


OSNOVI GENETIKE
skripta za studente
Poljoprivrednog fakulteta u Kruševcu
docent dr Vesna Perišić
2019.





































































Studirajte zahvaljujući brojnim resursima koji su dostupni na Docsity-u

Zaradite bodove pomažući drugim studentima ili ih kupite uz Premium plan

Pripremite ispite
Studirajte zahvaljujući brojnim resursima koji su dostupni na Docsity-u
Nabavite poene za preuzimanje
Zaradite bodove pomažući drugim studentima ili ih kupite uz Premium plan
ova skripta napisana je u želji da vam približim problematiku koju ... Molekularna genetika proučava molekulsku strukturu gena i njihovo funkcionisanje u.
Tipologija: Rezime
1 / 77

Ova stranica nije vidljiva u pregledu
Ne propustite važne delove!
Razvoj genetike kao naučne discipline može smestiti u prethodnih 100 godina. Ljudi
su, doduše nesvesno, primenjivali genetičke principe od najranijih ljudskih civilizacija. Kao
dokaz ove tvrdnje može se uzeti proces pripitomljavanja biljaka i životinja, započet pre
10.000 - 12000 godina u oblasti Srednjeg istoka. Prvi odomaćeni organizmi bili su pšenica,
grašak, sočivo, ječam, pas, koza i ovca. U drevnoj Asiriji i Vavilonu, pre 4.000 godina, radom čoveka je stvoreno nekoliko stotina sorti urmi, različitih po vremenu sazrevanja,
veličini i boji ploda, ukusu i dr.
Stari Grci su veliku pažnju poklanjali reprodukciji i naslednosti kod čoveka. Grčki
filozofi su razvili koncept pangeneze , po kojem su posebne čestice (gemule) bile nosioci
informacija iz različitih delova tela u reproduktivne organe iz kojih su se prenosile na
embrion u momentu stvaranja. Iako naučno netačan, ovaj koncept se održao sve do početka
Može se smatrati da je genetika, kao biološka nauka koja se bavi proučavanjem
nasleđa promenljivosti i uzroka ontogenetskog razvića, u savremenom smislu značenja,
započeta serijom principa i analitičkih procedura 1860. godine.
Najznačajniji momenti u naučnim otkrićima koji su doveli do nastanka genetike su:
Hooke je otkrio ćeliju (1665).
Grew je izneo dokaze o polnom razmnožavanju biljaka (1676), putem muških i ženskih
polnih ćelija. Mendel je utvrdio osnovne principe nasleđivanja (1866) i smatra se osnivačem (ocem)
genetike.
Brown je opisao ćelijsko jedro (1833).
Schleiden i Schwann su predložili osnove teorije o ćeliji (1839), po kojoj je sav život
sastavljen od ćelija; ćelije nastaju samo iz prethodno postojećih ćelija i ćelija predstavlja
osnovnu jedinicu strukture i funkcije u živim organizmima.
Miescher (1869) utvrdio prisustvo dezoksiribonukleinske kiseline (DNK) u jedru ćelije. Flemming je prvi posmatrao deobu hromozoma i na osnovu toga dao detaljan opis mitoze
(1879), pri čemu je pretpostavio da su u jedru smeštene nasledne informacije.
Darwin je u publikaciji O poreklu vrsta (1859) objavio teoriju o evoluciji kroz prirodno
odabiranje, čime je negirano božansko poreklo živih bića (kreatizam).
Sutton i Boveri su utvrdili da su geni, kao jedinice nasleđivanja, smešteni na hromozomima
(1902).
Morgan je otkrio prve genetičke mutante voćne mušice - Drosophila melanogaster (1910) i
razjasnio mnoge nepoznanice u prenošenju naslednih informacija. Fisher , Haldane i Wright su, u prvoj polovini 20. veka, postavili osnove populacione
genetike povezujući klasičnu (Mendelovu) genetiku i teoriju evolucije.
Hershy i Chase (1952) dokazali da je DNK prenosilac genetičke informacije.
