Scarica Microbiologia generale e più Appunti in PDF di Microbiologia solo su Docsity!
MICROBIOLOGIA GENERALE ERMANNO FEDERICI
APPUNTI INTEGRATI DA SLIDE E LIBRO : “BIOLOGIA DEI MICRORGANISMI” DEHO’ e GALLI Tutti gli organismi viventi sono costituiti da cellule, le unità strutturali a cui attribuiamo le proprietà essenziali del vivente. Alcuni organismi sono unicellulari, altri pluricellulari, ma la maggior parte degli esseri viventi sulla terra sono unicellulari e insieme ad alcuni organismi pluricellulari costituiscono i MICRORGANISMI. Le cellule hanno tutti una propria identità, garantita dalla membrana plasmatica, che le delimita e le isola, ma allo stesso tempo le mette anche in comunicazione con l’ambiente esterno. Attraverso la membrana infatti la cellula è in grado di scambiare molecole in modo selettivo con l’esterno, di percepire le variazioni dell’ambiente e di mettere in atto processi di adattamento, che consentono di mantenere la propria integrità, l’ OMEOSTASI. Nel lungo periodo però, la capacità di mantenimento di una cellula è dato solo dal fatto che sono capaci di riprodursi, generando cellule identiche alla cellula madre, infatti secondo la scienza ogni cellula esistente deriva dalla divisione di un’altra cellula. Una cellula che non è in grado di replicarsi è destinata a scomparire. L’omeostati e la riproduzione cellulare richiedono che materiale non cellulare venga importato dall’ambiente esterno nella cellula, elaborato e trasformato in strutture cellulari e infine in nuove cellule. Tutto questo necessita di un continuo approvvigionamento di materia ed energia; queste operazioni costituiscono il metabolismo e devono avvenire in modo ordinato e organizzato. Perciò tutte le cellule sono dotate di un apparato che contiene informazioni necessarie alla vita della cellula: PROTEINE e DNA. Tramite la replicazione di quest’ultimo e alle proteine, che vanno sanno svolgere tutte le operazioni richieste per mantenere e replicare una cellula, si riproducono le informazioni genetiche che vengono trasmesse dalla cellula madre alla cellula figlia, mantenendo le caratteristiche specifiche. EVOLUZIONE TASSONOMICA L’uso del microscopio, da cui deriva il termine “microrganismi”, ha permesso l’identificazione di numerosi esseri viventi che non rientravano nei classici regni: animali e piante. Nel 1866 venne proposto un regno dal nome “ PROTISTI ” dove potevano rientrare tutti gli altri esseri viventi. Successivamente alla scoperta del microscopio elettronico, si venne a conoscenza di cellule che non erano per niente uguali dalle cellule del regno animale o delle piante. Le cellule di quest’ultimi avevano un nucleo delimitato da membrana e un complesso sistema di organelli, mentre altre no; fu così che vennero coniati i nomi “procarioti” ed “eucarioti” per distinguere questi diversi esseri viventi composti da diverse cellule. Successivamente i procarioti presero l’indipendenza dai protisti diventando il regno “ MONERA ” e un ulteriore scissione dal regno protista avvenne con l’avvento del regno dei “ FUNGH I”. Tuttavia il regno dei procarioti, comunemente chiamati batteri, non poteva vivere all’interno di una sola categoria, in quanto avevano caratteristiche peculiari molto diverse tra loro. Venne quindi proposti di riclassificare questo mondo in 3 domini: “ Eukarya ” che comprende piante, animali, funghi e protisti; “ Bacteria ” e “ Archea ” che derivano dalla scissione dei procarioti. ORGANISMI MODELLO E DIVERSITA’ MICROBICA Le nostre conoscenze di processi, fenomeni e meccanismi biologici derivano dallo studio intensivo e specializzato di pochi organismi, detti “modello”, che estendiamo anche ad altri organismi su cui non sono stati condotti studi specifici. Organismi modello molto utilizzati sono per esempio il SACCHAROMYCES CEREVISIAE, diventato il sistema modello per eccellenza della cellula eucariotica e il batterio, per le cellule procariotiche, ESCHERICHIA COLI, in cui sono stati scoperti meccanismi genetici che hanno facilitato lo studio e la manipolazione sperimentale.
