Scarica MALATTIE MONOGENICHE e più Appunti in PDF di Genetica Umana E Delle Popolazioni solo su Docsity!
TRASMISSIONE DEI CARATTERI MONOGENICI
Riprendendo le domande iniziali del corso, abbiamo fatto un buon approfondimento su quella che è la genetica molecolare, rispondendo alla domanda “qual è la causa della malattia? Perché queste persone si ammalano?”. Abbiamo fatto anche alcuni esempi e ne faremo molti altri. Altra domanda che ci siamo posti all’inizio del corso è “com’è l’ereditarietà di questi caratteri monogenici? Di fronte a una persona che ha una malattia monogenica, se può trasmetterla ai propri figli e da quale genitore l’ha ereditata”. Sono domande importanti soprattutto nella pratica quotidiana per dare risposte ai malati e alle loro famiglie. Fino ad ora abbiamo approfondito alcuni aspetti della genetica molecolare, sufficienti per poter affrontare l’analisi di diverse patologie monogeniche; adesso dobbiamo andare a richiamare alcuni concetti della genetica formale. Questo è un capitolo che si riferisce alla trasmissione dei caratteri monogenici; per affrontare questo argomento dobbiamo richiamare Mendel e le divisioni cellulari. Nell’ambito delle divisioni cellulari, in genetica siamo prevalentemente interessati alla meiosi. In questa diapositiva, però, c’è un confronto super semplificato tra meiosi e mitosi proprio per esaltarne le differenze. Innanzitutto, è rappresentata una cellula semplificata (non è una cellula umana) che presenta nel suo corredo diploide, un totale di 4 cromosomi [corredo aploide, 2 cromosomi; cromosoma grande e cromosoma piccolo]. In ogni coppia di cromosomi, uno è stato colorato di verde e l’altro di grigio, per distinguere il fatto che si tratta di cromosomi omologhi e non uguali identici (sono simili per contenuti genico, ma con differenze nucleotidiche che sono state esaltate con colori diversi). Abbiamo un cromosoma più grande che è costituito da una coppia di omologhi, quello grigio e quello verde, e un cromosoma più piccolo, anch’esso con un cromosoma a righe grigie e uno verde. Prima di una qualsiasi divisione cellulare, avviene la replicazione del DNA: ciascun cromosoma a questo punto risulta costituito da due cromatidi fratelli uniti a centromero. Poi avviene anche la perdita della membrana nucleare e la condensazione dei cromosomi. Vediamo quali sono le principali differenze tra la mitosi e la meiosi. Innanzitutto, a cosa serve la meiosi? Perché interessa tanto ai genetisti? Qual è la funzione della meiosi? Quali sono le cellule che vengono prodotte alla fine della meiosi? La meiosi serve a produrre i gameti (o cellule germinali). Questi gameti hanno una caratteristica importante da un punto di vista genetico, sono aploidi. Quindi nella meiosi partiamo da una cellula che è diploide e otteniamo quattro cellule apolidi. Questo è importante perché l’unione di due gameti aploidi, porterà nuovamente alla costituzione di una cellula diploide (che è uno zigote) e quindi alla generazione di un nuovo organismo (sempre diploide) che manterrà il numero di cromosomi tipico della propria specie. Se ciò non avvenisse e quindi se i gameti fossero diploidi, ora saremmo pienissimi di cromosomi. Questo meccanismo evolutivo è di fondamentale importanza.