Watson i Crick , uporedo sa Wilkins i Franklin , opisali su trodimenzionalnu strukturu DNK
(1953), započinjući eru molekularne genetike.
Izvedeni prvi eksperimenti sa rekombinantnom DNK (1973). Gilbert i Sanger su razvili prve metode za sekvencioniranje DNK (1977). Mullis (1944) je započeo rad na razvoju lančane reakcije polimeraze (PCR - polymerase chain reaction ), metode za brzo umnožavanje malih količina dostupne DNK. Očitan prvi genom živog organizma - bakterije Haemophilus influenzae (1995), metodom sekvencioniranja, a godinu dana kasnije i genom eukariotskog organizma (kvasac). Objavljen prvi, nepotpuni prikaz ljudskog genoma (2000), pri čemu je potpuno sekvencioniranje završeno 2003. godine. Pojednostavljeno rečeno, genetika je nauka koja se bavi proučavanjem prirode strukture, funkcije, kombinacija i prenošenja gena kod organizama.
U toku intenzivnog razvitka od oko osamdeset godina, genetika se razgranala na veći broj naučnih disciplina. Definisanje ovih disciplina mora se izvršiti na dosledan način, što ne znači da oblast koju zahvata jedna od tih disciplina ne zalazi i u druge koje se bave nešto drukčijom problematikom. Najpogodnije je podelu na genetičke discipline izvršiti na osnovu nivoa organizacije žive materije koja se proučava.
- Molekularna genetika proučava molekulsku strukturu gena i njihovo funkcionisanje u zavisnosti od te strukture. Ispitivanje funkcionisanja gena se u ovom slučaju ograničava na utvrđivanje procesa nastanka njihovih primarnih produkata (tj. jedne od RNK) i na procese prenošenja genetičke informacije preko tih primarnih produkata na stvaranje odgovarajuće strukture molekula proteina, koji predstavlja osnovni rezultat aktivnosti svakog gena. Pojam biohemijska genetika upotrebljava se često kao sinonim pojmu molekularna genetika, mada sa nešto širim značenjem. - Citogenetika izučava mehanizme promenljivosti i nasleđivanja osobina kroz analizu mikroskopske građe i funkcije pojedinih organela ćelije. Posebno intenzivna istraživanja vrše se na hromozomima, kao osnovnim nosiocima naslednih faktora. - Genetika razvića (ili fiziološka genetika) proučava na koji se način kod jedinki – pripadnika pojedinih vrsta ostvaruje razviće određenih osobina, i kako dolazi do toga da delovi tela sa istom genetičkom konstitucijom sasvim različito mogu da izgledaju, tj. „diferenciraju“ se u toku razvića. - Populaciona genetika izučava odnose u genetičkoj strukturi pojedinih grupa organizama, utvrđujući razlike i sličnosti među populacijama iste vrste, kao i uzroke i poreklo nastanka evolucionih adaptacija kod organizama.
U okviru svake od navedenih disciplina naučnici se mogu baviti nizom specifičnih problema. Na primer, u okviru molekularne genetike postoji genetika nukleinskih kiselina ili genetika proteina ili se istraživači bave procesima koji se na molekularnom nivou dešavaju u jednoj od organela (genetika ribozoma, jedarceta-nukleolusa i sl.). U okviru citogenetike, posebno mesto pripada izučavanju promena koja se dešavaju u strukturi hromozoma (mutaciona genetika), mada se ove promene izučavaju i na molekularnom nivou. U okviru
kontrolišu ove reakcije značajan, posebno proučavanje alela koji kontrolišu defektnu biohemijsku reakciju.
4. Mikroskopska analiza. Citogenetička istraživanja strukture hromozoma i hromozomskih aberacija korišćenjem svetlosne, elektronske i fluorescentne mikroskopije u identifikaciji lokacije gena na hromozomima i produkata njihove ekspresije u jedru i citoplazmi. 5. Tehnologija rekombinantne DNK. U posebnom poglavlju su prikazane mogućnosti koje pruža ova tehnologija u genetičkim eksperimentima.
