie della cellula, che spinge sulla membrana plasmatica (la membrana viene spinta in avanti in direzione della polimerizzazione dei microfilamenti). Come avviene il movimento ameboide? La cellula in movimento presenta un versante anteriore che si estroflette e permette l’avanzamento della membrana e un versante posteriore che si ritrae. Dal versante anteriore la cellula emette una estroflessione planare di estroflessione detta lamellipodio che avanza in una data direzione> estensione viene attaccata al substrato grazie a ADESIONI FOCALI. Con l’avanzamento del versante anteriore si verifica la retrazione del versante posteriore e si assiste al distacco delle adesioni focali. Altre estroflessioni della cellula sono i FILOPODI (estroflessioni molto sottili) che permettono alla cellula di sondare lo spazio circostante e muoversi nella direzione desiderata.
3. MICROTUBULI: Sono formati da polimeri di TUBULINA> nella cellula si presenta in forma eterodimerica, esiste: La tubulina alfa La tubulina beta I dimeri si associano tra loro attraverso interazioni della subunità beta di un dimero con la subunità alfa di un altro. I dimeri interagiscono lateralmente e formano più file rettilinee di dimeri dette protofilamenti> formano una struttura con cavità centrale, ossia il MICROTUBULO (nella cellula i microtubuli sono costituiti da 13 protofilamenti). Per i microtubuli possiamo fare considerazioni analoghe ai microfilamenti:
- I polimeri presentano una polarità strutturale: una estremità è caratterizzata da tubulina alfa (estremità -), l’altra estremità è caratterizzata da tubulina beta (stremità +, l’aggiunta di dimeri è più veloce qui.
- Il nucleotide legato alla tubulina alfa non si dissocia e non è scambiabile, mentre il nucleotide legato alla tubulina beta può essere idrolizzato e scambiato (prevale la forma con GTP legato).
Perché? Nella cellula l’estremità - non è mai libera, si trova legata ad un complesso ad anello (costituito da tubulina Y) che chiude e stabilizza l’estremità -. Questo complesso funge da centro di nucleazione per l’inizio della polimerizzazione e si trova in corrispondenza di addensamento proteico che circonda i centrioli e prende il nome di MATERIALE PERICENTRIOLARE. Il materiale pericentriolare + i centrioli = CENTROSOMA chiamato anche centro di organizzazione dei microtubuli. Il centrosoma fa sì che tutti i microtubuli si dispongano in un certo modo. Essi si dipartono dal centro della cellula, verso cui è rivolta l’estremità - e si irradiano verso la periferia, verso cui è rivolta l’estremità +. L’instabilità dinamica fa sì che: l’estremità + possa andare alla continua ricerca dei bordi della cellula fa sì che il centrosoma si trovi sempre al centro della cellula (quando i microtubuli su entrambi i lati raggiungono il cortex si produce un uguale forza che blocca il movimento del centrosoma) Al microtubulo si associano delle proteine motrici necessarie per il trasporto che deve avvenire dal centro alla periferia (chinesine si spostano verso estremità +) o dalla periferia verso il centro (dineina si sposta verso estremità -). IL CENTRIOLO: è un organulo costituito da microtubuli disposti in 9 triplette. Nelle triplette il microtubulo più vicino al centro è costituito da 13 protofilamenti, gli altri due microtubuli da 10. Nelle cellule animali si trovano due centrioli (DIPLOSOMA) disposti in modo perpendicolare:
- Centriolo parentale: presenta delle appendici che si estendono a una estremità
- Centriolo figlio privo di appendici
I centrioli si duplicano in maniera coordinata al ciclo cellulare: con la duplicazione accanto al centriolo originale si sviluppa un nuovo centriolo che si dispone perpendicolarmente e che accresce fino a raggiungere le dimensioni del centriolo maturo> il centriolo figlio all’inizio è privo di appendici, si sviluppa durante la mitosi e diventa il centriolo parentale della nuova coppia. LE CIGLIA: Sono estroflessioni di membrana deputate al movimento cellulare in organismi unicellulari e al movimento di materiale esterno alla cellula. Il loro movimento è utile per la pulizia delle superfici e richiede ATP. FLAGELLI: sono strutture di maggiori dimensioni rispetto alle ciglia, si trovano in cellule dotate di mobilità propria (es: spermatozoi). NB: Nei flagelli e nelle ciglia si osserva una struttura (prende il nome di assonema) composta principalmente da microtubuli. All'interno del citoplasma della cellula, nella porzione basale del ciglio, si osserva il corpo basale, che si allunga all’interno della cellula con l’apparato delle radichette, che hanno la funzione di ancorare saldamente tutta la struttura dell’assonema al citoplasma della cellula. Il corpo basale è costituito da nove triplette di microtubuli assimilabili alla struttura del centriolo. Il resto del corpo dell’assonema è costituito da una disposizione dei microtubuli detta 9+2: troviamo una coppia di microtubuli al centro (costituiti da 13 protofilamenti) circondata da 9 doppiette (microtubulo A 13 protofilamenti e microtubulo B 10 protofilamenti).