Lo scopo principale della mitosi è quello di produrre delle cellule che siano il più possibile uguali alla cellula madre. Da un punto di vista genetico devono sempre essere delle cellule diploidi e devono continuare a svolgere lo stesso compito della cellula madre. L’accrescimento di un organismo avviene grazie alla mitosi. La sostituzione di cellule che vanno in senescenza; la morte di globuli rossi precoce porta a una proliferazione del midollo e quindi interviene ancora la mitosi. Quindi accrescimento di un organismo, sostituzione di cellule morte fisiologicamente o dovute magari anche a una ferita, sono quindi varie le applicazioni della mitosi. a. In questa diapositiva c’è almeno un’altra differenza tra mitosi e meiosi molto evidente. Nella meiosi abbiamo due divisioni cellulari, c’è la prima divisione meiotica e la seconda divisione meiotica; nella mitosi c’è un’unica divisione cellulare. I cromosomi hanno un destino diverso. Nella prima divisione meiotica, i cromosomi si dispongono in maniera diversa nella mitosi e nella meiosi: nella prima divisione meiotica i cromosomi si appaiano e si forma la coppia di cromosomi omologhi [ciascun cromosoma riconosce nella cellula il proprio omologo, sulla base della dimensione, posizione del centromero…] e si forma un’impalcatura proteica che li tiene uniti per un certo periodo più o meno lungo. Nella mitosi tutti i cromosomi si dispongono nella cellula in una fila a singolo cromosoma e non a coppie. Perché i cromosomi si devono appaiare nella prima divisione meiotica? La funzione dell’appaiamento, che se si è conservata vuol dire che è importante, è che successivamente avviene il crossing over : uno scambio reciproco di materiale genetico tra cromosomi omologhi. Alla fine, quando termina la prima divisione meiotica, avrò dei cromosomi non ricombinanti (tutti verdi/grigi nell’esempio) e anche cromosomi ricombinanti, derivanti dall’incrocio (= crossing) tra cromatidi non fratelli che si scambiano materiale perché avviene una rottura della doppia elica del DNA, uno scambio e una riparazione; il tutto porta poi a dei cromatidi che hanno una parte grigia, oltre a una parte verde (sempre riferito alla figura). In ogni coppia di omologhi avviene almeno un crossing over. Perché possa avvenire, i cromosomi devono essere appaiati [generalmente avviene tra cromosomi omologhi, ma possono esserci rari eventi di ricombinazione anche tra cromosomi non omologhi, ma la ricombinazione deve normalmente avvenire tra cromosomi omologhi]. Più il cromosoma è grande, maggiori sono questi eventi di ricombinazione. Se non avviene almeno un crossing over, il passaggio successivo che porta alla formazione dei cromosomi, non avviene in maniera corretta; si possono formare cellule con un difetto di contenuto di cromosomi già alla prima divisione meiotica. L’appaiamento di cromosomi omologhi è rilevante per il crossing over e poi anche per un altro fenomeno importantissimo, che è la segregazione dei cromosomi. I cromosomi non si separano in maniera casuale, ma se tutto va come deve andare, si separano in maniera tale che in ciascuna delle due cellule figlie si separi un solo cromosoma della coppia di omologhi. Se si appaiano, le fibre del fuso possono intrattenere relazioni con il centromero: le fibre del fuso del polo superiore intratterranno relazioni con il centromero di un omologo e le fibre del fuso inferiore le intratterranno con quelle dell’altro omologo. Quando poi avverrà la depolarizzazione delle fibre del fuso, un cromosoma omologo sarà attratto verso il polo superiore e un cromosoma omologo verrà attratto verso il polo inferiore, quindi si separeranno in maniera corretta. Se non avvengono l’appaiamento e il crossing over, non si ha la separazione corretta e si possono poi avere delle cellule che hanno magari due cromosomi da una parte e nessuno dall’altra (di un certo omologo). Questo è un evento che può capitare ed è la causa di molti difetti cromosomici come le trisomie, perché la separazione dei cromosomi omologhi è praticamente la base della prima legge di Mendel.
La cellula viene spesso definita come il globo terrestre: ci sono i poli e c’è il piano equatoriale. Se questo avviene, significa che poi, quando le fibre del fuso prendono contatti con il centromero di questi cromosomi, alla fine della prima divisione meiotica avrò una cellula in questo caso che conterrà i due cromosomi verdi e, di conseguenza, l’altra cellula che conterrà i due cromosomi grigi. Una volta che avviene l’interazione con una parte della cellula, quella inferiore per i grigi e quella superiore per i verdi, ci sarà una separazione secondo quest’ordine. Se invece, come in questo caso, ho un cromosoma verde e un cromosoma grigio che si appaia verso il polo superiore (cromosoma più grande, chiamato