Ćelija predstavlja osnovnu gradivnu i funkcionalnu jedinicu živog organizma, u kojoj su sadržane sve neophodne informacije za nastanak i razvoj novog organizma. Citologija je nauka koja se bavi proučavanjem ćelije. Svaka ćelija ima karakterističan oblik i veličinu. Organizmi mogu biti jednoćelijski i višećelijski. Osnovna podela živih organizama, prema stepenu složenosti građe, izvedena je na dve glavne grupe:
Prokariote su jednoćelijski organizmi i odlikuju se relativno jednostavnom strukturom ćelije (prosta ćelijska građa sa malim brojem prostijih organela). Ovi organizmi nemaju nukleus, obim ćelije je relativno mali (od 1 do 10 μm), nemaju membrane koje okružuju organele. Genom prokariota čini cirkularni (kružni) molekul DNK bez proteinskog omotača.
Eukariote unutar protoplazme poseduju ćelijske organele sa diferenciranim membranama (endoplazmatični retikulum, ribozomi, plastidi, Goldžijev kompleks, lizozomi, centrozomi i vakuole), značajne za normalno funkcionisanje ćelije. Sa stanovišta genetike, glavna razlika u odnosu na prokariote je što eukariotske ćelije imaju jasno diferencirano jedro sa jedarcetom. Membrana jedra jasno okružuje genetički materijal (DNK) i odvaja ga od drugih gradivnih elemenata ćelije.
Druga značajna razlika između prokariota i eukariota zasniva se u načinu pakovanja (smeštanja) DNK u jedru. U prokariotskim ćelijama, geni su smešteni, najčešće, na pojedinačnom molekulu DNK, koji predstavlja hromozom. Odstupanje od ovog pravila predstavljaju neke bakterije, koje imaju više od jednog hromozoma, kao i postojanje posebnih molekula DNK – plazmida, koji su nosioci važnih bakterijskih gena. U eukariotskim ćelijama, geni su razmešteni na većem broju, često pravolinijskih hromozoma DNK molekula. Manji broj eukariota poseduje u određenim organelama kružne hromozome DNK molekule sa manjim brojem gena na njima. Virusi ne pripadaju ni prokariotima, niti eukariotima, s obzirom da nemaju ćelijsku strukturu. Poseduju spoljašnji proteinski omotač koji okružuje nukleinsku kiselinu (DNK ili RNK). Mogu se razmnožavati samo u ćelijama domaćina, što ukazuje na činjenicu da su prvi virusi nastali tek nakon razvoja (evolucije) prve ćelijske strukture. Takođe, među virusima je prisutna i određena diferencijacija s obzirom da geni biljnih virusa imaju više sličnosti sa genima iz biljnih ćelija, nego sa životinjskim virusima. Suština procesa nasleđa, nasleđivanja i promenljivosti sadržana je u procesu reprodukcije organizma. Ćelija predstavlja osnovnu jedinicu organizma i nosilac je nasleđa i nasleđivanja što je nemoguće u potpunosti razumeti bez poznavanja njene fizičke i biohemijske prirode. Izučavanje uloge i funkcije ćelije i njenih organela kroz proces nasleđivanja i promenljivosti je osnovni zadatak citogenetike. Ćelija se sastoji iz tri osnovne komponente: ćelijskog zida, citoplazme i jedra, svaka od navedenih komponenti ima svoju specifičnu građu i ulogu u integralnim procesima nasleđivanja, što će biti u daljem tekstu predstavljeno.
