Scarica Computational Cardiology e più Appunti in PDF di Ingegneria Biomedica solo su Docsity!
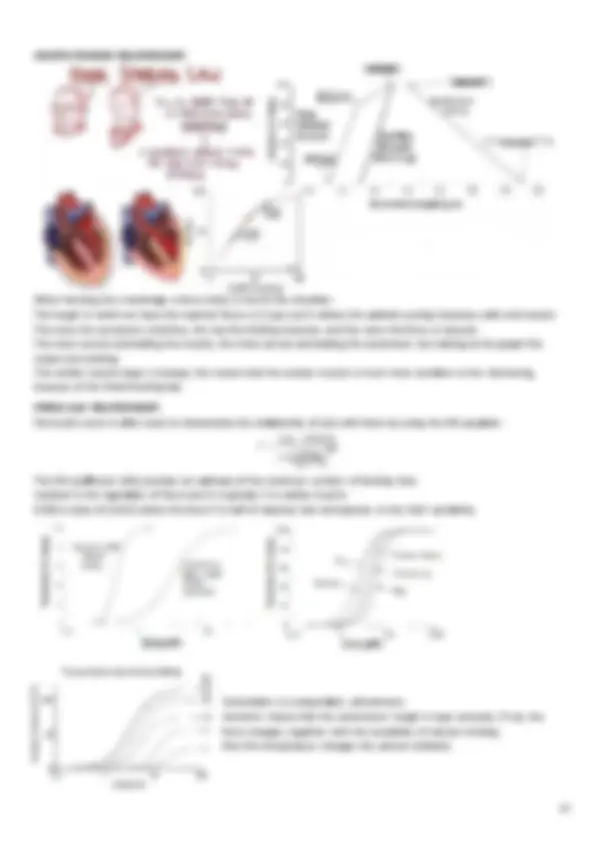
The heart is a pressing and aspiring pump: contraction and relaxation are active processes (with energy consumption), formed by several phases, each one lasting from few to many tens of milliseconds. The heart is a muscle, with a pumping capacity of about 7 L/min. It contracts thanks to electrical coordination of the muscle cells, meaning that its electrical function has a cellular basis: the action potential (AP), that is generated in the sinoatrial node (with the pacemaker cells, the cardiomyocytes) and then propagates through a conducting system, for fast activation. The heart contains 3 main types of muscle fibers:
- fibers of the specific system of excitation (nodal tissue), characterized by self-excitation (they produce spontaneously the AP)
- fibers of the specific system of conduction, characterized by high velocity of conduction, enabling a fast propagation of the AP, to allow fast and sequential activations of the portions of the heart
- fibers of the contractile myocardium (atria and ventricles), activated by the AP that causes their contraction, enabling the mechanical pumping function of the heart
HEART PHISIOLOGY
COMPUTATIONAL CARDIOLOGY
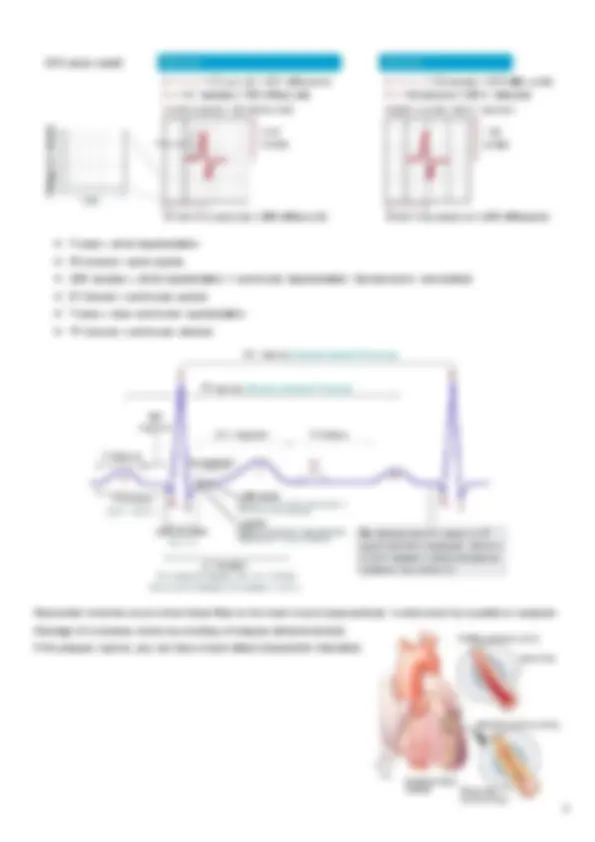
All PM structures are capable of spontaneous depolarization (automaticity) and can, therefore, serve as the heart’s PM, but the rate of the SA node is higher, so it imposes its rhythm on the others, being the primary one. An ECG lead displays an ECG curve (diagram) and at least 2 electrodes are necessary to obtain an ECG lead: one serves as a reference and the other serves as an exploring electrode. The electrocardiograph compares the electrical potentials detected in the electrodes: if a vector travels towards the exploring electrode and away from the reference electrode, then a positive wave is printed and viceversa.