- e, di conseguenza, il grigio insieme al piccolo verde che va verso il polo inferiore, quando si separano avrò una cellula che presenta il cromosoma 1, quello verde di origine materna, insieme al cromosoma grigio 2, di origine paterna e, di conseguenza, 1 di origine paterna grande, insieme a quello piccolo di origine materna. Già con solo due cromosomi, dopo la prima divisione meiotica, ho quattro cellule che da un punto di vista genetico sono tutte cellule aploidi, ma che sono diverse tra di loro. L’assortimento casuale dei cromosomi omologhi è una elevatissima fonte di variabilità delle cellule gametiche che si vanno a produrre. Qui ne ho prodotte 4 diverse perché ho 2 cromosomi; nell’uomo, che ha 23 coppie di cromosomi, si possono avere 2^23 = circa 6 milioni di cellule possibili , tutte diverse tra loro. Quindi un individuo può formare, nel corso della sua vita nell’ambito delle diverse meiosi, 2^23 diverse combinazioni cromosomiche. In più a questo si aggiunge il crossing over, che può avvenire in maniera del tutto casuale. Quindi la variabilità dei gameti che un soggetto può produrre è praticamente tale che nessun gamete di un soggetto è uguale a un altro gamete prodotto nel corso della propria vita ad un altro gamete dello stesso soggetto. C’è un altro aspetto che può essere di interesse nell’ambito della meiosi nell’uomo che si riferisce ai tempi della gametogenesi. La meiosi appunto è fonte di variabilità ed è indispensabile per la produzione di gameti aploidi. La gametogenesi maschile e femminile presenta alcune differenze che sono argomento, così come la meiosi e la mitosi, non tanto della genetica; in genetica ci si interessa di alcuni aspetti, che sono quelli poi funzionali al discorso tipico della genetica, quindi il concetto di aploidia, di separazione dei cromosomi omologhi e il concetto di variabilità del patrimonio genetico delle cellule aploidi. Un altro aspetto importante riguarda un pochino alcuni dettagli della gametogenesi, cioè dove avviene la meiosi nel maschio e dove avviene nella femmina, e perché questo. La gametogenesi maschile avviene a livello dell’organo deputato alla produzione di cellule riproduttive che è il testicolo, dove ci sono cellule progenitrici della linea germinale (si chiamano spermatogoni) che inizialmente si dividono per divisione mitotica, perché devono produrre tante diverse cellule progenitrici dei gameti. Ad un certo punto gli spermatogoni subiscono un accrescimento e un differenziamento, formano lo spermatocita primario che è una cellula ancora diploide. Gli spermatogoni, come lo spermatocita primario, sono cellule diploidi (2n). È lo spermatocita primario che va incontro a meiosi e produce, nella prima divisione meiotica due spermatociti secondari (cellule già aploidi, ma con contenuto di DNA ancora doppio); nella seconda divisione meiotica , invece, abbiamo la formazione di un totale di quattro spermatidi che sono aploidi e che poi maturano e diventeranno, con la formazione della coda, spermatozoi maturi.
L’altra differenza riguarda il tempo: la gametogenesi maschile è piuttosto rapida , dura in termini di giorni, 48 giorni e poi c’è ancora la differenziazione degli spermatidi in spermatozoi maturi, però tutto avviene in termini di giorni e avviene a partire dalla pubertà e può durare tutta la vita; si possono produrre diversi milioni di spermatozoi nel corso della vita. GAMETOGENESI FEMMINILE Nella femmina , l’ovaio è l’organo e le cellule progenitrici di gameti si chiamano oogoni, che si dividono per mitosi, perché se ne devono produrre tante. L’oogonio poi si accresce e si differenzia e diventa un oocita primario, che è una cellula diploide che inizia la prima divisione meiotica. Già nella prima divisione meiotica vediamo delle differenze rispetto alla gametogenesi maschile: la differenza è che nel maschio alla prima divisione meiotica si formano due spermatociti secondari uguali (con riarrangiamento cromosomico, ma da un punto di vista del fenotipo due cellule uguali), nella femmina si forma quello che si chiama l’oocita secondario (analogo allo spermatocita secondario) e però si forma una cellula più piccola, che è sempre aploide e che si chiama globulo polare. Il globulo polare è una cellula che non è destinata a diventare un gamete maturo. Avviene la seconda divisione meiotica, il globulo polare può dividersi o non dividersi, dipende. L’oocita secondario si divide in un ovulo maturo e di nuovo in un globulo polare secondario. La divisione è asimmetrica soprattutto per quanto riguarda il contenuto citoplasmatico, non tanto per quanto riguarda invece il contenuto genetico; da un punto di vista genetico sono entrambe cellule aploidi. Di fatto, ad ogni meiosi nella femmina si produce un gamete maturo (ovulo) e poi due o tre globuli polari, mentre nel maschio abbiamo quattro spermatociti e quindi spermatoz oi maturi a partire da una cellula progenitrice dei gameti che va incontro a meiosi. Qui la tempistica ha un impatto anche su alcuni aspetti genetici, come alcune malattie genetiche come poi vedremo. Innanzitutto, inizia molto prima la meiosi ; inizia nel terzo mese di vita embrionale e si ferma alla prima divisione meiotica, non prosegue alla nascita neanche la prima divisione meiotica. Gli oociti primari della femmina entrano in meiosi al terzo mese di vita embrionale e si fermano, coi cromosomi omologhi appaiati con il crossing over, alla nascita e vi rimangono fino all’età pubere quando, mensilmente, in corrispondenza di ogni ciclo mestruale riprende la meiosi. Quindi la meiosi può durare 13/14 anni minimo, fino a 45/50 anni, a seconda di quando riprende mensilmente. Ha una durata, in termini di anni, lunghissima. L’espulsione del secondo globulo polare avviene solo se c’è la fecondazione dell’ovulo, quindi, può completarsi o no (se non avviene la fecondazione). In ogni caso, la ripresa della meiosi a livello della prima divisione meiotica avviene solamente in corrispondenza del ciclo mestruale e quindi riprende alla pubertà. Questo ha un impatto importante perché ci sono dei difetti di separazione di cromosomi omologhi che sono più frequenti nella linea femminile che in quella maschile; non sappiamo la ragione di questo precisamente, però possiamo sapere che può essere una conseguenza di questa lunga fase di meiosi dove i cromosomi omologhi rimangono appaiati per anni.