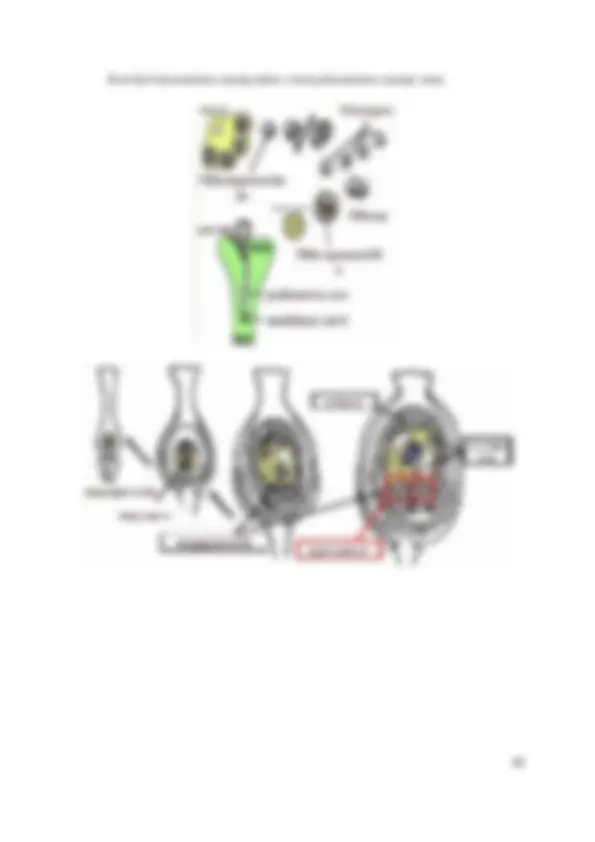
JEDRO ( NUCELUS )
Jedro (lat. nucelus , grč. karyon ) je najveća i najznačajnija organela unutar ćelije jer je u njemu DNK kao nosilac genetičkih informacija, koje regulišu i kontrolišu mnoge značajne procese u životu ćelije (replikaciju, sintezu proteina, ćelijsko disanje). Nekada se smatralo da jedro predstavlja kesicu ispunjenu tečnošću u kojoj plivaju hromozomi. Međutim, danas se zna da se u jedru nalaze: jedarce, hromozomi i visoko organizovana unutrašnja struktura (jedarni matriks, sok, citoplazma). Jedro se ne postoji kod zrelih eritrocita, ćelija spoljašnjeg sloja kože, ćelija očnog sočiva, sitastih ćelija floema kod cvetnica. Kod eukariota jedro je obavijeno dvostrukom membranom (karioteka) koja odvaja sadržaj jedra od citoplazme, a veza između njih ostvaruju se preko pora u nuklearnoj membrani. Kod prokariota nema oformljenog jedra tako da nukleinske kiseline nisu odvojene od citoplazme i formiraju strukturu označenu kao nukleoid. Većina vrsta ima jedno jedro – mononuklearne ćelije (alge, gljive, protozoe), ali postoje i ćelije koje su – binuklearne , i više – polinuklearne. Svaka ćelija eukariota ima jedno ili više jedara u toku trajanja njene aktivnosti, ili je jedro prisutno bar na nekom stupnju u toku ćelijske deobe. Za brojne vrste „nižih“ eukariota karakteristično je prisustvo dva ili više jedara. Po pravilu, gubitak jedra izaziva vrlo brzu smrt ćelije. Izuzetak u tom smislu su eritrociti sisara i ćelije floema kod cvetnica. Posle nestanka jedra eritrociti funkcionišu nekoliko meseci, dok se kod ćelija floema normalna funkcija bez jedra održava i u toku više godina. Citoplazma jedra (karioplazma, nukleoplazma) ima veću gustinu od ćelijske citoplazme, što je posledica manjeg sadržaja vode. Neorganski joni, kao što su Na, K, Cl, nalaze se u jedru u znatno višoj koncentraciji, nego u citoplazmi. Unutar jedra nema
organizatori jedarca raspoređeni na većem broju hromozoma haploidne garniture (kod čoveka ti lokusi nalaze se na pet različitih hromozoma).
HROMOZOMI: NOSIOCI GENA Hromatin i metafazni hromozomi
Najvažnija funkcija hromozoma je to da je u njima smeštena DNK - funkcionalna jedinica nasleđa. DNK predstavlja genetički materijal svih ćelija, materijalnu osnovu nasledne supstance. Genomi bakterija i nižih eukariota su vrlo kompaktni, tj. celokupna DNK sekvenca iskorišćena je za kodiranje funkcionalnih genskih produkata. Nasuprot tome, kod viših eukariota hromozomska DNK, pored gena, sadrži i velike količine nekodirajućih sekvenci. Molekuli DNK kod prokariota su cirkularnog oblika, nisu odvojeni membranom od citoplazme i zajedno sa njom čine strukturu koju nazivamo nukleoid. Često ćete naići na termin ”ogoljena DNK”, tj. DNK bez proteina.