Arrhythmia (ectopic beats) means any situation that cannot be classified as a normal heart rhythm, intended as a rhythm originating in the sinus node, regular (when the variation between consecutive cycles is <0.16 s) and with normal frequency (between 50 e 100 bpm) and conduction (when the P wave lasts <120 ms, PR interval lasts ≤200 ms, QRS complex lasts ≤100 ms).
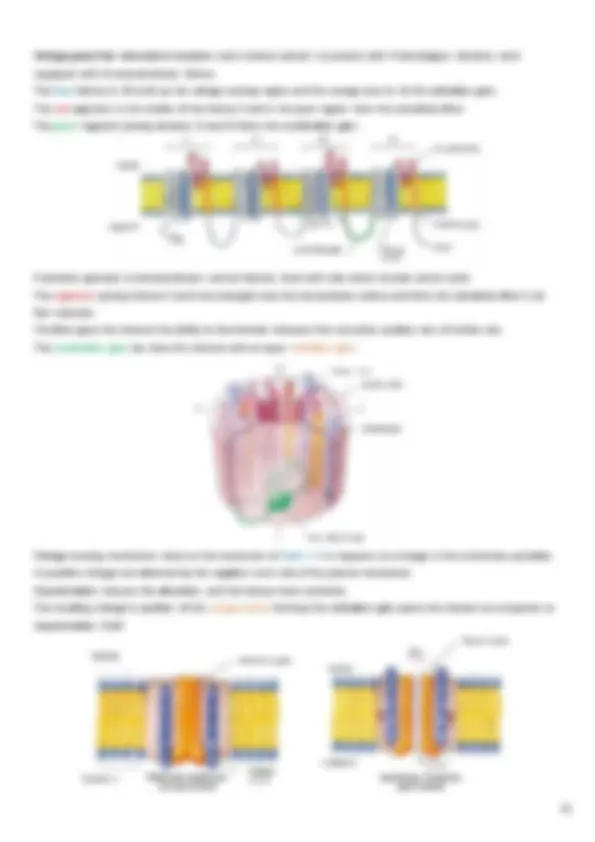
Each cardiomyocyte contains a single, centrally located nucleus surrounded by a cell membrane known as the sarcolemma , that contains voltage-gated Ca2+ channels. Excitation-contraction (EC) coupling in striated (skeletal and cardiac) muscle physiology defines the transduction of an electrical stimulus (AP) to a mechanical response (contraction) and it involves the following events:
1. depolarization of the plasma membrane and its invaginations (the t-tubular system) by an AP 2. transduction of the depolarization signal to the sarcoplasmic reticulum (SR) membrane 3. activation of Ca2+^ release from the SR and subsequent global elevation of intracellular [Ca2+] 4. transient interaction of Ca2+^ with contractile proteins leading to muscle contraction 5. return of [Ca2+] back to levels at resting conditions and muscle relaxation
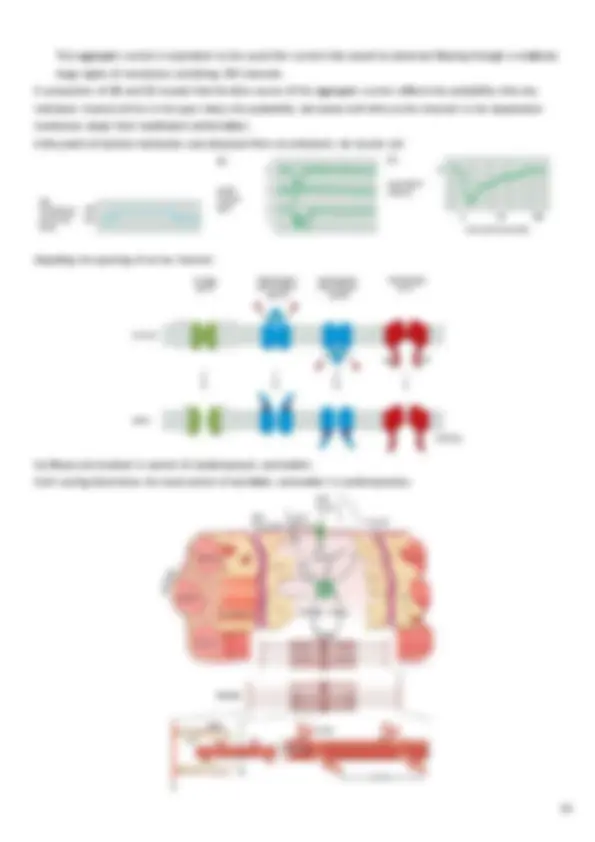
Time evolution of a reaction:
- A(t), B(t) and C(t) state variables
- initial conditions: A(t=0) = A 0 B(t=0) = B 0 C(t=0) = C 0 = 0
- assuming there are no other reactions which involve A, B and C other than A(t) = A 0 – C(t) and B(t) = B 0 – C(t) only 1 state variable remains: C Substituting A(t) and B(t): we obtain a quadratic differential equation whose solution represents, at each time t, the amount of complex C(t) present in the system. The equation is non-linear and, in general, there is only a numerical solution. In this case, however, there is also an analytical solution. The numerical solution can be obtained, e.g, by implementing a model of the differential equation in Simulink: Considering A 0 > B 0 and C(0) = 0 the analytic solution becomes: with The reaction stops when C=B 0 given that B is the reagent present in a lower amount. This is not a mono-exponential growth, even when A 0 >> B 0 it can be approximated by a first order kinetics:
doesn’t represent the time constant, because it depends on the initial conditions (A 0 ,B 0 ). It is not possible to derive the transfer function of the system, but it is possible to study the equilibrium points:
There are 2 equilibrium points: C = B 0 and C = A 0
but it is not possible that they are both stable, because there would be indeterminacy. By linearizing the system equation, it is possible to evaluate the sign of the eigenvalue λ in the 2 equilibrium points: Having hypothesized A 0 > B 0 , the stable equilibrium point is C = B 0 , which corresponds to a negative eigenvalue. Also, note that the condition C = A 0 would lead to a negative B value = B 0 – C, so it is not physically reachable.