Esempio dell’AB0. Partendo da questo esempio, possiamo poi seguire questa metodica analitica ed applicarla a qualunque quesito che si riferisce anche a malattie monogeniche. Il tipico quesito è: ho una coppia di soggetti in cui la mamma è di gruppo AB e il papà è di gruppo
0. Voglio sapere quali sono il genotipo e il fenotipo dei figli e qual è la proporzione. So che anche a memoria sapreste rispondere, ma vi invito lo stesso a seguire il metodo perché può essere molto utile poi per risolvere qualsiasi quesito, non solo sull’AB0. Il metodo suggerito è: una volta che mi si presenta questo quesito, io ho sempre di fronte dei fenotipi, cioè delle persone che manifestano un carattere o non lo manifestano, dei quali però devo come prima informazione definirne il genotipo, perché devo arrivare a ricostruire le informazioni genetiche presenti sui due cromosomi omologhi e applicare poi la prima legge di Mendel (la segregazione dei cromosomi omologhi) e capire che gameti producono. Quindi il primo passaggio è dal fenotipo al genotipo. Questo è un esempio super semplice, per cui la mamma che ha fenotipo AB, avrà genotipo AB; il papà che è gruppo 0, avrà genotipo 00. Quindi vuol dire che la mamma ha sul cromosoma l’allele A e sull’altro cromosoma l’allele B, e il papà ha su tutti e due i cromosomi l’allele 0. La mamma produrrà il 50% di gameti A e il 50% di gameti B, nel corso della sua vita; nel papà, i due cromosomi omologhi si separeranno, ma i suoi gameti saranno tutti di tipo 0 (perché è un omozigote). A questo punto possono avvenire tutte le possibili combinazioni: il gamete maschile di gruppo 0 può incontrare il gamete femminile di gruppo A o può incontrare un gamete femminile di gruppo B. Questo sistema di incrocio io lo posso esemplificare facendo il disegnino dei cromosomi, oppure lo posso fare usando un antico strumento che è il quadrato di Punnet , che è un metodo efficacissimo per rispondere a un quesito come quello che vi ho appena presentato. Il quesito che ci verrà posto non sarà tanto “quali sono genotipo e fenotipo dei figli di una coppia con un determinato gruppo sanguigno?”, ma vi verrà chiesto “qual è la probabilità che questa coppia abbia un figlio malato?”. Prima cosa, dal fenotipo passare al genotipo; ricavare il genotipo, descrivere i gameti che vengono prodotti (con il quadrato di Punnet).
Nel nostro caso, descrivo i gameti che verranno prodotti: la mamma 50% gameti A e 50% gameti B, il papà descrivo due volte 0. Poi faccio venire il quadrato di Punnet, che è un modo più semplice per effettuare incroci e la produzione di zigoti. Si possono qui produrre: lo zigote A0 e lo zigote B0. Formo i quattro possibili zigoti, che in questo caso sono uguali a due a due, perché il papà è monozigote. Posso quindi dire che i figli avranno genotipo 50% A0 e 50% B0. Il fenotipo, quindi, sarà 50% A e 50% B. È indifferente indicare 50% = ½ = 2 su 4. Una volta che si definisce chiaramente il genotipo, è poi praticamente impossibile sbagliare la risposta. Altro possibile quesito: mamma gruppo A e papà gruppo B. PROBLEMA: se non ho altre informazioni non posso definire in maniera univoca il genotipo, perché un soggetto con fenotipo A può avere genotipo AA o A0. Tuttavia, se aggiungo l’informazione “entrambi hanno almeno un genitore di gruppo 0”, posso risalire al genotipo. I genotipi sono A0 e B0 perché il genitore 00 ha trasmesso a entrambi un allele 0, mentre se sono di fenotipo A, vuol dire che l’allele A l’ha trasmesso l’altro genitore e quindi questi sono individui eterozigoti. In questo caso, qual è la proporzione di figli? I genotipi che si possono produrre sono tutti diversi, quattro possibili zigoti: ¼ AB, ¼ BO, ¼ A0, ¼ 00. Ho fenotipi che sono tutti diversi, per esempio ho la presenza del fenotipo 0, che non era presente nei genitori. Quindi ricavare sempre il genotipo; nei fenotipi recessivi è semplice, in quelli dominanti devo capire se si tratta di un omozigote o di un eterozigote. Una volta ricavato il genotipo possiamo costruire il quadrato di Punnet, dove inseriremo la proporzione di gameti materni e paterni e la formazione dei possibili zigoti che si possono formare, che sono sempre 4 e che possono essere uguali o diversi.
