Scarica Microbiologia - Parte 3 e più Sbobinature in PDF di Microbiologia solo su Docsity!
Lezione 16
Bacillus anthracis
Bacillo Gram positivo sporigeno aerobio; è facile da riconoscere al microscopio elettronico perché le cellula sembrano canne di bambù, perché sono di grandi dimensioni come bacilli e poi perché hanno la spora al centro della cellula quando viene generata che non deforma la cellula. Le colonie poi, osservate al microscopio su terreno solido, hanno un margine frastagliato e protusioni filamentose, chiamate “colonie caput medusa ” (vedi immagine su slide). B. anthracis è l’agente eziologico dell’ Antrace , detto anche Carbonchio, tipica patologia nozonosica che colpisce gli animali erbivori che spesso passa anche all’uomo (con contatto diretto tramite ferite o oggetti contaminati, oppure assumendo carni contaminate da spore o le pelli degli animali). Questa malattia viene anche chiamata appunto “Carbonchio” perché provoca delle ferite nere, color carbone. La causa è la liberazione all’interno dell’ospite di una tossina formata da 3 subunità che, a seconda della funzione della subunità della tossina, hanno nomi differenti:
- Antigene protettivo subunità/ parte della tossina che permette di legarsi in modo specifico ai recettori cellulari delle nostre cellule o di quelle degli animali.
- Fattore dell’edema l’esordio dell’Antrace è di tipo edemigeno, cioè di forma un edema e si produce soprattutto liquidi che vengono fuori alle cellule; quindi il fattore dell’edema provoca la fuoriuscita dei liquidi dalle nostre cellule.
- Fattore letale quando penetra all’interno della cellula questa muore, è una metallo-proteasi, tossica per le nostre cellule. Non funzionano separatamente: per poter penetrare deve prima legarsi, quindi deve essere contestualmente integra la tossina per poter essere efficace. Che manifestazioni ha il Carbonchio sull’uomo e sugli animali? Sono 3: dipende dalla via di penetrazione ˗ Carbonchio cutaneo : la forma cutanea è la più evidente, facile da diagnosticare ed è anche la più comune; tramite una ferita la spora batterica, essendo poi un bacillo aerobio, germina facilmente all’interno della ferita, germinando si libera il bacillo metabolicamente attivo che libera le tossine: le tossine hanno un’azione
necrotizzante (se è a livello cutaneo non è un grosso problema – si cicatrizza e si stacca la crosta – però può succedere che l’infezione si diffonda ai linfonodi e da lì passano al sangue, e quindi si muore rapidamente – è raro). ˗ Carbonchio da inalazione : tempo di incubazione molto lungo, forma molto rara; succede che si inalano delle spore batteriche, le spore di B. anthracis arrivano ai polmoni a livello alveolare e arriva il macrofago di turno che lo internalizza e rimane quiescente per un certo periodo (di solito dall’inalazione del batterio e dalla manifestazione della malattia possono passare anche 2 mesi); nel momento in cui le spore germinano all’interno del macrofago, i bacilli si liberano e incominciano a provocare danni estesi in maniera molto veloce (nel giro di poche ore) e il paziente muore nel giro di 24 ore, se va bene 48 ore. ˗ Carbonchio gastrointestinale : ancora più raro; se mi mangio una carne contaminata da spore succede che le spore resistono alla barriera gastrica, arrivano nella zona intestinale, germinano e la tossina fa lesioni ulcerative, necrosi a livello intestinale, e anche in questo caso si muore. Bacillus anthracis è facile da coltivare e questa facilità di coltivazione e di produzione di spore la resa purtroppo una facile arma biologica di distruzione di massa.