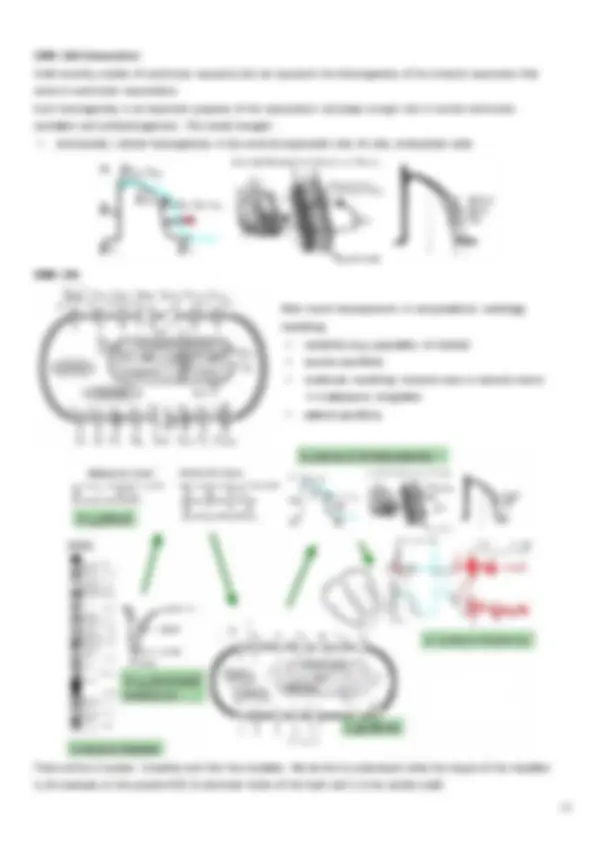
In an enzymatic reaction, an enzyme (E) binds to a substrate (S) to form a complex (C), which leads to the formation of a product (P) and to the release of the enzyme: E catalyses the transformation of S into P and, at the end of the reaction, it is released unchanged, ready to bind to a new substrate molecule. E is not "consumed" in the reaction, and it is present in a much lower amount (e.g. 10-6) than that of S (E<<S). Michaelis-Menten Model (1913): With: • k 1 → rate of C formation
- k- 1 → rate of C dissociation
- k 2 → rate of P formation The link between E and S is reversible, i.e. not all C successfully leads to the formation of P: sometimes, E and S are again obtained. The formation of P, on the other hand, is not reversible, therefore it is no longer possible to obtain S from E and P. Given appropriate initial conditions, i.e. let’s assume to start with E 0 << S 0 and no C and P:
- E 0 = E(t=0) → E initial amount
- S 0 = S(t=0) → S initial amount
- C 0 = 0
- P 0 = 0 → it is possible to write 2 balance equations for E and S, by assuming that they are not affected by other reactions: Since there are 4 unknowns (E, S, C e P), 2 other equations are needed, which are obtained by applying MAL. 1. kinetics of C formation: 2. kinetics of E: 3. kinetics of P formation: 4. kinetics of S: Of the 4 available differential equations (which are linearly dependent on each other), only 2 are needed:
- let’s keep n. 3, which is the output equation (in which P appears)
- among the other 3 we choose n. 1 (it would be equivalent to choose the n. 2 or n. 4)
ENZYMATIC KINETICS
The following equation system is then obtained: From the analytical point of view it is not possible to find a solution, at least without appropriate simplification hypotheses. We can solve the system numerically, for example by implementing the equations in Simulink. We can analytically study the condition of the system in steady state, with an equilibrium analysis: A single equilibrium point is obtained that corresponds to the complete transformation of S into P: the quantity of enzyme is equal to the initial one and there is no complex. The equilibrium solution does not depend on the k of the reactions, which only affects its speed. To have an analytical solution, given the complexity of the problem, it is necessary to introduce simplifying hypotheses : they’re quasi-steady-state hypotheses, since the faster dynamics (which are always considered in steady- state) are neglected and only the slower ones are analysed. For example, we can consider that after the initial phase of the reaction the C complex is in steady state:
- whenever a complex will give rise to a product, there will be a substrate molecule that will bind to the enzyme forming a new complex
- C will therefore remain constant over time Simplification: 𝑑𝐶 𝑑𝑡
C in steady-state → we can study the trend of C as a function of S: When S ↑↑, the enzyme is almost all bound into the complex. When S ↓↓, only a small part of the enzyme is bound.