A B
0 A0 B
0 A0 B
A 0
B AB B
0 A0 00
Le malattie rare sono molto difficili da gestire da un punto di vista medico e diagnostico anche perché molto spesso non ci sono i farmaci per curarle. In passato, è stato creato un farmaco apposito per una bambina affetta dalla malattia di Batten che è stato chiamato MILASEN (in onore della bambina malata). Genereviews è un catalogo, o database, che sarà importante consultare in futuro. L’ALBERO FAMILIARE La prima cosa che si deve fare quando si sospetta la presenza di una malattia genetica è disegnare l’ albero familiare perché esso può offrire dei primi indizi riguardanti la patologia come la modalità di trasmissione, se la malattia sia recessiva o no, se essa riguardi i cromosomi sessuali o quelli autosomici e anche quali parenti all’interno della famiglia siano a rischio. L’albero genealogico si disegna attraverso dei simboli universali, si tratta di una esiste una convenzione internazionale per disegnarli. Il maschio si disegna col quadrato mentre la femmina col cerchio e se qualcuno di questi presenta la malattia si annerisce il rispettivo quadrato o cerchio. Queste due possibilità (bianco/nero) si riferiscono esclusivamente al fenotipo e infatti la rappresentazione dell’albero familiare non garantisce che generalmente un soggetto non abbia un allele mutato dal momento che essa non è correlata al genotipo. Il matrimonio tra maschio e femmina si disegna con una riga orizzontale, se il matrimonio avviene tra due consanguinei si mettono due linee orizzontali etc. Dal matrimonio originano delle righe verticali che vanno a indicare i figli.
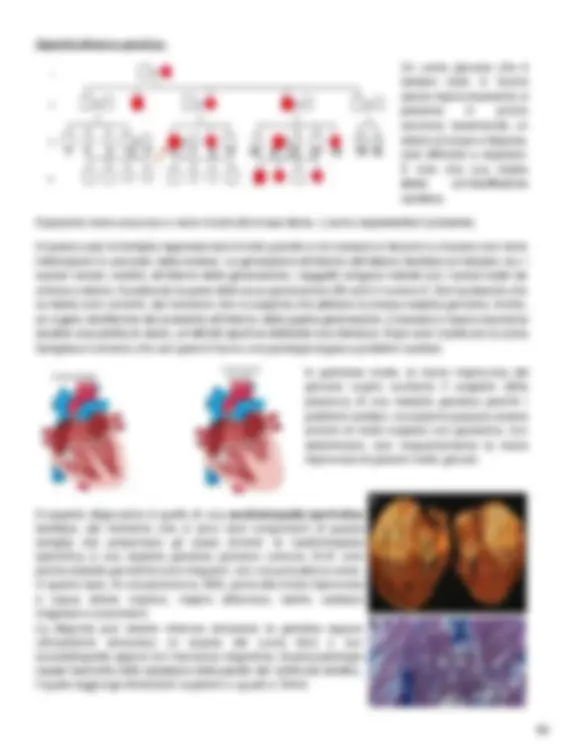
Quando è necessario indicare tanti figli non affetti dalla malattia essi si possono rappresentare con il simbolo corrispondente al loro sesso, contenente al suo interno il numero di figli presenti. Un soggetto deceduto viene indicato con una riga trasversale sopra il simbolo corrispondente. Il simbolo del rombo indica un individuo di sesso non noto, questo si usa prevalentemente negli studi genetici sulle malattie rare; poiché per motivi di privacy si preferisce non indicare se il paziente è maschio o femmina, in modo che la famiglia non sia riconoscibile. Allo stesso modo, durante una gravidanza in corso, il nascituro con sesso ignoto si rappresenta con un rombo tratteggiato. Per indicare due gemelli si disegnano due righe che originano dallo stesso punto e, nel caso in cui essi siano omozigoti, i due gemelli vengono uniti tra di loro attraverso un’ulteriore riga orizzontale. Similmente, nel caso in cui l’omozigosità sia incerta, si sostituisce la riga con un punto di domanda. La freccia indica il probando cioè il soggetto dal quale la nostra indagine parte (per esempio il primo malato che ci chiede consulenza genetica o anche il primo sano che si rivolge a noi per lo stesso motivo). Quando si inizia l’indagine sulla malattia genetica si disegna sempre l’albero per comprendere le linee di trasmissione. È la persona con la quale si inizia l’approfondimento. La prima cosa che devo fare in quanto medico genetista è quello di ricostruire il probando cioè l’albero genealogico e descriverlo. Questo è importante perché può dare indicazioni sulla modalità di trasmissione di una malattia. Possiamo infatti trovarci davanti ad una malattia con modalità