Le specie microbiche sono le più numerose rispetto a gli altri organismi viventi e la loro diversità biologica è la più vasta che si conosca. Tale diversità biologia è necessaria per l’adattamento, che consente ai microrganismi di vivere negli habitat più disparati e utilizzare le più diverse fonti nutritive. Ovunque ci sia vita, ci sono i microrganismi, che siano batteri, archei o microrganismi eucarioti; la loro presenza è indispensabile per sostenere la vita di tutti gli altri organismi. LA CELLULA La caratteristica primaria che distingue le cellule eucariote da quelle procariotiche è costituita dal nucleo. Nella cellula eucariotica i cromosomi contenuti in un nucleo delimitato da membrana nucleare, mentre nella cellula procariotica tale membrana è assente e il materiale genetico, costituito da un solo cromosoma, è a diretto contatto con il citoplasma e forma una struttura chiamata nucleoide. Le cellule eucariotiche presentano organelli cellulari delimitati da membrane, assenti nelle cellule procariotiche; oltre ad essere più complesse le eucariotiche hanno anche dimensioni maggiori. Hanno però anche delle caratteristiche comuni, a cui corrispondono funzioni indispensabili: tutte le cellule presentano la membrana plasmatica , che separa il citoplasma dall’ambiente esterno. La composizione di quest’ultimo è simile per entrambe le tipologie di cellule: entrambi contengono i ribosomi necessari per svolgere la sintesi proteica. La divisione cellulare nei procarioti avviene in genere per scissione binaria. Durante questo processo il DNA viene replicato e ciascuna cellula figlia ne acquisisce una copia. Negli organismi eucarioti invece, la divisione cellulare, avviene per mitosi. Un’altra importante differenza è che nei procarioti non sono note modalità di riproduzione sessuale e non sono presenti processi di zigosi e meiosi. I microrganismi, tipicamente unicellulari, possono essere sia eucarioti (alghe, funghi e protozoi) sia procarioti (batteri e archei). LA CELLULA PROCARIOTA: MORFOLOGIA, DIMENSIONI E ORGANIZZAZIONE i batteri hanno generalmente dimensioni lineari nell’ordine del micron. Questo fa sì che il rapporto superficie/volume delle cellule procariote sia molto più elevato, con conseguenza sulla velocità con cui avvengono gli scambi molecolari tra cellula e ambiente, il trasporto di nutrienti, la divisione cellulare e anche l’evoluzione.
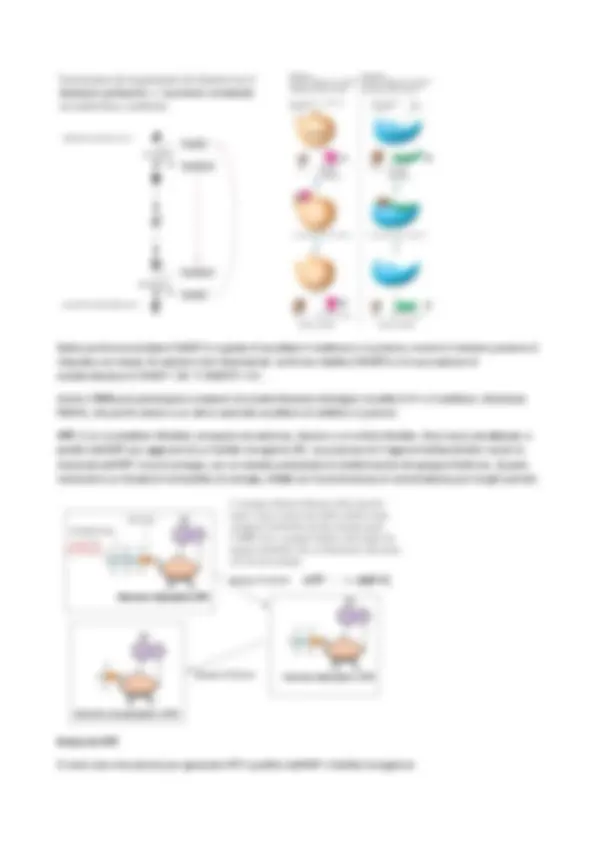
La morfologia di solito è propagata e mantenuta di generazione in generazione, ma sono noti casi anche di differenziamento morfologico in seguito a variazioni ambientali o del ciclo fisiologico. Forma e dimensioni hanno una grande importanza per le funzioni cellulari. La morfologia batterica è determinata dalla parte mureinica o sacculo, costituito da un'unica molecola, il PEPTIDOGLICANO , che avvolge la cellula. Il CITOPLASMA dei batteri Il citoplasma batterico è formato al 70% da acqua e contiene: genoma, RNA, ribosomi, enzimi e metaboliti. Non è compartimentalizzato come nelle cellule eucariotiche, infatti mancano gli organelli e le altre strutture delimitate da membrana, ma ciò non impedisce un’elevata funzionalità dei processi che si svolgono al suo interno. L’insieme della membrana plasmatica e di tutte le sue componenti interne viene definito PROTOPLASTO. Il citoplasma appare denso, trasparente ed elastico. Ha una struttura granulare, causata dall’elevata concentrazione di ribosomi. Tutte le macromolecole presenti all’interno del citoplasma hanno il compito di portare avanti la sintesi delle proteine e di altre componenti cellulari. Le proteine costituiscono circa il 20% del citoplasma, il 7-20% è costituito da RNA. RIBOSOMI : questi insieme all’RNA messaggero e al tRNA sono deputati alla sintesi delle proteine; essi possono essere anche debolmente legati alla membrana plasmatica: quelli che stanno all’interno del citoplasma sintetizzano proteine destinate a rimanere in quell’ambiente, mentre i ribosomi adesi alla membrana plasmatica producono proteine destinate ad essere trasportate all’esterno. Man mano che l’mRNA viene allungato ad opera dell’RNA polimerasi, i ribosomi iniziano a legarvisi dando luogo a strutture definite POLISOMI, legati al nucleoide tramite l’mRNA. NUCLEOIDE : i procarioti non possiedono un nucleo definito da una membrana e il DNA costituisce il loro genoma, che associato alle proteine va a costituire il nucleoide. GENOMA : è l’apparato che detiene, in tutte le cellule di qualsiasi organismo, l’informazione genetica necessaria alla loro biosintesi, riproduzione, omeostasi e adattabilità. Nei batteri esso è costituito da un’unica molecola circolare di DNA, che insieme alle proteine costituisce un unico CROMOSOMA , condensato in una