Scarica virus e batteri, . e più Appunti in PDF di Chimica solo su Docsity!
I virus
I virus: caratteristiche generali
I virus sono agenti infettivi con le seguenti caratteristiche distintive:
- dimensioni estremamente piccole (media diametro tra 20 e 800 nm)
- parassitismo intracellulare obbligato
- organizzazione strutturale semplice non cellulare
- presenza di un solo tipo di acido nucleico (DNA o RNA)
- si moltiplicano per sintesi separata e successiva riunione dei loro componenti Spettro d’ospite. I virus si moltiplicano solo in particolari cellule ospiti e di conseguenza vengono suddivisi in tre classi principali:
- virus batterici (o batteriofagi)
- virus animali
- virus vegetali
Origine dei virus
Evoluzione retrograda I virus sarebbero forme degenerate di parassiti intracellulari: i loro genomi si sarebbero progressivamente ridotti in quanto la maggior parte delle funzioni vengono fornite dalla cellula ospite. Questa ipotesi potrebbe spiegare l’origine di virus complessi quali i Poxvirus
- Teoria della “evasione dei geni” I virus si sarebbero evoluti da elementi genetici capaci di replicazione autonoma (plasmidi e trasposoni)
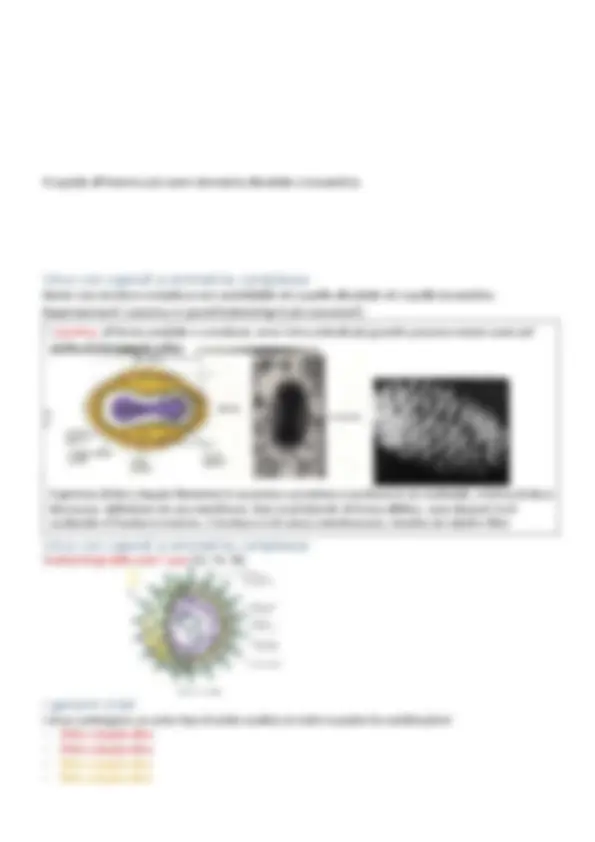
Struttura dei virus
- Le particelle virali, o virioni, sono costituite da una o più molecole di DNA o RNA rivestite da un involucro proteico o capside.