di trasmissione nota oppure ad una malattia nuova e mai analizzata prima. Quest’albero, inoltre, ci permette di capire se nella famiglia ci sono stati altri casi e chi può aver ereditato la stessa malattia o chi può essere considerato a rischio. MALATTIE AUTOSOMICHE DOMINANTI Iniziamo dalle malattie autosomiche dominanti che corrispondono alla maggior parte delle malattie genetiche del catalogo OMIM (parliamo quindi di diverse migliaia). Verranno di seguito analizzati sia gli aspetti di genetica formale che quelli di genetica molecolare. Approfondimento genetico: Un uomo di 45 anni si reca al pronto soccorso presentando segni tipici dell’infarto (dolore al petto, debolezza). Viene curato e dopo alcuni giorni risulta stabile ma vengono comunque eseguiti ulteriori accertamenti poiché la patologia è sorta in un individuo di giovane età (di solito l’infarto colpisce dopo i 60 anni). Se l'infarto è accompagnato da una manifestazione troppo precoce (come in questo caso) potremmo aspettarci un difetto genetico di tipo monogenico.
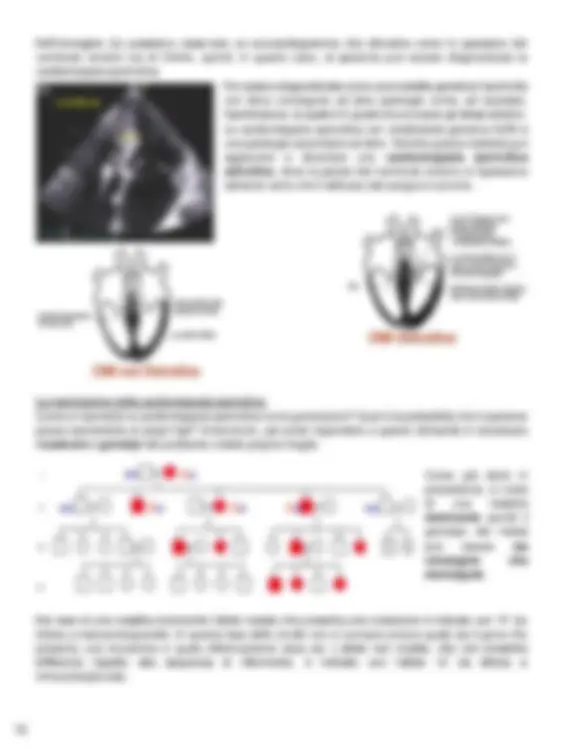
Approfondimento genetico: Un uomo giovane che è sempre stato in buona salute improvvisamente si presenta in pronto soccorso lamentando un dolore al torace e dispnea, cioè difficoltà a respirare. È noto che sua madre abbia un’insufficienza cardiaca. Il paziente viene soccorso e viene ricostruita la sua storia. L’uomo rappresenta il probando. In questo caso la famiglia rappresentata è molto grande e non sempre si riescono a ricavare così tante informazioni in una sola visita medica. Le generazioni all’interno dell’albero familiare si indicano con i numeri romani, mentre, all’interno della generazione, i soggetti vengono indicati con i numeri arabi da sinistra a destra. Il probando fa parte della terza generazione (III) ed è il numero 6. Sia il probando che la madre sono anneriti, dal momento che si sospetta che abbiano la stessa malattia genetica. Inoltre, un cugino diciottenne del probando all’interno della quarta generazione è deceduto improvvisamente durante una partita di calcio, un’attività sportiva dilettante ma intensiva. Dopo aver ricostruito la storia famigliare è emerso che vari parenti hanno una patologia legata a problemi cardiaci. In particolar modo, la morte improvvisa del giovane cugino aumenta il sospetto della presenza di una malattia genetica perché i problemi cardiaci, nonostante possano essere sintomi di molte malattie non genetiche, non determinano così frequentemente la morte improvvisa di pazienti molto giovani. Il sospetto diagnostico è quello di una cardiomiopatia ipertrofica familiare, dal momento che ci sono tanti componenti di questa famiglia che presentano gli stessi sintomi: la cardiomiopatia ipertrofica è una malattia genetica piuttosto comune (N.B: solo poche malattie genetiche sono frequenti, con una prevalenza come, in questo caso, di una persona su 500), porta alla morte improvvisa e causa dolore toracico, respiro affannoso, battito cardiaco irregolare e svenimenti. La diagnosi può essere ottenuta attraverso la genetica oppure clinicamente attraverso un esame del cuore fatto o con ecocardiografia oppure con risonanza magnetica. Questa patologia causa l’aumento dello spessore della parete del ventricolo sinistro, il quale raggiunge dimensioni superiori o uguali a 15mm.