Hromozomi se nalaze u jedru i zavisno od stadijuma ćelijskog ciklusa dosta menjaju svoj izgled. Najčešće se pod mikroskopom vide kao končaste tvorevine čija veličina i broj može da varira od jedne grupe živih bića do druge, ali je tipična za svaku vrstu. Kada se ćelija podeli, jedro i njegovi hromozomi se takođe podele. Hromozomi su vidljivi pod svetlosnim mikroskopom samo tokom deobe ćelije što je posledica spiralizacije DNK molekula hromozoma, a pod elektronskim mikroskopom tokom čitavog ćelijskog ciklusa. Razlikujemo interfazne hromozome (hromatin) i hromozome neposredno pred deobu, tokom deobe i neposredno posle deobe (hromozomi sa jednom ili dve DNK). Iako smo i danas još uvek daleko od razumevanja fine strukture hromozoma u interfazi, kod eukariota, hromatin predstavlja kompleks od DNK molekula, proteina i malih količina RNK. ( napomena - ovaj hemijski sastav hromatina i hromozoma je isti) Proteini hromatina su grupisani u dve velike klase:
Nehistonski proteini su frakcija satavljena od velikog broja jako heterogenih, kiselih proteina. Sastav ovih proteina jako varira, ne samo između vrsta, već i između različitih ćelija unutar istog organizma. Ovi proteini mogu biti regulatori genske aktivnosti. Pod elektronskim mikroskopom možemo primetiti intenzivno obojen hromatin (heterohromatin) i svetlije obojen hromatin (euhromatin). Heterohromatin je genetički neaktivan. Sastavljen je od gusto pakovanih hromatinskih vlakana i ostaje kondenzovan tokom celog ćelijskog ciklusa. Većina do danas okarakterisanih eukariotskih gena locirana je u euhromatinskim regijama hromozoma. Pri deobi ćelija (mitozi) dolazi do kondenzovanja hromatina da bi se omogućila ravnomernija raspodela jedarnog genetičkog materijala u dve ćerke ćelije. Kondenzovanjem hromatina jasno se primećuju osnovni gradivni elementi hromozoma: jedna ili dve hromatide, centromera, telomere i mesto replikacije (znači ne menja se hemijski sastav, već samo oblik i veličina). Od kraja S1 faze do kraja metafaze primećuje se hromozom koji ima dve hromatide i u svakoj hromatidi po jedan lanac DNK. Od početka anafaze do S1 faze hromozomi su sastavljeni od jedne hromatide u kojoj je jedan lanac DNK. Na početku interfaze hromozomi se dekompenzuju DNK i tada su hromozomi vidljivi kao hromatinski materijal (“rasut materijal”).
Centromere predstavljaju primarna suženja, mesta vezivanja niti deobnog vretena, koje omogućavaju kretanje hromozoma ka polovima tokom ćelijske deobe. Centromera sadrži specifične DNK sekvence na kojoj se vezuju proteini stvarajući kinetohor. Kinetohor je struktura za povezivanje hromozoma sa nitima deobnog vretena. Primarno suženje deli hromozom na dva “kraka”. Ukoliko kraci nisu jednake dužine, kraći se obeležava sa p (fran. “ petite ”=mali), a duži krak se obeležava sa q. Prema položaju centromere razlikujemo nekoliko morfoloških tipova hromozoma:
Svaki hromozom, koji ne poseduje centromeru (acentrik), ne može se pomeriti ka jednom od polova i ući u sastav novoformiranog jedra, tako da se gubi u procesu deobe ćelije. Ovakva pojava ima nesagledive posledice za ćeliju.
Sve nasledne osobine jedinki koje se prenose kroz generacije kontrolišu geni. Gen predstavlja jedinicu strukture, funkcije i transmisije naslednog materijala. Gen je deo molekula DNK (kod nekih virusa RNK) koji poseduje šifru za sintezu nekog funkcionalnog molekula RNK (iRNK, tRNK, rRNK ili snRNK). Svaki gen ima određeno mesto ( lokus ) na hromozomu. Pored određenog mesta na hromozomu, svaki gen ima određenu strukturu. Struktura gena je određena sastavom i redosledom nukleotida (osnovnih gradivnih jedinica DNK). Jedan gen se može sastojati od 400 do 1800 nukleotidnih parova. Redosled nukleotida u jednom genu - struktura gena određuje funkciju gena, odnosno informaciju o sintezi tačno određenog molekula proteina ili drugog funkcionalnog molekula RNK (tRNK, rRNK ili snRNK). Geni mogu da se jave u više različitih oblika (genski aleli). Aleli su forme jednog istog gena koji su smešteni na istom lokusu (mestu) homologih hromozoma, determinišu istu osobinu, ali se mogu kvalitativno razlikovati.