C in equilibrium → study of P(t): When S ↑↑, the speed of formation of the product is maximum: P grows linearly. When S ↓↓, the speed of formation of the product reduces up to 0: all the substrate was transformed and P=S 0. Progression curve of a simple reaction catalysed by an enzyme: The kinetic ratio of an enzyme as a function of the concentration of available substrate can be obtained with a simple experiment:
- prepare a series of sample tubes containing increasing concentrations of substrate
- at T0, a fixed quantity of enzyme is added
- in the subsequent minutes, the concentration of formed product is measured (e.g., absorbance measure) and the velocity is derived
- when the substrate is much more than the enzyme, the observed ratio is the maximal velocity of the enzymatic reaction
C in equilibrium→ study of C(S): When S ↑↑, the enzyme is almost all involved in the complex. When S ↓↓, only a small part of the enzyme is tied to the substrate. COMPETITIVE INHIBITION It occurs when in the reaction an inhibitor (I), which is capable of binding to the active site of the enzyme (E), is added, preventing the binding of the enzyme with the substrate (S) to form the complex C1 that will produce the product (P). Another complex (C2) is formed, that will not lead to any product. I and S compete to finalize the binding with the active site of E. 6 variables → 6 equations are needed Balance equations of E, S e I: The other 3 equations are obtained by applying the MAL.
1. C1 formation kinetics: 2. C2 formation kinetics: 3. P formation kinetics: Also here, we introduce the hypothesis of quasi steady state:
The system is non-linear: therefore, let’s introduce the hypothesis of quasi-steady state: For simplicity, let’s take k1=k1’ and k3=k3’, considering the effect of the inhibitor implicit in its reaction with E to produce C2. Under these hypotheses, it is possible to compute C1 as a function of S: with and In the allosteric inhibition also when S ↑↑ it is evident the effect of the inhibitor, that reduces the asymptotic value of C1: Given that , the final effect of the inhibitor is to reduce the velocity of formation of the product P, independently from the quantity of S. Small amount of inhibitor is enough to drastically change the velocity: I << E << S COOPERATIVITY It occurs when an enzyme (E) has more than 1 binding site for the substrate (S) and, when 1 of these sites is occupied, the affinity of the others increases. Considering 2 binding sites for the substrate, the product can be obtained from 2 different reactions: The system is non-linear: therefore, let’s introduce the hypothesis of quasi-steady state: The double binding site leads to a quadratic relationship for C2 and therefore for dP/dt. The equation represents the velocity of formation of the product P as a function of the quantity of substrate S, in case of 2 completely independent reactions and affinity constants.