struttura detta NUCLEOIDE. Il cromosoma è sempre presente ed è quasi sempre 1 ed è presente in singola copia. Oltre che nel genoma, l’informazione genetica può essere “arricchita” da elementi genetici accessori, i PLASMIDI. Essi non sono essenziali per la vita della cellula, possono essere più di uno e presenti in più copie; essi riescono a replicarsi in maniera autonoma dal cromosoma, infatti vengono detti elementi extracromosomici. La cosa interessante dei plasmidi è che sono a replicazione autonoma, si replicano autonomamente rispetto al batterio. Quando la cellula si divide per scissione binaria, deve replicare il proprio genoma per dare una copia di genoma identico alle sue figlie, perciò prima di dividersi ha due copie di cromosoma e due di plasmidi che però si replicano per conto proprio. Mentre il cromosoma viene ripartito perfettamente nelle
cellule figlie, i plasmidi non vengono ripartiti equamente. Le funzioni che riportano i plasmidi sono: resistenza agli antibiotici, la capacità di sintetizzare molecole che non servono direttamente al batterio ma che gli servono per colonizzare alcuni ambienti, hanno una funzione F, di fertilità, che gli permette di passare da una cellula all'altra in modo orizzontale (è la capacità di scambiarsi materiale genetico un po' come fanno i gameti). Il fatto che il genoma negli eucarioti si trovi nel nucleo, è dato dal fatto che il genoma eucariotico è molto grande e complesso, fatto da introni ed esoni. Quando viene trascritto a mRNA questo deve essere processato (per esempio tagliando via delle sequenze dette introni), poi diventa RNA maturo e successivamente viene tradotto. Nei batteri tutto è più semplice perché tutto il genoma è codificante (non ci sono introni ed esoni); non avviene lo splicing e trascrizione e traduzione avvengono nello stesso compartimento e nello stesso momento; infatti questo processo è molto rapido (si tratta di pochi minuti). Struttura del DNA batterico : è un'unica molecola di DNA circolare super avvolto, perché deve essere compatto, altrimenti non entrerebbe all’interno della cellula. È sempre associato alle proteine che si chiama ISTOLIKE. REPLICAZIONE DEL DNA L’apparato di replicazione, detto REPLISOMA , ha un’architettura simile nei procarioti e negli eucarioti e la sua funzione essenziale è duplicare in modo efficiente e fedele l’informazione contenuta nelle molecole di DNA. La differenza fondamentale tra i batteri e gli eucarioti è che mancando il nucleo, tutto avviene contemporaneamente nei batteri. La prima funzione del genoma senza la quale il genoma non verrebbe trasmesso alle generazioni successive è la replicazione del DNA. Il batterio si replica per scissione binaria: processo per cui di generazione in generazione da una cellula madre si ottengono delle cellule figlie geneticamente identiche alla cellula madre, in quanto tutte le sue strutture vengono replicate, compreso il genoma. Oltre al cromosoma anche tutti i plasmidi devono replicarsi in modo che lo stesso identico genoma vada alle cellule figlie. I meccanismi di replicazione posso essere 2: aum histon-like
come delle goccioline di grasso, simili alle strutture di cui si compongono gli adipociti. Invece alcuni batteri stoccano i poli-fosfati perché il fosfato serve come molecola per stoccare energia e alcuni batteri, rarissimi, riescono a farlo. Anche per il solfuro vale la stessa cosa. a. granuli idrossil-butirrato b. granuli di meta-poli-fosfati c. granuli di solfuro VESCICOLE GASSOSE : Un altro tipo di struttura interna al citoplasma dotata di membrana è rappresentata dalle vescicole gassose. Sono costituiti da una membrana proteica, hanno una struttura rigida e vuota → sono delle vere e proprie camere d'aria. Essi si trovano in alcuni batteri acquatici in grado di fare la fotosintesi. All'inizio queste strutture vennero confuse con i cloroplasti, ma in realtà sono strutture vuote, al cui interno c'è aria e permettono ai batteri di galleggiare, di catturare la luce e di conseguenza fare la fotosintesi. MEMBRANE E PARETI: il rivestimento delle cellule procariotiche Le cellule procariote son dotate di complesse strutture di rivestimento, che consentono loro di adattarsi a specifici habitat e di affrontare le diverse e difficili condizioni fisiche, chimiche e nutrizionali. La cellula è delimitata da una membrana plasmatica , che di norma è protetta da una parete , che assume strutture diverse in archea, nei batteri Gram positivi e Gram negativi. Inoltre molti procarioti, in aggiunta alla parete, possiedono ulteriori strati di rivestimento: strato S, capsula o glicocalice. Possiedono poi appendici di superficie come pili e flagelli , che consentono loro ulteriori proprietà per interagire con ambienti complessi.