Često se gen ne javlja samo u dva različita oblika (Aa ili Bb ili Cc…), već postoji veći broj alela na datom lokusu ( multipli alelizam ) (npr. genski aleli za krvnu grupu čoveka IA^ IB^ IO). U slučaju multiplih alela, divlji tip gena ( A ) je mutirao u različito vreme u različite alelne oblike.
Napomenimo da kod diploidne jedinke, bez obzira na broj alela za jedan gen, na jednom lokusu može biti samo dva alela. Tako, čovek na lokusu za krvnu grupu ima tri
genska alela IA^ IB^ IO, ali na lokusu su aleli IA^ IB^ ili IB^ IO^ ili IA^ IO^ ili IO^ IO^ ili IA^ IA^ ili IB^ IB^. Ukoliko jedinke na jednom lokusu imaju identične genske alele, takve jedinke označavamo kao homozigotne , ukoliko se aleli na istom lokusu homologih hromozoma razlikuju te jedinke su heterozigoti.
Napomena - homozigotnost i heterozigotnost jedne jedinke se odnosi samo na posmatrane genske alele (znači, jedna jedinka može biti homozigotna za oblik AA, ali heterozigotna za boju Bb).
Homozigoti su obeleženi kao AA, BB, cc, aa, bb, IO^ IO, IA^ IA^ IB^ IB^ - istim slovima. Heterozigoti su obeleženi kao Aa, Bb, IA^ IO^ , IA^ IB^ - različitim slovima.
Hemizigot
Kada je u genotipu individue prisutan samo jedan alel nekog gena, dok drugi alel ne postoji na homologom hromozomu, tada se govori o hemizigotu. Hemizigoti su muške individue za one gene koji se nalaze samo na X hromozomu koji nije homolog sa Y hromozomom. Skup gena u jednoj hromozomskoj garnituri (setu) označava se kao genom. Međutim, ukoliko se posmatraju svi nasledni činioci (cela genetička konstitucija) u jednom organizmu, mi ustvari posmatramo genotip. Iako genotip obuhvata sve gene jedne jedinke, radi lakše analize mi često posmatramo jedan lokus (ili nekoliko) i onda objasnimo genotip za taj lokus. Npr. ukrštanje homozigotnih, dominantnih alela za oblik zrna kod graška daje genotip AA za okruglo zrno dok ukrštanje heterozigotnih alela za boju zrna graška daće 3 genotipa – BB i Bb za žutu i genotip bb za zelenu (žuta : zelena=3:1). Organizam čoveka je diploidan (2n), kariotip čoveka ima 46 hromozoma, genom sadrži 23 hromozoma (samo jedan set označava se kao genom). Pšenica je heksaploid (6n), kariotip pšenice sadrži 42 hromozoma, genom sadrži 7 hromozoma. U svakom organizmu postoji veliki broj gena. Dekodiranjem humanog genoma utvrđeno je da je broj gena koji kodiraju proteine između 20.000 i 25.000. Genotip nosi informaciju za razviće karakterističnog izleda organizma ( fenotip ). Znači, fenotip se ostvaruje u konkretnim uslovima sredine (ekspresija genotipa sa nizom morfoloških, fizioloških i ponašajnih osobina).
Genotip+sredina →Fenotip
Geni predstavljaju predispoziciju da se razvije odgovarajući fenotip, dok uslovi sredine mogu predstavljati ogranićavajući faktor te ekspresije. Ekspresija jednog gena ograničena je i interakcijom sa drugim genima. Takođe, pod uticajem sredine
Ukoliko ima 4 genska alela (navedeni primer boje kunića) javiće se 10 genotipova, dok će se kod nasleđivanja krvne grupe gde ima tri genska alela za ovu osobinu javiti 6 genotipa (IA^ IB^ ; IB^ IO; IA^ IO; IO^ IO; IA^ IA; IB^ IB^ ).