Bacillus cereus
Batterio Gram positivo sporigeno aerobio, tipico batterio β emolitico: Le sue tossine hanno un’azione fosfolipasica. B. cereus è un tipico agente eziologico di tossinfezione alimentare , è molto diffuso nell’ambiente soprattutto sotto forma di spore, ma è anche coinvolto in alcune infezioni ospedaliere: Infezioni oculari Endoftalmite dopo interventi chirurgici agli occhi (erano negli impianti di trattamento dell’aria e le spore sono finite dentro), germinata la spora il batterio produce tossine e distrugge il tessuto a livello degli occhi, si perde la vista. Infezioni sistemiche quando sono stati usati dei cateteri intravascolari contaminati sempre da spore di questo batterio; ancora più pericoloso perché il batterio riesce ad entrare nel circolo sanguigno. Come faccio ad accorgermi che ho mangiato un alimento contaminato? A seconda della tossina liberata, o meglio presente nell’alimento, ho 2 effetti tipici:
˗ Saprofiti funghi che crescono sul materiale organico in decomposizione, e favoriscono la decomposizione stessa. ˗ Simbionti funghi che collaborano con altri organismi per sopravvivere meglio: o Licheni = sono un’interazione simbiontica tra ascomiceti (che sono funghi) e le alghe. o Micorrize = funghi che interagiscono con le piante. o Ci sono funghi che crescono all’interno di esseri viventi, come gli insetti. ˗ Parassiti funghi patogeni (parassiti dei vegetali e anche parassiti degli uomini e degli animali)
FUNGHI PATOGENI
Le infezioni provocate dai funghi si chiamano micosi. Che tipo di micosi possono sussistere nell’ospite (uomo o animale che sia)? Superficiali: si hanno quando i funghi interagiscono con gli strati più esterni dell’epidermide, quindi non c’è una risposta/reazione immunitaria (dovrò utilizzare farmaci ad azione localizzata per eliminarli). Cutanee: i funghi interagiscono con la componente cheratinizzata dell’epidermide (unghie, capelli, peli e anche la cute stessa), in questo caso c’è una reazione immunitaria quindi c’è un’azione infiammatoria da parte dell’organismo (si cura con farmaci ad azione locale o con farmaci sistemici). Nell’ambito delle infezioni cutanee si distinguono due tipologie:
- Dermatofizie (dette anche Tigna o Tinea) es: Tinea pedis
- Dermatomicosi Sottocutanee: si formano i cosiddetti micetomi (micetoma vuol dire che il fungo è riuscito a penetrare attraverso una ferita, un taglio, un graffio) e comincia a diffondersi. Profonde: vengono chiamate anche infezioni sistemiche (es: aspergillosi polmonare: il soggetto inala i propagoli riproduttivi di un’ Aspergillus o di un altro fungo e può dar luogo ad un’infezione polmonare). Se un soggetto è immunocompetente di solito il fungo che penetra si ferma alle prime vie aeree, se però il soggetto è un immunodeficiente il fungo riesce ad arrivare nelle vie respiratorie inferiori e attacca il parenchima polmonare (può arrivare al torrente circolatorio). Quindi dipende da dove il fungo colonizza, se nella parte più esterna del nostro organismo oppure se via via si approfondisce nell’ospite. Ovviamente gli effetti che provoca una micosi sono differenti a seconda di dove colonizza.
Struttura dei funghi
Tipicamente si usa lo Stereomicroscopio per avere una visione tridimensionale del fungo (cioè i funghi microscopici filamentosi, non dei lieviti che sono ovviamente unicellulari). ˗ Tallo corpo del fungo: tallo unicellulare se parliamo dei lieviti, tallo pluricellulare se parliamo di muffe (cioè di funghi filamentosi). ˗ Ife un insieme di grosse cellule tubulari comunicanti tra loro, che danno luogo appunto ad una struttura che prende il nome di Ifa che cresce linearmente ma soprattutto in maniera ramificata e ha una particolarità: ha un allungamento di solito di tipo apicale. A livello fungino ci sono 3 tipi di Ife:
- Ife cenocitiche (sembra una cellula gigantesca con tanti nuclei, perché non ha setti, non hanno divisioni)
- Ife settate con cellule uninucleate
- Ife settate con cellule polinucleate Questi setti non sono integri, cioè non sono completamente separate, ma c’è un poro che permette la comunicazione e quindi il trasferimento di sostanze nutritive tra una cellula all’altra; quindi la punta assimila le sostanze nutritive che poi passano via via alle cellule sottostanti.
˗ Micelio quando le Ife si allungano e si intrecciano danno luogo al Micelio; il Micelio fungino ha 2 funzioni, a seconda di dove vanno le Ife: o Se le ife vanno verso l’alto costituiscono il Micelio aereo , la cui funzione tipica è quella riproduttiva, perché dal Micelio aereo si staccano i propagoli riproduttivi. o Se le ife vanno verso il basso allora costituiscono il Micelio vegetativo che sono le radici del fungo, traggono nutrimento dal substrato di colonizzazione. Nei funghi poi c’è un fenomeno particolare che si chiama “ dimorfismo cellulare ”, o meglio detto “ dimorfismo fungino ”: si cono funghi capaci di adottare la forma lievitiforme (cioè unicellulare) piuttosto che quella miceliale (cioè pluricellulare) a seconda dell’ambiente di crescita.
Processo di riproduzione dei funghi :
I funghi si riproducono attraverso la produzione dei cosiddetti propagoli riproduttivi (quelli che stanno nel Micelio aereo); i propagoli riproduttivi sono di 2 tipi, a seconda se ci sia una riproduzione di tipo:
1. Asessuato
Se parliamo di riproduzione asessuata allora si deve parlare di Conidi , ed è tipica dei funghi anamorfi (quelli che a livello medico vengono chiamati “funghi imperfetti”) e quindi si riproducono attraverso i conidi.
2. Sessuato
In questo caso si parla di spore sessuali o di meiospore ( funghi telomorfi , chiamati anche “funghi perfetti”). L’obiettivo finale è lo stesso: rigenerare un nuovo fungo, ma il metodo riproduttivo è differente (Meiosi da un parte e dall’altra la Mitosi).