Le diverse caratteristiche delle pareti consentono di distinguere i batteri in due gruppi fondamentali: Gram positivi e Gram negativi. Nella parete dei Gram negativi si osserva una vera e propria membrana con doppio foglietto di lipidi e glicolipidi con proteine associate detta membrana esterna (per cui la membrana plasmatica è anche detta membrana interna ). Nello spazio delimitato dalle due membrane, il PERIPLASMA , si trova un sottile strato di peptidoglicano , che avvolge il batterio come una maglia. La membrana esterna dei Gram negativi rappresenta una barriera contro il passaggio per diffusione di molti composti, conferendo cosi alla cellula una resistenza a sostanze tossiche e la capacità di adattarsi ad ambienti ostili. I batteri Gram positivi sono invece privi di membrana esterna e la loro parete è composta da uno spesso strato di peptidoglicano, con altre proteine di superficie. Questi batteri allora non possiedono un vero e proprio strato periplasmatico delimitato dalle due membrane, ma il termine periplasma sarà comunque usato per indicare lo spazio immediatamente esterno alla membrana plasmatica. Le due diverse composizioni della parete batterica stanno alla base della risposta, positiva o negativa, ad una specifica colorazione ideata da microbiologo Gram. Oggi si indica con il termine DIDERMI i batteri Gram negativi composti da 2 membrane e MONODERMI i batteri Gram positivi composti da 1 membrana. La membrana plasmatica nei Batteri La membrana plasmatica delimita il contenuto cellulare in uno spazio definito e separa la cellula dall’ambiente esterno. È fondamentale per conferire alla cellula la sua individualità e per regolare gli scambi tra cellula e ambiente; infatti le cellule interagiscono in modo complesso e selettivo con quest’ultimo. La membrana permette di acquisire ed eliminare rispettivamente nutrienti e sostanze di rifiuto o secernere molecole specifiche. Permette di mantenere costante e in uno stato organizzato il contenuto cellulare, indipendentemente dai cambiamenti esterni. Oltre a svolgere tutte queste funzioni di permeabilità selettiva , la membrana è la sede di importanti processi metabolici, che nelle cellule eucariotiche sono delegati a organelli specifici, come per esempio respirazione e fotosintesi. Grazie alla presenza di recettori specifici , la membrana è una struttura fondamentale per la trasduzione del segnale che consente alle cellule di recepire e rispondere ai diversi stimoli ambientali. La membrana è costituita da un doppio strato fosfolipidico a cui sono associate vari tipi di proteine. La maggior parte dei lipidi che le compongono sono molecole anfipatiche costituite da una testa polare e una coda idrofobica. La membrana plasmatica dei batteri e degli eucarioti è tipicamente costituita da FOSFOLIPIDI in cui la testa polare è rappresentata dal glicerolo 3-fosfato e la coda da due molecole di acidi grassi non ramificati ed esterificati al C1 e C2. Al fosfato esterificato in C3 possono essere legati gruppi funzionali diversi, dando origine quindi a molecole diverse. In ambiente acquoso, i fosfolioidi tendono a formare un doppio strato con le catene laterali degli acidi grassi che si associano tramite legami idrofobici, mentre le porzioni polari sono rivolte verso l’esterno. Questa è la disposizione termodinamicamente più stabile delle molecole lipidiche.
Il ruolo energetico della membrana : è svolto dalle proteine che permettono il passaggio di sostanze e dai lipidi che fungono da barriera. Nella membrana citoplasmatica avviene la catena di trasporto degli elettroni → una serie di enzimi associati alla membrana che sono dei trasportatori di elettroni. Questi trasportano elettroni ad un accettore finale (ossigeno) e contemporaneamente fungono da pompe protoniche che espellono protoni, ovvero delle cariche positive. Queste cariche positive si accumulano fuori dalla membrana e così facendo la membrana si carica; abbiamo una separazione di cariche e questa energizza la membrana. Questo ruolo detto " ruolo di conservazione di energia della membrana " è tipico dei batteri, gli eucarioti svolgono lo stesso processo ma sulla membrana mitocondriale. Questo ruolo è possibile grazie alle proteine di membrana e grazie ai lipidi (che mantengono separate le cariche). FUNZIONE DI TRASPORTO : La membrana è una barriera permeabile in modo selettivo, questo perché i trasportatori sono enzimi che interagiscono con il substrato in modo molto selettivo. Il substrato, nel caso del trasporto, è la molecola che viene trasportata. La quantità di trasportatori presenti è anche un modo per regolare il trasporto e per capire quindi la quantità di sostanze che la cellula fa entrare. Il trasporto di tutte le molecole richiede consumo di energia, perché per portare all'interno una sostanza la cellula spende dell’energia. I trasportatori funzionano permettendo alla cellula di assorbire solo ciò che gli serve. Se queste proteine fossero dei semplici fori succederebbe che la quantità di molecole e substrati trasportati sarebbe direttamente proporzionale alla quantità di molecole presenti all'esterno, cosa che non è possibile perché la cellula non è mai in equilibrio con l'ambiente esterno. Gli enzimi e quindi tutti i sistemi di trasporto, sono saturabili: all'inizio la molecola che funge da substrato entra e poi le concentrazioni di questo composto aumentano all'interno e gli enzimi non lo fanno più entrare. Il secondo modo per regolare il trasporto è che gli enzimi sono altamente specifici, quindi la cellula sintetizza solo la proteina di trasporto che le serve. La cellula non ha tutte le proteine di trasporto che esistono sempre presenti sulla membrana ma solo quelle che le servono: quelle presenti sulla membrana vengono sintetizzate solo quando c'è la necessità di portare un determinato metabolita all'interno (e poi ovviamente c'è un corredo proteico presente sulla membrana che è sempre presente). Un esempio è quello della capacità della cellula di usare glucosio come zucchero per ricavare energia insieme ad altri zuccheri come il fruttosio. Fu scoperto che se ai batteri diamo il glucosio usano quello, se gli diamo il lattosio usano questo. Mentre se gli diamo il glucosio e il lattosio insieme, i batteri usano il glucosio perché per usare il lattosio, la cellula ha bisogno di una serie di geni apposta per consumare il lattosio, tra cui il trasportatore del lattosio: "lac permeasi".