VI CLASSE: nestina METABOLISMO ENERGETICO DELLA CELLULA All’interno della cellula si verificano continuamente reazioni chimiche necessarie per sintetizzare composti organici specifici e per trasformare in energia chimica utilizzabile al proprio interno l’energia prelevata dall’ambiente esterno sotto diverse forme. Tutte le reazioni che consentono di produrre energia dalla trasformazione dei nutrienti in molecole più semplici e di trasferirla, utilizzarla e accumularla nelle cellule costituiscono il metabolismo energetico. Carboidrati, lipidi e proteine sono utilizzati dalle cellule come fonte di energia attraverso la loro graduale ossidazione. Tuttavia, la loro energia non può essere utilizzata direttamente , ma deve essere prima trasferita in un composto, l’adenosina trifosfato (ATP) , che viene utilizzato per fornire energia alla maggior parte dei processi che ne richiedono. L'ATP è formato da una base azotata, l’adenina, dal ribosio, che è uno zucchero pentoso e da tre gruppi fosfato. Nel citoplasma il pH è neutro e gli ossidrili dei gruppi fosfato che compongono l’ATP sono completamente ionizzati, pertanto l’ATP ha 4 cariche negative concentrate intorno alla struttura lineare di polifosfati. La repulsione tra le cariche negative spiega la tendenza del gruppo fosfato terminale a staccarsi per idrolisi, dando luogo a una molecola di ADP e a una di fosfato inorganico in una reazione esoergonica. Una reazione biochimica , che avviene all’interno della cellula, può svolgersi solo nella direzione che porta a una diminuzione dell’energia libera del sistema. Cosa si intende per energia libera? Per energia libera (G) si intende la quantità di energia disponibile per svolgere un lavoro, cioè l’energia che può essere utilizzata. NB: più lontano dall’equilibrio è un sistema, maggiore è la sua energia libera; a mano a mano che la reazione procede, l’energia libera diminuisce e il sistema si avvicina all’equilibrio.
Quando un sistema raggiunge uno stato che possiede una energia libera minima allora si parla di stato stazionario. Una reazione avviene spontaneamente solo se essa porta il sistema più vicino al suo equilibrio, cioè se è accompagnata da una variazione di energia libera (deltaG) negativa, dove per deltaG si intende la differenza tra energia libera dei prodotti della reazione (stato finale) e l’energia libera dei reagenti (stato iniziale). Tuttavia, se una reazione chimica può svolgersi, non è detto che questa avvenga autonomamente: alcune volte la reazione non avviene a temperatura ambiente, ma è necessario che i reagenti vengano surriscaldati> il calore applicato rappresenta l’energia di attivazione. Nelle cellule l’energia di attivazione è ridotta grazie alla presenza di catalizzatori proteici, gli enzimi. Essi non influiscono sulla direzione, ma unicamente sulla velocità di reazione (perché la reazione si volge sempre nella direzione che porta a una diminuzione dell’energia libera del sistema). Questa condizione è soddisfatta dalla maggior parte delle reazioni cataboliche, a deltaG negativo, dove molecole complesse vengono degradate a prodotti più semplici e dotati di una minore quantità energia libera (reazioni esoergoniche). La cellula deve comunque compiere un gran numero di reazioni di sintesi (reazioni anaboliche), elaborando molecole e macromolecole complesse, dotate di un alto grado di energia libera. Queste reazioni, dette endoergoniche, hanno un deltaG positivo e non potrebbero teoricamente avvenire. Al fine di poter rendere possibili reazioni con deltaG positivo, nelle cellule tali reazioni sono accoppiate a una reazione esoergonica in cui deltaG sia maggiore, in valore assoluto, del deltaG della reazione endoergonica, in modo che il deltaG complessivo delle due reazioni sia comunque <0. MITOCONDRIO I mitocondri sono gli organuli responsabili della produzione di energia, variano molto nella forma, da lunghi e sottili a corti e larghi. La morfologia e le dimensioni variano secondo il tipo di cellula, ma anche all’interno di una stessa cellula si comportano come strutture dinamiche (hanno la capacità di cambiare forma). Il loro numero varia secondo il tipo di cellula e tendono ad accumularsi in corrispondenza delle zone nelle quali c’è una maggior richiesta di energia. I mitocondri sono presenti in tutte le cellule eucariotiche, in particolare sono abbondanti in quelle in cui ha luogo: Motilità (cellule ciliate e coda dello spermatozoo) Contrazione (cellule muscolari striate scheletriche e cardiache) Intenso lavoro metabolico (epatociti) Attività sinaptica (cellule nervose) Produzione di calore (adipociti). NB: Quando il consumo energetico delle cellule muscolari è elevato, essi sono in grado di duplicarsi rispondendo ad un aumento delle richieste energetiche. L'aumento dei mitocondri determina un aumento della loro capacità di produrre energia.