Nell’immagine (b) possiamo osservare un ecocardiogramma che dimostra come lo spessore del ventricolo sinistro sia di 24mm, quindi, in questo caso, al paziente può essere diagnosticata la cardiomiopatia ipertrofica. Per essere diagnosticata come una malattia genetica l’ipertrofia non deve conseguire ad altre patologie come, ad esempio, l’ipertensione, la quale è in grado di provocare gli stessi sintomi. La cardiomiopatia ipertrofica con ereditarietà genetica NON è una patologia secondaria ad altre. Talvolta questa malattia può aggravarsi e diventare una cardiomiopatia ipertrofica ostruttiva , dove la parete del ventricolo sinistro si ispessisce talmente tanto che il deflusso del sangue è ostruito. La trasmissione della cardiomiopatia ipertrofica Come si trasmette la cardiomiopatia ipertrofica tra le generazioni? Qual è la probabilità che il paziente possa trasmetterla ai propri figli? Innanzitutto, per poter rispondere a queste domande è necessario ricostruire i genotipi del probando e della propria moglie. Come già detto in precedenza, si tratta di una malattia dominante quindi il genotipo dei malati può essere sia omozigote che eterozigote. Nel caso di una malattia dominante l'allele malato che presenta una mutazione è indicato con “A” (la lettera a maiuscola/grande). In questa fase dello studio non si conosce ancora quale sia il gene che presenta una mutazione e quale effettivamente essa sia. L’allele non mutato, che non presenta differenze rispetto alla sequenza di riferimento, è indicato con l’allele “a” (la lettera a minuscola/piccola).
La frequenza in genetica coincide con la prevalenza in epidemiologia. La frequenza è il numero di malati per unità di popolazione. La malattia autosomica dominante più frequente è l’otosclerosi dominate, una forma di sordità. Ciascuna malattia ha il proprio numero univoco di catalogo OMIM.
PERCHÉ UN ALLELE CODIFICA PER UN CARATTERE DOMINANTE?
Perché una mutazione in eterozigosi è sufficiente per determinare una malattia? Quando si verifica una mutazione, essa può causare effetti differenti:
- perdita di funzione della proteina codificata;
- guadagno di funzione : il conferimento di una nuova funzione non è compatibile con un quadro fisiologico , quindi, causa una malattia;
- dominante negativo. Una mutazione può essere dominante sia in caso di perdita che di guadagno di funzione, mentre un carattere recessivo è quasi sempre accompagnato da perdita di funzione. Infatti, nel caso di mutazioni di carattere recessivo, la proteina codificata dal gene mutato non si esprime e quindi nel soggetto eterozigote si ha solo il 50% di proteina normale, quella espressa dall’altro allele sano. Questa condizione viene anche detta " aplosufficiente ", dato che la proteina sarà prodotta in quantità minore rispetto ad un soggetto normale, ma essa potrà essere comunque sufficiente per le normali funzioni cellulari senza provocare nessuna manifestazione clinica. Se invece il 50% della proteina non è sufficiente affinché la cellula possa svolgere tutte le sue normali funzioni, si avrà una condizione di " aploinsufficienza " e la mutazione sarà di carattere dominante, c’è manifestazione clinica. 1 - APLOINSUFFICIENZA
- Nel caso dell’ ipercolesterolemia , il difetto è il gene per la LDLR (LDL è il ligando, Low Density Lipoprotein e R è il recettore). Questa LDLR sarebbe una proteina transmembrana espressa sulla superficie di molte cellule, che viene soprattutto esplicata nell’ambito del fegato e del surrene. Questa proteina (rappresentata in verde nella schema sotto) lega le LDL e crea un complesso lipidico e proteico a bassa densità all’interno del quale è contenuto il colesterolo. Tale legame è consentito dalla proteina Apo B-100, l’unica proteina del complesso LDL. NB. Nel sangue ci sono vari complessi lipoproteici distinti in base alla densità, dunque possono essere ad alta densità, media, bassa, ecc Questo recettore con le LDL legata viene poi internalizzato nella cellula dove avviene un processamento in cui il colesterolo viene esterificato grazie all’enzima ACAT e depositato nei tessuti. Abbiamo quindi un processamento mediato da endocitosi e avviene una metabolizzazione del complesso. Il recettore viene riportato sulla membrana cellulare pronto per legare una nuova LDL sempre tramite Apo B-
- Viene sottratto il colesterolo dal sangue e rilasciato nei tessuti. Fino all’ 80% delle LDL in realtà lega anche le IDL (intermedie DL), sempre con internalizzazione per mezzo di endocitosi.