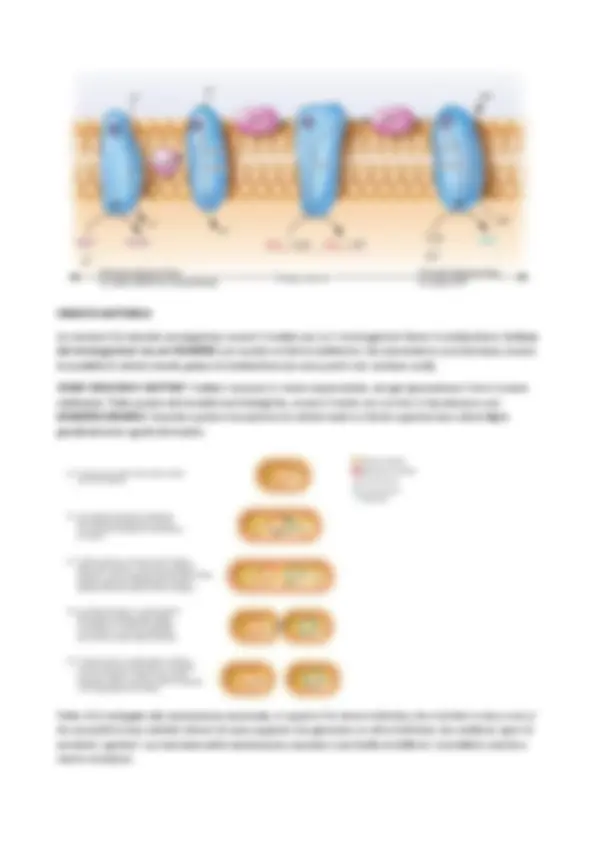
La capacità di sintetizzare vuol dire saper esprimere quei geni che codificano per quel trasportatore secondo le necessità della cellula, tutto ciò perché sono dei trasportatori specifici e non dei semplici pori (come nella parete in cui vi troveremo le porine). SISTEMI DI TRASPORTO Esistono tre sistemi di trasporto che funzionano in modo completamente diverso. Questi tre sistemi coinvolgono solo canali, oppure coinvolgono canali e delle attività enzimatiche accessorie, altrimenti coinvolgono dei canali e delle attività enzimatiche accessorie presenti in proteine associate alla membrana. Queste tre diversità di trasportare sostanze all’interno della cellula, ci spiegano 3 diverse modalità di dispendio di energia. L'unico sistema di trasporto che non consuma energia è la diffusione, che però è permessa solo all'acqua che può diffondere tramite la membrana, senza consumare energia, continuamente.
- TRASPORTO SEMPLICE : le proteine di trasporto si chiamano “ channels ” (=trasportatori) grazie a questo sistema, perché sono dei veri e propri canali che hanno il compito di schermare la parte idrofobica della membrana e di permettere quindi il passaggio di molecole. Sono dei trasportatori perciò altamente specifici e saturabili. Ci sono dei canali specifici per molecole come il potassio, il sodio e il fosfato. Questo consuma energia, perché la membrana è carica (positiva esternamente e negativa all’interno) e di conseguenza viene sfruttata la forza proton-motrice da questo tipo di trasporto. Osservando i trasportatori che siano uniporter, symporter o antiporter vediamo che trasportano cationi, che entrano essendo attratti dalle cariche negative interne e così facendo si consuma energia, perché le cariche vengono ribilanciate. Esistono poi dei sistemi di antiporter e symporter che permettono l'entrata di anioni, come per esempio il fosfato che è fondamentale e serve, ma ha una carica negativa. Il fosfato entra grazie ad un symporter, che possono trasportarlo perché insieme a lui entrano anche dei protoni. Questi ultimi, rientrando, permettono di cedere energia per far riaprire il passaggio. Anche molecole non cariche ma grandi come il lattosio possono entrare insieme ad un catione, questo è il meccanismo della “lac permeasi” (l’enzima di trasporto del lattosio” che è un symporter. Questo sistema è anche usato per espellere molecole cariche positivamente (così come quelle cariche negativamente non possono entrare liberamente quelle positive non possono uscire). Per espellere il Na+ si usa un’antiporter, che permette l'entrata del protone che cede energia utile a far uscire il Na+.