- In alcuni virus il nucleocapside (acido nucleico + capside) può essere avvolto da un rivestimento membranoso (envelope)
- Le dimensioni possono variare da 10 a 400 nm
- In base alle caratteristiche morfologiche è possibile individuare 4 gruppi principali: Virioni con capside icosaedrico Virioni con capside elicoidale Virioni dotati di envelope Virioni complessi Sia i capsidi elicoidali che icosaedrici sono strutture macromolecolari costruite da una o poche subunità proteiche, dette capsomeri, ripetute un gran numero di volte. Questa strategia costruttiva ha il vantaggio di utilizzare con la massima efficienza l’informazione immagazzinata nel genoma virale
Capsidi a simmetria elicoidale
Di forma cilindrica, costituiti da un unico tipo di capsomero avvolto a spirale attorno all’asse centrale contenente l’acido nucleico I capsidi possono essere rigidi (virus del mosaico del tabacco e il batteriofago M13) o flessibili (virus influenzali) Esempio: virus del mosaico del tabacco (TMV) Capside cilindrico rigido di 15-18 nm di diametro e circa 300 nm di lunghezza. L’unica proteina del capsomero è di 158 aa (amminoacidi) Acido nucleico: RNA a singolo filamento di 6000 nucleotidi Cristallizzato da Stanley nel 1935 Spesso la ripetizione di una sola proteina rappresenta l’unica possibilità di rivestire un acido nucleico di piccole dimensioni
Capsidi a simmetria icosaedrica
Icosaedro: solido regolare a 20 facce triangolari e 12 vertici I capsomeri sono formati da 5 o 6 protomeri che possono essere anche tutti identici I pentoni o pentameri possiedono 5 subunità e sono disposti ai vertici dell’icosaedro; gli esoni o esameri hanno 6 subunità e costituiscono le facce Esempi: i batteriofagi ΦX174, MS2, QβX174, MS2, Qβ parvovirus (12 capsomeri), poliovirus (32 capsomeri), papilloma virus (72 capsomeri), adenovirus (252)
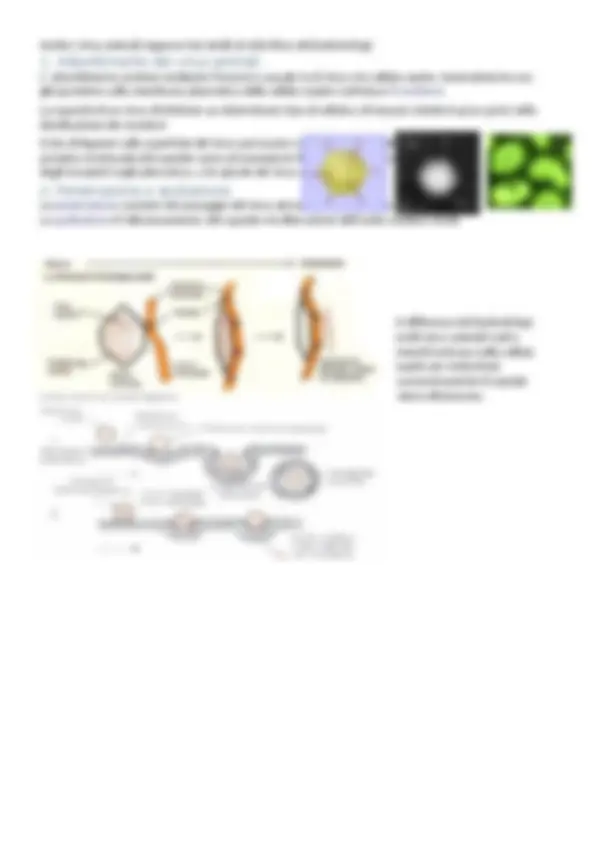
Virus dotati di envelope
Presentano il nucleocapside rivestito da strutture membranarie complesse (involucro pericapsidico) I virus con involucro sono comuni nel mondo animale, ma si conoscono anche virus batterici L’involucro pericapsidico è generalmente costituito da un doppio strato lipidico in cui sono inserite glicoproteine (spicole) La membrana deriva dalla cellula infettata in seguito al processo di liberazione dei virioni; le glicoproteine sono di origine virale 1 A = 0,1 nm
Anche la struttura del genoma è varia nei diversi virus. Il materiale genetico può essere organizzato in:
- un’unica molecola lineare
- un’unica molecola circolare (DNA)
- segmentato in più molecole
I genomi virali
La capacità codificante va dalle 3-4 proteine dei virus più piccoli (MS2 e Qb) alle oltre 100 proteine dei batteriofagi della serie T-pari e dei Poxvirus Nella maggior parte dei virus a DNA il genoma è costituito da un doppio filamento (ds DNA) che può essere lineare oppure circolare
La formazione dell’RNA messaggero virale
Affinché possano essere sintetizzate le nuove proteine virali, è prima necessario che siano sintetizzati nuovi mRNA (messaggeri) virali Virus a DNA
I genomi virali
Per la maggior parte dei virus a RNA il materiale genetico è rappresentato da un singolo filamento di RNA (ss RNA) Se la sequenza corrisponde a quella dell’mRNA virale il filamento è definito positivo o + Esempi: il virus polio, il virus del mosaico del tabacco, e il virus del sarcoma di Rous sono virus con genoma a ssRNA positivo Se la sequenza è complementare a quella dell’mRNA virale il filamento è definito negativo o - Esempi: il virus della rabbia, il virus del morbillo, e il virus dell’influenza sono virus con genoma a ssRNA negativo Esistono virus a RNA che possiedono un doppio filamento di RNA.