MEMBRANA e CITOPLASMA A livello della membrana troviamo come componente irrobustente degli steroli , più precisamente Ergosterolo (diversi farmaci antimicotici hanno come bersaglio proprio l’ergosterolo). A livello citoplasmatico è una classica cellula eucariote, ricca di organuli, ma ha numero variabile di nuclei all’interno (si va da 1 a 30 circa, a seconda del tipo di ifa), i mitocondri hanno una forma simile a quella dei vermi, e cambia anche la quantità di inclusioni, cioè ci sono tantissime vescicole secretorie e di Vacuoli (che sono una riserva di acqua ma anche di oligoelementi che servono alla cellula fungina).
Differenza tra funghi e batteri
A livello ambientale sono più resistenti i funghi o i batteri? I funghi; perché se escludiamo la spora batterica, i funghi resistono di più all’ambiente perché appunto tollerano meglio i cambiamenti ambientali proprio per il loro corredo enzimatico molto ricco (possono stare in ambienti modo umidi come possono stare anche in ambienti aridi). Le muffe si adattano a stress più forti di quelli che possono sopportare la maggiori parte dei microrganismi, sono più resistenti dei batteri alle pressioni osmotiche e le muffe possono tollerare ed accrescersi in concentrazioni relativamente alte di acidi. Lezione 17
Lieviti
Sono dei miceti con tallo unicellulare. Saccharomyces boulardii viene usato come probiotico perché resiste alla barriera gastrica, quindi rimane intatto, e a livello intestinale può aiutare la flora batterica semplicemente producendo vitamina B o acido lattico che aiuta la flora microbica intestinale a contrastare patogeni obbligati che non sono presenti nell’organismo. Come appaiono i lieviti al microscopio? Sono cellule ovoidali che hanno un diametro vario che mediamente va dai 3 micrometri ai 15; se mi dovessi basare
solo sull’osservazione microscopica non riesco a identificarli perché risultano essere tutti uguali, stesso problema quando vedo la crescita a livello macroscopico, cioè su terreno di coltura specifico per i funghi (vedrei delle colonie color avorio, cremose, lucide, ma che sembrano tutte uguali). Per cui per identificarle devo ricorrere a strategie differenti: Utilizzo i sistemi API , che sono un insieme di test che valutano il profilo biochimico dei microrganismi che cerco di identificare (sono tante microprovette in cui inserisco il lievito che voglio andare ad indentificare e vado a vedere, a seconda del contenuto della provetta – zuccheri, amminoacidi ecc. –, cosa succede). In questo caso ho un’identificazione del profilo metabolico del fungo. Vado a visualizzare le mannoproteire , che sono componenti tipici della parete dei lieviti (che è composta da mannani, che sono zuccheri, e proteine di vario genere), quindi a seconda della sequenza amminoacidica e a seconda dell’organizzazione dei mannani posso dare un profilo.
Conidiogenesi blastica = Gemmazione:
La gemmazione è come viene chiamata la riproduzione dei lieviti. Ad un certo punto la cellula madre, la cellula conidiogena, inizia ad ingrossarsi e si formano delle escrescenze, delle vere e proprie gemme in cui andrà a migrare una copia del DNA del fungo; una volta che dentro c’è il genoma, la gemma si stacca (in lieviti attivamente metabolizzanti ci sono più escrescenze contemporaneamente). Ogni volta che una gemma si stacca si formano delle cicatrici, sia sulla “cellule madre”, cioè la cellula coidiogena, sia sulle cellule che ne derivano; per cui quando la superficie sarà tutta coperta di cicatrici il processo riproduttivo di quella cellula si blocca. I leviti possono dar luogo a diversi tipi di rapporti biologici con l’ambiente e con gli esseri viventi: tipicamente i lieviti sono saprofiti ; diversi sono commensali dell’uomo ; altri sono patogeni opportunisti ; poi ci sono dei lieviti che sono dei patogeni franchi ( Cryptococcus neoformans ).
Candida:
I funghi del genere Candida sono anche dei commensali, ma alcuni sono patogeni opportunisti ( riproduzione asessuata , quindi per gemmazione). Candida albicans è un commensale nel nostro organismo, però in alcuni casi può dar fastidio (infezioni). Altri fungi di questo genere che possono dar fastidio sono Candida glabrata e Candida tropicalis. Candida albicans ha 2 particolari caratteristiche quando è dentro l’ospite:
- È riconoscibile con il Test-tubulo germinativo se metto un campione contaminato da lieviti in un terreno di coltura liquido contenente siero di animale piuttosto che umano (a 37 °C per un t variabile dai 60 ai 90 min), se ho Candida albicans si forma il tallo unicellulare allungato, si forma una sorta di peduncolo (il cosiddetto “tubulo”), quindi c’è una protrusione della cellula ovoidale.