trasportare e le portano al trasportatore. In questo sistema è coinvolto un canale, la proteina idrolizzante ATP ed è coinvolta una proteina esterna. Molti batteri hanno uno spazio fuori dalla membrana in cui possono avvenire delle reazioni. Questo spazio è tipico di alcuni batteri che oltre alla membrana hanno una parete detta MEMBRANA PARIETALE , che deriva da un piccolo spazio detto “ spazio periplasmatico ” → esso funge da citoplasma esterno, in cui ci sono delle proteine che si stanno preparando al trasporto. Questa membrana esterna è presente solo in alcuni batteri detti Gram-negativi. Ha una composizione simile alla citoplasmatica ma fa parte della parete. Lo spazio periplasmatico ha un ruolo metabolico molto importante solo nei Gram negativi e questo perché questi hanno, oltre al peptidoglicano, ha un'altra membrana parietale e quindi fanno sì che il contenuto dello spazio periplasmetico non possa uscire. È un involucro che possiede delle funzioni in più funzioni, diverse da quelle della membrana plasmatica, soprattutto strutturali ma è completamente permeabile. Nel sistema ABC abbiamo il trasportatore transmembrana, abbiamo delle proteine che idrolizzano ATP e abbiamo delle proteine che non sono associate alla membrana che rimangono allo spazio periplasmatico. CATENA DI TRASPORTO DEGLI ELETTRONI : Nella membrana c'è un sistema che frutta la forza proton- motrice, ovvero l'ATP sintetasi. Questo è un canale →un trasportatore di protoni. I protoni entrano dissipando energia e questo complesso cede energia ad ADP che usa energia per formare ATP. La forza proton-motrice è sfruttata da ATPasi e trasporto semplice.
Membrana plasmatica negli Archea I lipidi di questa membrana hanno caratteristiche uniche che li distinguono dal mondo batterico e da quello eucariotico. Le differenze riguardano la chiralità del glicerolo , il tipo di legame tra glicerolo e catena alifatica e il tipo di catena alifatica. Infatti in questa categoria, il fosfato è esterificato con il glicerolo in C1, producendo così l’enantiomero L-glicerolo. Le catene laterali alifatiche non sono costituite da acidi grassi lineari insaturi esterificati al glicerolo, ma da catene isoprenoidi , il cui gruppo alcolico terminale è condensato all’L- glicerolo in C2 e C3 tramite legame etere ; le catene alifatiche sono costituite da isoprenoidi che formano lunghe catene sature ramificate. Inoltre i lipidi degli Archea possono essere dieteri (costituiti da due catene alifatiche a 20 atomi di C legate ad un’estremità a una molecola di glicerolo) o tetraeteri di glicerolo (hanno due catene alifatiche a 40 atomi di C e ciascuna estremità è legata a 2 molecole di glicerolo). I tetraeteri formano membrane monostrato che offrono una maggiore resistenza alla separazione rispetto al doppio strato; questo tipo di membrana lo troviamo negli Archea che vivono a temperature elevate.
- Funzioni della membrana plasmatica La membrana plasmatica è una barriera selettivamente permeabile tra il citoplasma e l’ambiente extracellulare. Il doppio strato permette la diffusione di acqua, gas e piccole molecole liposolubili; mentre risulta impermeabile a ioni, composti polari e grandi molecole. Queste molecole attraversano la membrana plasmatica grazie a specifici sistemi di trasporto, costituiti da proteine, che possono esseri sistemi di tipo passivo che avvengono secondo gradiente di concentrazione e di tipo attivo che avvengono contro gradiente. La membrana plasmatica è una delle sedi principali per le trasformazioni di energia in forme utilizzabili dalla cellula. Nei procarioti, a livello di questa struttura, avvengono respirazione e fotosintesi; in entrambi i processi viene generata la forza Proton-motrice. Questa è utilizzata dalla cellula per sintetizzare ATP e per alimentare altri processi. Grazie ai sistemi di trasduzione del segnale , le cellule sono in grado di rispondere rapidamente ai cambiamenti ambientali. Questi sistemi sono capaci di percepire gli stimoli esterni, trasmettere il segnale all’interno della cellula, modificare il profilo dell’espressione genica. I segnali possono essere di tipo fisico o fisico-chimico. Le proteine responsabili della trasduzione del segnale sono associate alla membrana e possiedono dei domini esposti all’ambiente extracellulare o domini citoplasmatici. La membrana plasmatica è sede di processi biosintetici : la sintesi di fosfolipidi e alcuni passaggi della sintesi del lipopolisaccaride e del peptidoglicano. Nel caso della biosintesi della membrana, la sintesi dei lipidi garantisce l’accrescimento della membrana durante la crescita cellulare. La parete batterica La maggior parte dei batteri, esternamente alla membrana plasmatica, possiede una parete. Sia la parete del Gram negativo che del Gram positivo possiedono un’unica enorme molecola polimerica detta SACCULO DI MUREINA, che avvolge completamente la cellula come una sorta di esoscheletro. Questo sacculo conferisce alla cellula rigidità e forma, ne protegge l’integrità contrastando la pressione osmotica ed è capace di restringere ed espandere il suo volume, fino a 3 volte senza rompersi. È sostanzialmente costituita da PEPTIDOGLICANO , che ha una struttura reticolare formata da catene glicaniche costituite da due amminozuccheri alternati, NAG (N-acetilglucosammina) e NAM (N- Il NAG e il NAM sono legati insieme dal legame 1,4-Beta-glicosidico
