Margherita Re ISTOLOGIA I A.A 2025/2026
TESSUTO EPITELIALE
→ tessuto costituito da cellule strettamente adese tra loro, con giunzioni specializzate
→ il tessuto epiteliale ha funzione di rivestimento e protezione
→ la matrice extracellulare è pochissima tra le cellule: è strategico perché ha funzione di isolare da patogeni
. TIPI DI TESSUTO EPITELIALE:
• rivestimento: riveste le superfici esterne del corpo e le superfici interne degli organi cavi (organi e vasi)
e le superfici delle tonache sierose (pleure, pericardi)
• ghiandolare
• sensoriale
• particolarmente differenziati (ad esempio lo smalto dei denti)
→ GLI EPITELI DERIVANO DA TUTTI E 3 I FOGLIETTI EMBRIONALI
. FUNZIONI DEGLI EPITELI DI RIVESTIMENTO:
• secrezione (epitelio stomaco): produce e secerne i succhi gastrici
• assorbimento (intestino): assorbe sostanze nutritizie
• trasporto: favoriscono trasporto materiali o lungo o attraverso l’epitelio
• protezione (epidermide e transizione della vescica)
• funzione recettoriale: riceve e trasmette stimoli esterni
→ CARATTERISTICHE GENERALI:
• non sono vascolarizzati ma sono INNERVATI e contengono cellule sensoriali che sono dotate di polarità
morfologica
→ non è vascolarizzata perché evita al patogeno di entrare nel circolo sanguigno
→ innervata e con recettori sensoriali perché permette la sensibilità
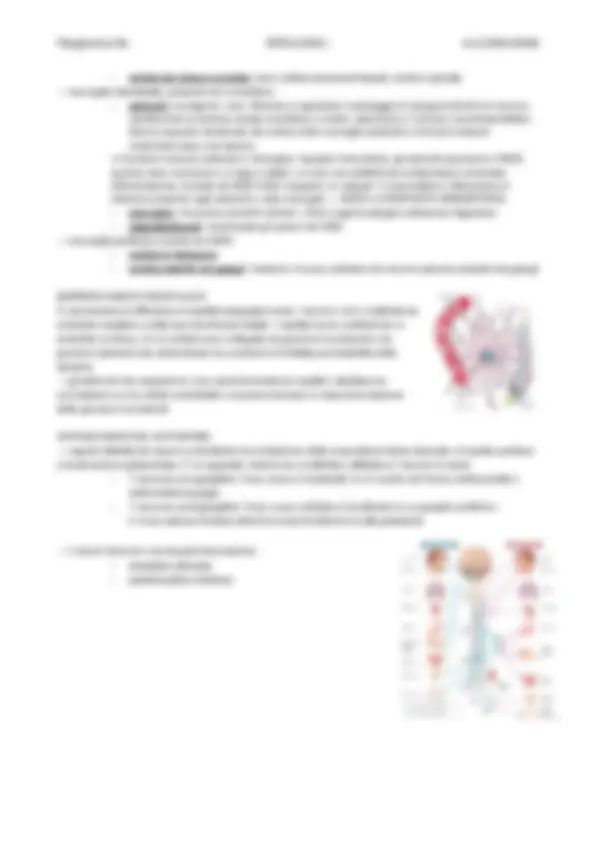
• è dotata di polarità cellulare: 3 domini (basale, laterale, apicale) che danno caratteristiche diverse
perché hanno composizioni morfologiche diverse
→ Il dominio basale ha delle giunzioni che lo legano alla lamina basale, una struttura che sta sotto le cellule
epiteliali che si legano ad essa, conferendo stabilità e protezione del tessuto connettivo (molto
vascolarizzato perché nutre le cellule, per diffusione, dell’epitelio di rivestimento) inoltre essa fa da filtro
per le molecole che passano in entrata e/o in uscita
→ 3 DOMINI CELLULARI:
. LATERALE: è caratterizzato dalla presenza di 3 tipi di giunzioni: - occludenti
- ancoraggio
- comunicanti
⇉ occludenti: sono nella parte più vicino all’epitelio in modo da chiudere lo spazio intercelluare
2 tipi di trasporto: - paracellulare (passaggio di piccole molecole)
- transcellulare (usa sistemi trasporto selettivi)
le proteine delle giunzioni occludenti sono:
tetraspannine (4 domini transmembrana e porzioni N- e
C- terminali intracellulari)
⇉ le principali son 3: - occludina
- claudina
- di adesione cell.
⇉ ancoraggio: assicurano stabilità meccanica alle cellule
epiteliali legando il citoscheletro di una cellula a quello
della cellula adiacente: unità strutturale dell’epitelio
→ giunzioni aderenti: sono funzioni transmembranali con
la E-caderina collegata a microfilamenti di actina ⇉ sono
giunzioni calcio dipendenti
! sono molto numerose e creano dispositivi giunzionali
⇉ LE GIUNZIONI SI GENERANO DALLA PROTEINA NECTINA CHE SI LEGA AI MICROFILAMENTI DI ACTINA
→ mediano l’inibizione da contatto
→ processo di transizione epitelio-mesenchimale (trasformazione di cellule epiteliali in cellule mesenchimali
cioè cellule molto immature che porta alla perdita di E-caderina e quindi alla trasformazione tumorale)