- A 37 °C a volte può dare una fase germinativa, cioè dalla cellula ovoidale partono delle sorte di ife che si intrecciano, quindi si forma una specie di micelio; questa fase germinativa provoca un danno alle mucose molto più esteso, ma soprattutto fanno sì che questi lieviti poi possono penetrare a livello ematico. CANDIDOSI SUPERFICIALI, dette anche CANDIDOSI LOCALIZZATE: Si sviluppano anche in soggetti immunocompetenti. Le candidosi localizzate vengono divise in 3 tipologie a seconda della localizzazione dell’infezione: ˗ Candidosi pseudomembranose è una candidosi che si sviluppa a livello del cavo orale, si forma una patina biancastra (che si chiama anche mughetto ) a livello della faringe, ma anche a livello della lingua. Succede soprattutto nei bambini piccoli e negli anziani, o anche soggetti con malattie croniche. ˗ Candidosi esofagee quando il mughetto si diffonde anche al tubo digerente. ˗ Candidosi VVC classica candidosi vulvo-vaginale; succede quando questo lievito incomincia a predominare sulla flora microbica vaginale.
Virus a RNA
Chiamati anche ribovirus.
Struttura del virus:
Un altro fattore a livello classificativo è la struttura.
Virus nudo Virus con involucro o rivestito
Ci sono virus nudi e virus rivestiti. Virus nudo , che è il virus più semplice strutturalmente è fatto: o Di genoma , la parte centrale (la parte centrale del virus viene anche chiamata core ); genoma con alcune proteine che hanno diverse funzioni che lo compattano. o Attorno al core c’è un involucro proteico che si chiama Capside , formato da tanti capsomeri (capsomeri che possono essere fatti da un tipo di protomero, cioè un tipo di proteine, o più proteine), quindi il Capside è una struttura proteica. Nel virus nudo il Capside è un fattore di protezione dall’ambiente (da agenti chimici o da fattori enzimatici) e in più permette l’interazione con le cellule ospiti. Il Virus rivestito riveste il virus nudo, cioè la struttura interna è identica, ho però un rivestimento (sempre di natura proteica); questo involucro viene chiamato Pericapside o Envelope (o anche Peplos ) e questo rivestimento lo derivano dalla cellula ospite (SarsCo2 è un virus rivestito).
Composizione chimica:
˗ Virus nudi acido nucleico e proteine. ˗ Virus rivestiti acido nucleico, proteine, lipidi e carboidrati. In entrambi, acido nucleico e proteine sono di codificazione virale ; lipidi e carboidrati derivano dalla cellula ospite.
Componenti virali:
Proteine strutturali , cioè proteine che fan parte della struttura del virus (quindi i capsomeri ad esempio, ma anche le spicole del Pericapside – sono delle antennine che protrudono, ad esempio in SarsCo2 è la proteina S, che permette l’interazione con la cellula ospite). Proteine funzionali , che sono proteine che il virus si porta dietro per conquistare il metabolismo della cellula ospite, quindi saranno polimerasi (che permettono la replicazione del genoma), piuttosto che proteasi (che bloccano il metabolismo cellulare per favorire quello virale – proteine che inducono lo “shift ribosomiale”, cioè inducono i ribosomi cellulari di smettere di lavorare per la cellula e iniziano a lavorare per il virus). Proteine di matrice (M) ; la matrice è chiamata anche “tegumento” (praticamente riguarda i virus rivestiti). Nei virus rivestiti, sopra al Capside, c’è uno spazio “cuscinetto” sempre costituito da proteine, dove si poggia il Pericapside e ha un ruolo con l’interazione con le cellule ospiti.
Non sempre posseggono un genoma costituito da DNA a doppia elica (può essere a singolo filamento oppure a RNA) Ci sono poi delle caratteristiche che sono solo peculiari dei virus: ˗ Acidi nucleici: nei virus ci sono o RNA o DNA, mai contemporaneamente; mentre nelle nostre cellule è il contrario, sono sempre insieme. ˗ Hanno una precisa specificità d’ospite ˗ Mentre batteri e funghi li possiamo far crescere in terreni abiotici, cioè sintetici, i virus hanno bisogno per forza di cellule per crescere. GENOMA DEI VIRUS Quello che si distingue poi, nell’ambito di due acidi nucleici, è anche la conformazione delle molecole, per cui avremo virus con:
- DNA ss
- DNA ds
- DNA RT il DNA viene trascritto in RNA e poi di nuovo viene sintetizzata una molecola di DNA.
- RNA RT vengono chiamati Retrovirus; per dar luogo all’informazione genetica della progenie virale, passerà attraverso un intermedio a DNA.
- RNA ss (+ e -)
- RNA ds “ss” è un acronimo in inglese che sta per “singola catena”. “ds” quindi sta per “doppia catena”. “RT” sta per Retro Trascrizionale, vuol dire che questi virus nel loro ciclo riproduttivo, per dar luogo a progenie virale, passano attraverso un intermedio a RNA o a DNA, a seconda se si tratta rispettivamente di DNA RT o RNA RT.