Biosintesi del peptidoglicano : è un processo complesso che possiamo suddividere in 3 stadi, ciascuno dei quali si svolge in un diverso comparto cellulare. Nel citoplasma avviene la sintesi dei precursori; a livello della membrana plasmatica si ha la sintesi dell’unità monomerica, il glicano pentapeptide, legato ad un trasportatore lipidico e abbiamo il suo ribaltamento sulla faccia esterna della membrana. Nello spazio esterno alla membrana avvengono le reazioni di polimerizzazione e transpeptidazione (ovvero la formazione dei legami crociati). PARETE DEI BATTERI GRAM POSITIVI (con il colore Gram diventa VIOLA) I legami crociati tra le subunità do glican tetrapeptide non sono diretti ma costituiti da un ponte peptidico, diverso tra le varie specie. La parete è una complessa struttura di macromolecole essenziali per la forma, l’integrità e il funzionamento della cellula. È costituita da un spesso strato di mureina in cui sono inserite altre molecole come gli acidi teicoici e acidi teicuronici , a cui si aggiungono proteine di superficie. Gli acidi teicoici si possono legare ai lipidi di membrana e costituiscono dal 30 al 60% della parete cellulare e sono ritenuti essenziali per il normale funzionamento della cellula. Gli acidi teicoici possono essere anche acidi di parete, che sono dei polimeri polianionici ricchi in fosfato e responsabili della carica negativa della superficie cellulare. Essi vanno a definire, insieme al peptidiglicano e a gli acidi teicuronici, le caratteristiche chimico-fisiche; controllano il movimento degli ioni e di altre molecole, antibiotici inclusi; regolano il movimento e l’attività di proteine periplasmatiche ed extracellulari. Lo spazio periplasmatico si trova tra la membrana plasmatica e la parete cellulare ed è più sottile rispetto a quello dei batteri Gram negativi. Gli acidi teicoici sono polisaccaridi acidi contenenti altri zuccheri e D-alanina. Possono legarsi ai lipidi della membrana, ancorando il peptidoglicano alla parete (ACIDI LIPOTEICOICI). Conferiscono stabilità alla parete, ma sono assenti dei Gram Negativi, perché in questo caso il peptidoglicano si trova in mezzo a due membrane. ↓
PARETE DEI BATTERI GRAM NEGATIVI (con il colore Gram diventa ROSA) Questa parete è molto più complessa della parete dei Gram positivi. Sono circondati da una membrana esterna, detta MEMBRANA PARIETALE , a doppio strato lipidico, ma diversa per composizione da le altre membrane biologiche. Oltre che da fosfolipidi e proteine infatti è costituita da complessi lipopolisaccaridici e quindi è anche detta STRATO LIPOPOLISACCARIDICO (LPS). Lo spazio tra la membrane esterna e quella interna è detto PERIPLASMA : qui si trova anche una parete mureinica costituita soltanto da un sottile strato di peptidoglicano. Questo spazio contiene molte proteine, dette proteine chaperon periplasmatiche. Il LPS è una grande e complessa molecola anfipatica contenente sia lipidi e saccaridi; vi si possono distinguere 3 diverse porzioni:
- Porzione lipidica, detta LIPIDE A
- Una regione polisaccaridica detta NOCCIOLO o CORE
- Una lunga catena polisaccaridica detta ANTIGENE O La membrana parietale ha dei fosfolipidi nella metà del foglietto che guarda verso l’interno, ovvero verso il peptidoglicano, mentre i lipidi che compongono lo strato più esterno sono detti lipo-polisaccaridi , in quanto hanno delle lunghe catene zuccherine. Questa ha un importante rilevanza biologica, infatti LPS (il foglietto esterno) è diverso tra batteri e batteri, in particolare la lunga catena polisaccaridica detta ANTIGENE O. In molti Gram negativi, che sono patogeni intestinali, quando entrano all’interno dell’organismo causano un danno immediato, sono tossiche perché interagiscono con le mucose in quanto sono endotossine. Queste sono la parte variabile dell’LPS, ed è il ruolo principale del foglietto esterno. La diffusione nel periplasma di ioni e piccole molecole è possibile grazie alla presenza di proteine canale, dette PORINE. Sono dei canali proteici, non specifici, che attraversano tutto lo spessore della membrana garantendo un canale acquoso per molecole di piccole dimensioni. Queste non sono dei trasportatori ma sono dei veri e propri pori, che non richiedono energia per l’attraversamento, perché sono dei varchi idrofilici che schermano il doppio strato fosfolipidico. In più sono presenti anche proteine transmembrana che sono fungono da trasportatori specifici.
- > (^) cambia (^) da (^) specie in (^) specie
strutture che non tutti i microrganismi possiedono, sono uno strato aggiuntivo che non interagisce con l’esterno; quello che interagisce è l’LPS. È considerato come un muco che avvolge la cellula. La presenza della capsula conferisce alle colonie batteriche una morfologia liscia e un aspetto lucido, mentre quelli privi di capsula formano colonie rugose. Gli involucri :
- CAPSULA : è uno strato zuccherino amorfo ma poco abbondante; è ben strutturata e adesa alla parete cellulare. Ha un ruolo fondamentale nei microrganismi patogeni, infatti i batteri dotati di capsula riescono a sfuggire al sistema immunitario dell’ospite (in particolare ai MACROFAGI, che hanno un ruolo di difesa in quanto riescono a fagocitare i batteri non voluti dall’organismo), chiamata anche fattore di virulenza. I macrofagi non riescono ad attaccare tutti i batteri perché la coevoluzione ha fatto sì che molti batteri diventassero positivi per l’organismo. I macrofagi riconoscono i batteri negativi tramite dei recettori presenti sulla superficie dei batteri, mentre quelli con la capsula hanno questi recettori coperti e quindi non li riconoscono: questi quindi diventano dei fattori di virulenza in quanto prolifereranno. Le capsule sono fortemente idratate (circa 90% acqua) e sono costituite da singole unità ripetute di monosaccaridi uniti da legami glicosidici.