Coltivazione dei virus
I virus animali vengono coltivati inoculando animali sensibili o uova embrionate di pollo (uova fecondate e incubate per 6-8 giorni) Di recente i virus vengono coltivati su monostrati di cellule animali; questa tecnica mette in evidenza aree localizzate di lisi cellulare chiamate placche, o foci proliferativi (virus oncogeni) I virus vegetali possono essere coltivati impiegando colture di tessuti vegetali, oppure piante intere. Le foglie si inoculano meccanicamente spalmandole con una miscela di virus e una sostanza abrasiva. Nelle aree infette si sviluppa una lesione necrotica localizzata
I batteriofagi sono i virus più facilmente coltivabili. L’infezione di una coltura di batteri sensibili porta nel giro di circa un’ora alla lisi dei batteri con produzione di una notevole quantità di virus (lisato)
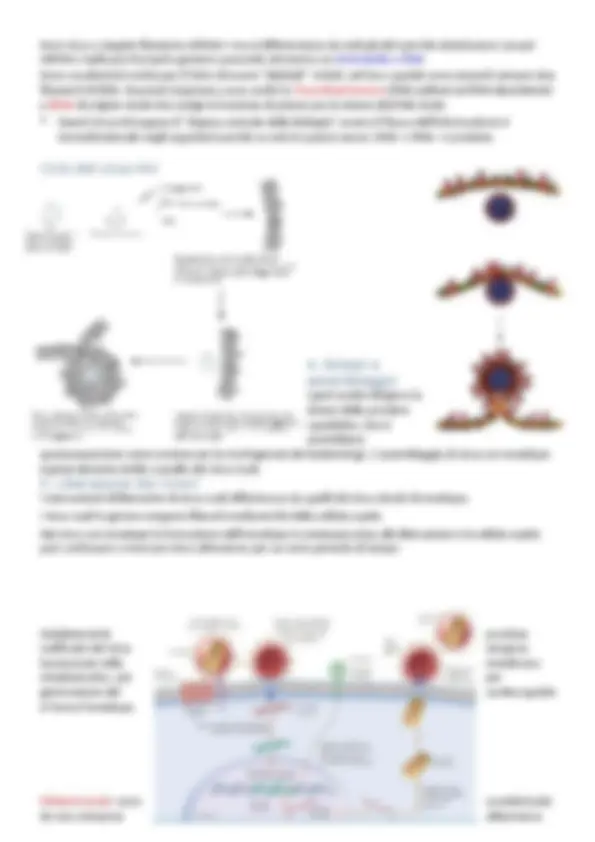
Le fasi della moltiplicazione dei batteriofagi
- Adsorbimento
- Iniezione dell’acido nucleico
- Fase replicativa precoce
- Replicazione del genoma virale
- Sintesi delle proteine capsidiche
- Assemblaggio del capside e impacchettamento del genoma virale
- Rilascio dei virus maturi (lisi) Queste sono le fasi generali della moltiplicazione dei virus che infettano i batteri; il processo prende il nome di CICLO LITICO, la cellula batterica alla fine muore essendo lisata alla fuoriuscita dei fagi.
L’adsorbimento dei batteriofagi
I batteriofagi non aderiscono a un punto qualunque della superficie batterica ma riconoscono recettori specifici I recettori sono normali componenti della superficie dell’ospite quali proteine della parete, polisaccaridi, lipopolisaccaridi, acidi teicoici, flagelli e pili In assenza di siti recettoriali il virus non può adsorbirsi Se un recettore è alterato, l’ospite diventa resistente alla infezione da parte del virus che usa quel recettore. Tuttavia, anche i virus possono mutare per la struttura che riconosce il recettore tornando, così, capaci di infettare un ospite resistente Fago PBSI di B. subtilis
Penetrazione dell’acido nucleico
Nella gran maggioranza dei batteriofagi è solo l’acido nucleico virale a entrare nel batterio. I meccanismi di penetrazione differiscono notevolmente tra i diversi fagi finora studiati e sono in gran parte ancora oscuri. E. coli infettato da T
La via lisogenica
Il cromosoma di λ si integra nel cromosoma batterico. Una volta integrato un solo gene di λ è responsabile del mantenimento dello stato lisogenico: il gene cI
Virus animali
Possono essere nudi o rivestiti da envelope, gli ospiti sono le cellule eucariotiche degli animali Per la classificazione vengono prese in considerazione le caratteristiche morfologiche e la composizione in acido nucleico
Anche i virus animali seguono fasi simili al ciclo litico dei batteriofagi.