Ribovirus – virus a RNA
La maggior parte dei virus animali sono Ribovirus, cioè virus a RNA; i virus a RNA rappresentano un’anomalia assoluta a livello biologico, è un fatto unico, perché tutti gli organismi hanno l’informazione genetica nel DNA. Ad eccezione dei Retrovirus , che hanno un genoma diploide, tutti gli altri sono aploidi e poi l’RNA è variabile (può essere a singola o doppia catena, può essere lineare, circolare, ecc.), ma oltre alla conformazione, una caratteristica peculiare dei virus a RNA è che ci sono virus a RNA ss a polarità positiva o a polarità negativa. ~ POLARITÀ POSITIVA I virus a RNA ss hanno una sequenza nucleotidica che ha la stessa polarità del nostro mRNA cellulare; se hanno un genoma analogo al nostro RNA messaggero vengono chiamati “ virus immediatamente infettanti ” (come SarsCo2): l’RNA del virus viene scambiato dai ribosomi per RNA messaggero cellulare, ingannano i ribosomi che traducono immediatamente l’informazione genetica scritta in questo tipo di genoma. ~ POLARITÀ NEGATIVA In questo caso, il genoma non è immediatamente traducibile in proteine, quindi attraverso un enzima, una trascrittasi di tipo virale, verrà prima codificato un genoma positivo come polarità, che fungerà da messaggero, e poi si passa attraverso un antigenoma e poi si tornerà al mRNA negativo. I virus a RNA con polarità positiva o negativa hanno questi enzimi, queste trascrittasi, che sono in grado di trascrivere nuove coppie di genoma, il problema è che essendo degli enzimi virali spesso commettono degli errori, questo vuol dire che i virus a RNA sono più facilmente “capaci” di produrre errori e quindi di dar luogo a virus mutati cosa che non succede nei virus a DNA (cioè succede, ma con bassissima frequenza).
Deossiribovirus – virus a DNA
I virus a DNA sono tutti aploidi come informazione genetica e ci sono varie tipi di molecole: DNA bicatenario a molecola lineare
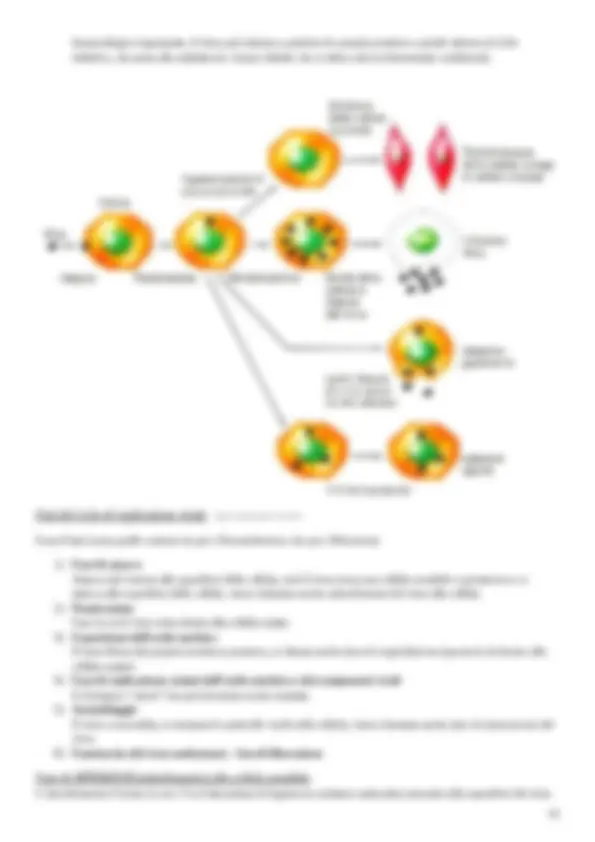
DNA bicatenario a molecola circolare DNA lineare formato da una sola catena nucleotidica ( Parvovirus ) Più è complesso il genoma di un virus più produce proteine: i Parvovirus sono piccoli e con un genoma piccolo, quindi produrranno meno proteine rispetto ad altri virus a DNA; i più complessi sono gli Herpesvirus e i Poxvirus che codificano per più di 100 proteine differenti. Perché i virus a DNA danno luogo a meno mutanti dei virus a RNA? Mentre i virus a RNA usano come trascrittasi degli enzimi che si portano dietro o che codificano loro, i virus a DNA nella maggior parte delle volte si replicano nel nucleo della cellula ospite e utilizzeranno le DNA polimerasi cellulari che hanno il cosiddetto “correttore di bozze”. Abbiamo detto che i virus sono dei parassiti endocellulari obbligati, quindi devono per forza penetrare all’interno della cellula ospite per riprodursi: si disassemblano ed espongono il loro acido nucleico in mood che venga trascritto e poi tradotto in varie proteine ecc. Quando questo succede? Succede quando i virus trovano cellule ospiti con 2 precise caratteristiche: le cellule ospiti devono essere SENSIBILI e PERMISSIVE. Sensibile vuol dire una cellula ospite che offra dei punti di attacco ai virus durante la penetrazione dell’ospite, cioè si dice tecnicamente che la cellula ospite abbia dei recettori a cui si possa legare il virus tramite i propri antirecettori virali, per permettere la prima fase del ciclo di replicazione virale (cioè l’attacco alla cellula ospite). Una cellula deve anche essere Permissiva perché, una volta attaccato, il virus entra nella cellula ospite e deve trovare quel complesso di fattori molecolari enzimatici che permettono di far esprimere il proprio genoma, quindi deve essere trascritto e poi sintetizzato di nuovo, e produrre soprattutto quei componenti virus-specifici, cioè proteine funzionali e strutturali del virus e non sempre accade, perché una cellula può essere sensibile ma non permissiva, oppure una cellula non è neanche sensibile, quindi non si può neanche attaccare. Dal punto di vista delle cellule ospiti, quali sono i tipi di infezioni che possono accadere? Che tipo di interazioni ha con la cellula ospite un virus, in base alla sensibilità e alla permissività della cellula ospite stessa? Ci sono diversi tipi di infezione: Infezioni di tipo litico il virus da luogo ad un’infezione citocida cioè che porta alla morte delle cellule ospite. Infezioni persistenti il virus trova una cellula sensibile e permissiva, però in questo caso non si ha una liberazione massiccia di virus, ma con più lentezza la cellula rilascia nel tempo le particelle virali, senza che però queste distruggano la cellula. Tipico nelle infezioni cronico virali. Infezioni trasformanti interazione dei virus con le cellule ospiti e le trasforma in cellule oncogene, perché durante il processo di replicazione che porta alla sintesi di nuove particelle virali, vengono prodotte delle proteine, ovviamente di codificazione virale, che interferiranno co il processo riproduttivo della cellula ospite, portandola ad una iperproduzione cellulare (le mortalizzano). Tutto questo succede quando i virus trovano cellule sensibili e permissive, ma ci sono delle volte che i virus trovano cellule sensibili ma non sempre totalmente permissive, e quindi si hanno: Infezioni abortive virus entra nella cellula, ma all’interno della cellula ospite non riesce a portare avanti il proprio ciclo replicativo fino in fondo e quindi non deriveranno particelle nuove virali, perché: o il virus si è danneggiato prima di entrare dentro la cellula e quindi non è in grado di esprimere completamente il proprio potenziale, oppure è la cellula stessa che legge una parte del suo genoma ma non integralmente e quindi non potrà portare alla sintesi di nuove particelle (può essere quindi un duplice ruolo, o del virus o della cellula ospite) quindi noi alla fine non percepiamo niente a livello infettivo. Infezioni restrittive il virus si replica sol quando la cellula ospite è in una determinata fase metabolica, quindi non sempre in questo caso quindi l’infezione la manifesta ma in certi periodi. Infezioni latenti come gli Herpesvirus: il virus trova una cellula sensibile e permissiva, entra dentro la cellula, ma il genoma non viene subito letto e trascritto e poi tradotto in proteine, ma si integra nel genoma della cellula e per un certo periodo, quando si duplica il genoma della cellula si duplica anche quel pezzettino del genoma virale che si è integrato. Solo in determinate condizioni, stress psicofisici piuttosto che una terapia
che vengono chiamati antirecettori virali (che sono le spicole) e molecole che fungono da recettori presenti sulla superficie della cellula ospite: il virus in pratica inganna la cellula perché si comporta come un “ligando fisiologico”. Questo tipo di legame tra antirecettore virale e recettore cellulare è un legame specifico ; questo legame è di 2 tipologie: ˗ Legame per interazione sterica, cioè per complementarietà strutturale, ed è tipico dei virus rivestiti; le strutture quindi si conformano bene, c’è una polarità strutturale con i recettori cellulari, quindi è un legame più saldo. ˗ Legame per complementarità di carica, e questo è un legame tipico dei virus nudi ed è un tipo di legame meno solido rispetto al precedente. L’adsorbimento può essere di più tipologie, si parla infatti di Adsorbimento: Reversibile vuol dire che il virus trova una cellula sensibile e quindi si lega, poi però non riesce ad andare alla fase successiva, cioè la Penetrazione, e quindi si deve staccare; viene definito “reversibile” perché questo distacco non comporta danni alla superficie e agli antirecettori virali che possono essere in grado quindi di infettare altre cellule e legarsi ad altre superfici. Irreversibile è il contrario: il virus si lega, ma il legame determina la modifica strutturare dell’antirecettore virale e quindi se il virus non dovesse penetrare non è più in grado di infettare altre cellule, perde il potere infettante (quindi in questo caso il virus, una volta avvenuto il legame tra antirecettore e recettore cellulare, o penetra o muore). Anticorpo mediato in pratica è dovuto ad errori dell’ospite, nel senso che la produzione di anticorpi per quel tipo di virus è troppo bassa, quindi si legano ma non sono in grado di neutralizzarli, quindi questi virus con attaccati degli anticorpi si legano a cellula dell’ospite che hanno recettori che hanno recettori per quel tipo di anticorpo e quindi ne approfitteranno per penetrare all’interno di quel tipo di cellule.