- GLICOCALICE : è di natura zuccherina e molto più abbondante della capsula. A differenza di quest’ultima, il glicocalice ha un ruolo nell’adesione, ha la funzione di permettere ai batteri di aderire alle superfici e tenere insieme più batteri andando a formare dei “tappeti” sulle superfici. Favorisce la formazione di biofilm , che sono questi tappeti di cellule, che hanno un ruolo molto importante in campo ecologico e dell’infezione. Non dobbiamo confondere il LPS con il glicocalice, perché LPS è la parte zuccherina che però non ha funzione di adesione, come quella che invece possiede quest’involucro.
- STRATO S : sono involucri di natura proteica che sono simili a quelli degli Archea ma non lo sono. In comune gli Archea e batteri, che posseggono tale involucro, hanno le condizioni estreme in cui vivono: sono vere e proprie corazze della cellula. Questi strati para-cristallini sono molto ordinati a differenza di quelli zuccherini, che hanno strutture amorfe. Si forma dall’assemblaggio di subunità proteiche sulla superficie cellulare e costituisce un reticolo cristallino spesso, con simmetria esagonale, quadrata od obliqua. Questo rivestimento è ancorato, mediante legami non covalenti, alle differenti strutture della parete cellulare dei diversi gruppi microbici. È composto da proteine insolubili in acqua, debolmente acide, con alto contenuto di acido glutamico, acido aspartico, lisina e amminoacidi. Esse vengono secrete attraverso la via di secrezione generale dipendente da Sec. Una caratteristica di queste proteine è quella di essere legate, in modo covalente, a catene di glicani (costituiti da sequenze zuccherine che sporgono verso l’ambiente extracellulare). Nel caso della capsula ci sono alcuni batteri che la posseggono altri no, anche se appartengono alla stessa specie; è una piccola differenza di strutture accessorie. Le appendici esterne (tutte di natura proteica):
- FIMBRIE: sono corte e proteiche e ricoprono completamente la superficie, come dei peli, corte ma numerose. Non si muovono, ma i batteri contraendosi, contribuiscono al movimento di quest’ultimo, facendolo nuotare. Le fimbrie rappresentano un tatto dei batteri, che percepiscono delle superfici e toccandole riescono a riconoscerle, trasmettendo poi i segnali all’interno della cellula. Non sono loro ad attaccarsi ma giocano il ruolo di riconoscimento della superficie.
- PILI: sono strutture simili alle precedenti illustrate, sempre di natura proteica, ma sono più lunghi e di diametro maggiore, cavi al loro interno e sono presenti in un limitato numero di copie, solo in
alcune fasi della cellula batterica (di solito la cellula ne produce solo uno). Il pilo ha un ruolo particolare in un determinato momento della vita della cellula batterica, infatti vengono sintetizzati per permettere alla cellula di connettersi ad altre cellule. Essendo cavo permette alla cellula di collegare il citoplasma a quello di un'altra cellula: fa un ponte citoplasmatico, che parte da dentro la membrana. Permette lo scambio di contenuto citoplasmatico, di materiale genetico, tra cellule della stessa specie ma anche diverse. Esempio: fenomeno della coniugazione si basa sul pilo. La riproduzione asessuata dei batteri permette il rimescolamento genetico.
- FLAGELLI: sono spessi, lunghi e non sono cavi. Partano dalla membrana, attraversano la parete e si estendono verso l’esterno. Essi si muovono e ruotano come un’elica; è legato alla locomozione della cellula, attaccato ad un vero e proprio motore. Solitamente è uno, ma può essere anche più di uno. La struttura del flagello è ad uncino che attraversa la parete e che parte della membrana citoplasmatica attraverso una struttura ad anello; a ruotare in primis è questa, che di conseguenza fa ruotare l’uncino. Nei Gram positivi, essendoci solamente uno spesso strato di peptidoglicano, l’obbiettivo è quello che l’uncino attraversi la parete e che l’anello sia ancorato alla membrana; nei Gram negativi invece c’è una struttura più complessa di anelli che attraversano la parete formata da più strati. Il fatto che l’anello sia attaccato alla membrana ha un motivo fondamentale: va a sfruttare la forza proton-motrice, dato dalla proteina Mot che è un canale protonico. Questa trasferisce l’energia cinetica all’anello che viene trasmessa all’uncino e successivamente al flagello. DIFFERENZIAZIONE NEI BATTERI I batteri che hanno uno stesso genoma hanno una stessa morfologia. Questo però non vale per gli organismi superiori e pluricellulari. I batteri non si differenziano e la loro morfologia dipende dal genoma: se hanno morfologie e capacità diverse sicuramente avranno genomi diversi. I BATTERI NON SONO CAPACI DI DIFFERENZIARSI , ma esiste un’eccezione molto grande alla loro incapacità di differenziarsi. Infatti esistono dei batteri che hanno la capacità di cambiare drasticamente la loro morfologia, a parità di genoma. Questa è la capacità di trasformarsi in SPORA (no formazione ma trasformazione). Il modo più corretto di chiamare la spora è chiamarla ENDOSPORA BATTERICA: è una struttura che si forma all’interno del batterio che si sta trasformando.