Zero cooperativity: let’s suppose that all bonds between S and E are equiprobable, i.e., when only 1 of the 2 binding sites is occupied with the substrate, the affinity of the other does not change. We seek for a relationship between the reaction constants during the binding between S-E: When E has 2 free sites, it will have a x2 probability to bind S. When E has 2 free sites, it will have a x2 probability to lose S or to generate P. The maximum velocity of formation of P is exactly the double with respect to the one with only 1 binding site. This velocity corresponds to the one that occurs when a double quantity of enzyme E 0 is used, with only 1 active site. Considering an enzyme with n binding sites, the formation velocity of the product would be multiplied by a factor n. Maximum cooperativity: when the substrate can bind the enzyme ‘almost only if’ the other site is already occupied. The structure of the equation remains unchanged, but we have S2 instead of S. Therefore, we have a quadratic increase in the origin instead of a linear one. The velocity of formation of P by varying S can be represented by a sigmoid: 3 different regions: the velocity is almost not affected by variations of S → the velocity varies almost linearly with S (depending on the enzyme’s amount) Generalization: n binding sites By varying n (integer), the slope of the sigmoid changes, but the inflection point S=km remains unchanged.
Equilibrium points: → Jacobian matrix of J(x) evaluated in xeq: Using the Taylor expansion of the 1 ° order: For an hyperbolic equilibrium, ALS trajectories are topologically equivalent (homeomorphic) to the ones of the NLS. This is true for nodes, saddles and focuses, while it is not possible to determine what is a center of the ALS in the NLS. Many Jacobian matrices exist, 1 for each equilibrium point of the system. Each matrix has its n eigenvalues 𝜆𝑖 𝑤𝑖𝑡ℎ 𝑖 = 1 … 𝑛 of which: The equilibrium point is stable only if 𝑛+ = 0 , otherwise it is unstable. It is possible to build the ALS relative to the NLS in correspondence of one of its equilibrium points only if 𝑛 0 = 0 , i.e., if the state matrix is non-singular. Example: 1° order nonlinear dynamical system Equilibrium points: Jacobian matrices (1x1): For each Jacobian, there is only 1 eigenvalue 𝜆, which is μ for the first equilibrium point and - μ for the second one. Associated linear systems: Study of the equilibrium: by varying the sign of eigenvalues changes, and, consequently, also the stability of the equilibrium points changes. Hartman-Grobman theorem: the trajectories around the equilibrium point of the ALS (origin) and those around the equilibrium point of the NLS are homeomorphic: so, it always exists a bijective (one-to-one) correspondence that allows to transform one trajectory into another. In particular, the topology is preserved: if in the ALS there is a stable focus, then also in the NLS there is a stable focus. The homeomorphism maintains the trajectory direction (but this does not mean that the trajectories are the same). Consequently, 2 homeomorphic trajectories are topologically equivalent. = associated linear system
BIFURCATION THEORY
Let’s consider a dynamical system with state x, described by an ODE dependent on a parameter p: by varying p, the state space is maintained but the evolution law changes: 2 different situations: the trajectories of S and S’ are homeomorphic or a topological change of the system occurs: a change in the number of equilibrium points and/or in their nature. The system crosses/undergoes a bifurcation, defined as the loss of the topological equilibrium that occurs in correspondence of a specific critical value of the parameter pbif and, after this point, the system loses or gains a structural property. The control plane shows how the equilibrium changes by varying a parameter: in the origin, the 2 equilibria collide, so there is a bifurcation and the stability is inverted. GRADIENT SYSTEMS For dynamical conservative systems., it can be defined a potential (U), that maps each state x to a real value. One advantage of the gradient systems is that equilibrium points can be found by finding the maximum and minimum points of U:
- potential maximum: unstable equilibrium 0
- potential minimum: stable equilibrium 0 Let’s consider the simplest gradient system that has bifurcation with change in stability (transcritical) : Equilibrium points: Eigenvalues: The stability depends on the sign of p. When x→ 0 , U(x) acts as a parabola opening up or down depending on the value of p: When x→ U(x) acts as a cubic function, tending to the infinity: U(x) has a double zero in the origin and a zero in the point 𝑥 = 3 2
Extrema are in x=0 and x=p and their role depends on the sign of p. (when n=1)