Fase di PENETRAZIONE :
Come può il virus entrare nella cellula? Lo può fare in 3 modi differenti a seconda del tipo del tipo di cellula che abbiamo davanti:
- Endocitosi , che è il meccanismo più comune di penetrazione e riguarda principalmente i virus nudi. In questo caso succede che il virus, dopo essersi attaccato/adsorbito sulla superficie della cellula ospite, al microscopio si noterà che il punto di attacco progressivamente si invagina e la membrana citoplasmatica tenderà a circondare il virus stesso, si forma una vescicola; perché la membrana si affossa? Perché l’adsorbimento, l’interazione con la cellula ospite, ha richiamato una proteina che si chiama clatrina che è quella che riveste le vescicole normalmente nelle cellule ospiti.
- Fusione , riguarda i virus rivestiti principalmente, perché il virus è ricoperto dall’Envelope (il Peplos) che altro non è che un pezzo della membrana cellulare (citoplasmatica o nucleare) quindi la fusione vuol dire proprio che il Pericapside si fonde con la membrana citoplasmatica della cellula ospite, in questo modo nel citoplasma entrerà solamente il virus con ancora il Capside attaccato.
- Traslocazione , riguarda principalmente i Picornavirus , tra cui il famoso Poliovirus : il Poliovirus è un virus nudo con simmetria del capside icosaedrica, quindi il virus si attacca/si adsorbe alla cellula ospite e l’antirecetttore virale è un componente del capside che si stacca, quando deve adsorbire alla cellula, e forma un poro attraverso al membrana di una cellula ospite, forma un canale; dal quel poro non entra tutto il virus, traslocherà solo il genoma del virus dentro al cellula (adsorbimento di tipo irreversibile).
Fase di SCAPSIDAZIONE :
Viene chiamata anche fase di uncoating: il virus dotato di capside, una volta entrato, nudo o rivestito che sia, si libera dell’involucro proteico, il capside si disgrega ed il genoma del virus entra nel citoplasma e questo avviene o in modo spontaneo oppure (più frequente) sono enzimi cellulari che pensano di avere a che fare con dei nutrienti e quindi vanno a disgregare il capside; a seconda che ci sia un genoma a DNA o a RNA avranno 2 destini differenti: La maggior parte dei virus a RNA stanno nel citoplasma Quelli a DNA vanno nel nucleo; alcuni virus a DNA non si scapsidano nel citoplasma perché essendo molto piccolini vanno direttamente nel nucleo e lo faranno lì (es: virus dell’epatite B). Quando c’è la scapsidazione inizia il “ periodo o fase di eclissi ”, vuol dire che quando il virus si è disgregato non lo vedo più e questo periodo dura per le fasi successive del ciclo di replicazione e finirà quando rivedrò di nuovo una forma virale che uscirà dalla cellula ospite.
Fase di REPLICAZIONE :
È una fase molto complicata e delicata, perché in questa fase il virus deve impadronirsi del metabolismo cellulare della cellula ospite. È un processo complicato e delicato perché il genoma virale deve competere con tutti i mRNA che dal nucleo arrivano ai ribosomi, quindi non è facile contrastarli; lo può fare in 2 modi:
- Grazie ad enzimi produce una quantità importante di mRNA però virali, che quindi possono competere.
- Impedisce con altri enzimi la fuoriuscita dell’RNA messaggero dal nucleo verso il citoplasma dove ci sono i ribosomi. Lo scopo quindi è contrastare i mRNA cellulari per far leggere e tradurre i propri. La modalità replicativa va distinta a seconda del genoma che ha il virus (a DNA oppure a RNA):
Deossiribovirus
I virus a DNA si possono suddividere in 4 gruppi replicativi. PRIMO GRUPPO: Adenoviridae
Virus a RNA ss con polarità negativa: In questo caso il genoma non può essere letto e tradotto immediatamente dai ribosomi cellulare e il problema è che non ci sono enzimi cellulari capaci di trasformare l’RNA negativo in positivo, quindi il virus lo porta dietro: ha un RNA polimerasi RNA-dipendenti e quindi trascriverà un RNA complementare a quello virale che ha polarità positiva e quindi poi a livello dei ribosomi viene tradotto e ovviamente tra queste ci sono le replicasi che sintetizzano delle coppie di RNA negativo. Virus a RNA ds: Hanno un filamento ha polarità positiva e uno ha polarità negativa; c’è il solito enzima che sul filamento a polarità negativa trascriverà un RNA messaggero a polarità positiva che va a livello ribosomiale e poi succederà tutto quello che succede per i RNA negativi. Retrovirus: Il più noto è il virus HIV ed è l’unico virus a RNA con patrimonio diploide, cioè ha due filamenti identici di RNA ss a polarità positiva, ma nessuno dei due si comporta da mRNA perché questi virus al loro interno hanno una trascrittasi inversa, cioè una DNA polimerasi RNA-dipendente che trascriverà una molecola di DNA partendo dal filamento di RNA (questo avviene a livello citoplamsatico). Questa molecola viene chiamata provirus : questo filamento di DNA deve andare a livello nucleare e lì incontrerà un altro enzima virale che si chiama integrasi che integra quel filamneto nel genoma dell’ospite; succede poi che o sta in latenza per un certo periodo, come tipicamente succede nei pazienti positivi al virus HIV che non manifestano subito l’AIDS, oppure inizia subito ad essere trascritto e tradotto in proteine strutturali e funzionali.