1. Adsorbimento dei virus animali
L’ adsorbimento avviene mediante l’incontro casuale tra il virus e la cellula ospite. Generalmente una glicoproteina sulla membrana plasmatica della cellula ospite costituisce il recettore La capacità di un virus di infettare un determinato tipo di cellula o di tessuto risiede in gran parte nella distribuzione dei recettori Il sito di legame sulla superficie del virus può essere rappresentato semplicemente da una proteina strutturale del capside come ad esempio le fibre che si estendono dai vertici degli icosaedri negli adenovirus, o le spicole dei virus dotati di envelope
2. Penetrazione e spoliazione
La penetrazione consiste nel passaggio del virus attraverso la membrana plasmatica. La spoliazione è l’allontanamento del capside e la liberazione dell’acido nucleico virale. A differenza dei batteriofagi, molti virus animali nudi e rivestiti entrano nella cellula ospite per endocitosi; successivamente il capside viene allontanato.
Sono virus a singolo filamento di RNA + ma si differenziano da tutti gli altri perché sintetizzano i propri mRNA e replicano il proprio genoma passando attraverso un intermedio a DNA Sono caratteristici anche per il fatto di essere “diploidi”. Infatti, nel loro capside sono presenti sempre due filamenti di RNA. Associati al genoma sono anche la Trascrittasi inversa (DNA polimerasi RNA dipendente) e tRNA di origine virale che svolge la funzione di primer per la sintesi del DNA virale
- Questi virus infrangono il “dogma centrale della biologia” ovvero il flusso dell’informazione è monodirezionale negli organismi perchè va solo in questo senso: DNA → RNA → proteine.
Ciclo del virus HIV
4. Sintesi e
assemblaggio
I geni tardivi dirigono la sintesi delle proteine capsidiche, che si assemblano spontaneamente come avviene per la morfogenesi dei batteriofagi. L’assemblaggio di virus con envelope è generalmente simile a quello dei virus nudi.
5. Liberazione dei virioni
I meccanismi di liberazine di virus nudi differiscono da quelli dei virus dotati di envelope. I virus nudi in genere vengono liberati mediante lisi della cellula ospite. Nei virus con envelope la formazione dell’envelope è contemporanea alla liberazione e la cellula ospite può continuare a riversare virus all’esterno per un certo periodo di tempo Inizialmente le proteine codificate dal virus vengono incorporate nella membrana citoplasmatica, poi per gemmazione del nucleocapside si forma l’envelope. Infezioni acute: sono caratterizzate da una comparsa abbastanza
rapida e da una durata ridotta; le cellule infettate generalmente vanno incontro a morte (effetto citopatico) e vengono liberati numerosi virioni. Le infezioni acute sono tipiche dei picornavirus , herpesvirus e adenovirus. Effetti citopatici
- inibizione della sintesi di DNA, RNA e proteine
- danneggiamento dei lisosomi con liberazione di enzimi idrolitici
- alterazione della membrana plasmatica per inserimento delle proteine virali (attacco del sistema immunitario)
- alta concentrazione delle proteine virali
- rotture cromosomiche Infezioni persistenti o croniche: possono durare anche molti anni. I virus possono replicarsi più lentamente senza causare una sintomatologia clinica. Sono presenti anticorpi diretti contro il virus infettante. Il virus dell’epatite B, il virus di Epstein-Barr e il virus del morbillo possono dare infezioni persistenti. Infezioni latenti: dovute a virus che, dopo una iniziale fase moltiplicativa, arrestano il proprio ciclo di sviluppo e diventano quiescenti. La fase di latenza può durare un certo periodo dopo il quale può verificarsi una recrudescenza dell’infezione con una manifestazione spesso diversa da quella iniziale. I virus erpetici possono dare infezioni latenti. L’ Herpes simplex di tipo 1 infetta in genere i bambini e rimane quiescente per anni a livello dei gangli del sistema nervoso; periodicamente può attivarsi e dare le caratteristiche lesioni erpetiche (herpes labiale) Il virus della varicella zoster (Herpes zoster) provoca la varicella come prima manifestazione; non viene eliminato dal sistema immunitario ma diventa quiescente nei gangli dei nervi spinali. La manifestazione ricorrente è il “fuoco di Sant’Antonio” per la riattivazione del virus a livello dei nervi intercostali