Il probando, a livello dell’esone che codifica per il domain C9, presenta, nel codone 1096, una sostituzione nucleotidica che codifica per un codone di stop prematuro. Si tratta quindi di una mutazione nonsense nel gene MYBPC3. La proteina non viene prodotta per il Nonsense Mediated Decay. La proteina C legante la miosina viene quindi espressa solo parzialmente nel caso di pazienti eterozigoti: l'allele “a” è sano, mentre l'allele “A”, colpito da una mutazione troncante che determina una perdita dell'espressione, codifica per una proteina che non viene espressa, dunque, in molti tratti del muscolo cardiaco questa proteina è assente. Si verifica, dunque, una condizione di aploinsufficienza. Infatti, la produzione solo del 50% di questa proteina, non è sufficiente per una normale manifestazione fenotipica, ed è per questo che la cardiomiopatia ipertrofica è una malattia monogenica autosomica di carattere dominante. Quanto visto fino ad ora appartiene ad una situazione di aploinsufficienza perché non è sufficiente la quantità di proteina prodotta in modo corretto. Altre malattie con condizione di aploinsufficienza sono la Sindrome di Waardenburg che è una sordità neurosensoriale che si manifesta già nei primi anni di vita e il malato presenta una diversa pigmentazione della pelle o capelli (ciuffo bianco) e occhi molto distanziati tra di loro (telecanto o canthorum). Come faccio a vederlo? Si fanno analisi della dimensione della circonferenza cranica, dell'altezza delle orecchie, tutta una serie di valutazioni antropometriche per verificare se il pz mostra caratteristiche che rientrano o meno nel range. In questo caso è coinvolto il gene PAX3 che ha un dominio che lega il DNA per cui si ha perdita di funzione che si manifesta anche in eterozigosi. È una malattia autosomica dominante. Di solito queste mutazioni portano ad una perdita di funzione ma può anche succedere che causino un acquisto di funzione. Lo vediamo nelle malattie monogeniche della linea “germ line”. Ci sono molte mutazioni somatiche sono a guadagno di funzione.
2 - ACQUISIZIONE DI FUNZIONE
L’ acondroplasia è un esempio di malattia causata da una mutazione che porta ad un guadagno di funzione cioè all'espressione di una proteina normalmente non funzionante. Essa è una malattia genetica che colpisce circa un individuo su 15 000/40 000 nati vivi, caratterizzata da una bassa statura non armonica, dove sono soprattutto le ossa lunghe ad avere un difetto di allungamento, quindi principalmente quelle degli arti. Il difetto è apprezzabile già alla nascita, e talvolta si osserva grazie a delle ecografie durante la gravidanza. Altre caratteristiche che possono essere osservate sono la radice del naso infossata, la fronte prominente, le mani corte (anche dette a tridente) e la colonna vertebrale esageratamente curvata in una lordosi lombare, condizione che in alcuni casi durante l'infanzia può causare arresto respiratorio, secondario a una stenosi della giunzione cranio-cervicale. L’acondroplasia è una patologia autosomica dominante causata da una mutazione del gene codificante il FGFR3 (recettore del fattore di crescita dei fibroblasti). È un recettore transmembrana contenente un dominio tirosin chinasico, presente in tutti i componenti della famiglia di questi recettori. Nel caso dell’acondroplasia, si riscontra una mutazione di tipo missense presente nel dominio che attraversa la membrana plasmatica. Questa mutazione porta alla sostituzione di una glicina da parte di un’ arginina sul codone 380 (Gly380Arg). Questi due aminoacidi sono piuttosto diversi; la glicina è un piccolo aminoacido polare, mentre l'arginina è un aminoacido carico , e si trova nel dominio transmembrana, in una posizione dove una carica può disturbare il normale funzionamento della cellula. Normalmente la funzione di questo recettore è di signaling negativo di accrescimento , funzione esplicata dal legame con il ligando (FGF). Quindi, il legame FGFR3-FGF trasduce nella cellula un segnale di diminuzione della velocità di accrescimento, un processo finemente regolato. In seguito alla mutazione missense che sostituisce ad una glicina un'arginina, questo recettore rimane sempre costitutivamente attivo , indipendentemente dal legame con il ligando, e manda costantemente un segnale di accrescimento negativo alla cellule della cartilagine, inibendo precocemente la crescita delle ossa lunghe e causando anomalie nel differenziamento anche delle altre ossa, come quelle del cranio.