Fase di ASSEMBLAGGIO :
Hanno caratteristiche differente se parliamo di virus nudi o di virus rivestiti. Virus nudi : Il processo di assemblaggio è un processo spontaneo, cioè le componenti strutturali del virus si automontano per polarità strutturale, questo processo avviene nel distretto dove è avvenuta la replicazione del virus (citoplasma o nucleo). Il Capside si assembla e incorpora dentro il genoma (si è visto che la incorporazione del genoma rende stabile la struttura e la rende salda), però è un processo non sempre esente da errori; ha volte di montano Capsidi vuoti, cioè dento non c’è il genoma, oppure si formano Capsidi con dentro il genoma ma mancano dei capsomeri, in entrambi i casi il virus non è infettante. Nei virus nudi si formano tante particelle virali, sia a livello nucleare che citoplasmatico, e danno luogo agli “ aggregati paracristallini ”, cioè un accumulo a livello citoplasmatico o nucleare di particelle virali insieme a dei piccoli cristalli che poi fuoriescono di solito per lisi cellulare. Virus rivestiti : La prima fase è analoga a quella dei virus nudi, cioè il processo di montaggio, ma poi questi tipi di virus hanno bisogno del rivestimento e quindi poi, dopo l’assemblaggio, questi virus si avvicinano ad una membrana (che può essere quella citoplasmatica se si assemblano nel citoplasma, o nucleare se si assemblano nel nucleo) e si aggancia ad un pezzo di membrana che è stata precedentemente funzionalizzata con glicoproteine virale. A questo punto il virus
uscirà per “gemmazione”, cioè si aggancia alle glicoproteine della membrana e si porta via un pezzo di membrana per rivestirsi. Spesso i virus rivestiti che gemmano non danno luogo a lisi cellulare. Lezione 19
I Fagi
I fagi batterici, o batteriofagi, sono virus che parassitano le cellule batteriche (non possono parassitare l’uomo perché c’è una forte specificità d’ospite che gli impedisce di entrare nelle nostre cellule). Struttura : È una struttura complessa, si dice a “ simmetria binaria ” cioè è fatta da una testa icosaedrica che contiene il genoma virale (i fagi tipicamente sono a DNA, rarissimi casi con genoma a RNA), sotto la testa icosaedrica c’è una struttura che viene chiamata coda, che è invece a simmetria elicoidale, che è quella che permetterà il passaggio del genoma nella cellula batterica quando questo si attaccherà ad essa; la coda termina con una struttura a punta che si chiama piastra basale , che permette proprio il passaggio dentro alla cellula batterica, che si sfrangia in 5 o 6 fibre caudali (sempre di natura proteica), queste fibre sono tipicamente gli antirecettori virali dei batteriofagi. I fagi sono gli unici virus che, oltre a poterli vedere al microscopio elettronico, posso vederli anche utilizzando un terreno per batteri, un terreno agarizzato, proprio perché interagiscono con i batteri: per visualizzarli prendo una piastra ci verso sopra una grande quantità di batteri, aggiungo il campione (di solito acqua) che sospetto sia contaminata da quei virus e poi metterò in incubazione, poi vedrò le colonie (se in quel campione sono contenuti i fagi) e sulla piastra vedrò delle zone di lisi circolare ( vedi immagine ), queste zone vengono chiamate placche e sono il prodotto di interazione del parassitismo del fago nei batteri: dove sono penetrati i fagi hanno ucciso i batteri e quindi le colonie non si sono generate e quindi rimangono dei piccoli buchi su tutta la piastra; in un terreno liquido questa cosa non si può fare. L’unità di misura è la PFU = unità formanti placche, ogni buco corrisponde ad ogni singolo fago. MOLTIPLICAZIONE:
- Adsorbimento
- Iniezione dell’acido nucleico
- Fase replicativa precoce
- Replicazione del genoma virale
- Sintesi delle proteine capsidiche
- Assemblaggio del capside e impacchettamento del genoma virale
- Rilascio dei virus maturi (lisi) Adsorbimento: Abbiamo l’interazione con gli antirecettori virali, che in questo caso sono le fibre caudali, e strutture dei batteri che saranno differenti a seconda del tipo di parete che possiede il batterio (se Gram positivo o negativo per esempio): tipicamente nei Gram positivi le fibre caudali si legheranno agli acidi teicoici, ma possono anche attaccarsi all’unica appendice cellulare, cioè il flagello; nei Gram negativi si possono agganciare alle appendici cellulari, come flagelli o i pili sessuali, oppure si attaccano al lipopolisaccaride. Penetrazione : Una volta che il fago si è attaccato si comporta in modo diverso dalla maggior parte dei virus animali, cioè che non entra tutto il virus, ma inietta solo l’acido nucleico: la piastra basale si poggia sulla superficie del batterio (lo buca) e grazie alla coda che si contrae (come se fosse uno stantuffo di una siringa) il genoma va dalla testa alla cellula batterica.