Inkompatibilnost kod biljaka
Kod nekih biljnih vrsta, koje imaju dvopolne cvetove, postoji čitava serija alela inkompatibilnosti, koji čine te biljne vrste stranooplodnim ili ksenogamnim biljkama. Broj alela inkompatibilnosti je različit kod različitih vrsta (raž, jabuka, višnja, tršnja, orah, detelina). Ovaj gen se tokom evolucije razvio u veći broj alela. Kod inkompatibilnosti, majka biljka ne prihvata polen koji nosi alel za inkompatibilnost. Na ovaj način se forsira stranooplodnja.
Primer: Ako i polenovo zrno i plodnik cveta sadrže iste allele za inkompatibilnost taj polen neće klijati i neće doći do oplodnje Ako se u plodniku nalaze aleli za inkompatibilnost S 1 S 2 , samo polenova zrna koja nemaju ove alele S 1 S 2 će proklijati kroz žig tučka (znači ako imaju alele npr. S 3 S 4 ili S 3 S 5 ).
(Dominantno-recesivno ukrštanje)
Ove zakone je formulisao 1865. godine češki biolog (kaluđer), sa austrijskim državljanstvom, Gregor Mendel, a tiču se raspodele gena i osobina u tri sukcesivne generacije monohibridnog ukrštanja različitih sorti baštenskog graška. Mendel je pratio osobine graška koje se ispoljavaju alternativno.
Alternativne osobine koje je Mendel proučavao
Osobina Dominantna Recesivna
Ljubičasta Bela Žuta Zelena Okruglo Naborano Zelena Žuta Nabubrela Sužena Bočni Terminalni Visoka Niska
Grašak je diploidna (sadrži dva seta hromozoma), samooplodna biljka. Da je za istraživanja izabrao neku poliploidnu biljku (što je česta pojava kod biljaka), nikada ne bi dobio tako razumljive rezultate. Za ukrštanja, Mendel je koristio genotipove graška sa visokim stepenom homozigotnosti (čiste linije), zahvaljujući samooplodnji iz generacije u generaciju. Mendel je vršio ukrštanje čistih linija tako što je sa cvetova jedne linije uklanjao prašnike i na tučak nanosio polenov prah druge linije, koja je ispoljavala alternativno svojstvo. Roditeljsku generaciju označio je sa P (parentalna), a dve generacije potomaka sa F 1 (prva filijalna) i F 2 (druga filijalna generacija).
ispoljena osobina u F 1 generaciji dominantna (npr. okruglo zrno), dok je alternativna osobina recesivna (npr. smežurano zrno) i maskirana u F 1 generaciji. Znači, nasledni faktor se javlja u različitim oblicima (danas poznati aleli) gde jedan oblik kontroliše okruglo seme (osobinu u F 1 generaciji) i to je dominantni alel, dok drugi oblik naslednog faktora kontroliše naborano zrno, koji se ispoljava u F 2 generaciji te je taj oblik recesivan (recesivni alel). Oblik zrna ali i ostale posmatrane osobine su, dakle, određene naslednim faktorom koji se javlja u dominantnom (alel A) i recesivnom obliku (alel a).
Nasleđivanje boje zrna (monohibridno ukrštanje)
Kada je Mendel ukrstio hibridne biljke iz F 1 generacije sa recesivnim roditeljem iz P generacije dobio je jedinke i sa dominantnom osobinom (okruglim zrnom) i sa recesivnom osobinom (smežurano zrno) u odnosu 1:1. Ovo ukrštanje je nazvano povratno ukrštanje. Međutim, ovakvo ukrštanje je i test ukrštanje jer predstavlja dodatnu potvrdu pravila rastavljanja, uz napomenu da se rastavljanje odvija samo kod heterozigota (Aa) koji stvara dva tipa gameta (A i a).
Test ukrštanje monohibrida P: ♀ Aa x ♂ aa F1:
♀/ ♂ (^) a Fenotip
A (^) Aa Žuto 1
a (^) aa Zeleno 1
Dihibridno ukrštanje
Mendel je ukrštao biljke graška koje su se razlikovale u dva para alela (za dve osobine) i pratio je nasleđivanje dve fenotipske osobine. Takvo ukrštanje se naziva dihibridno. Ukrštanjem čiste linije graška sa dve dominante osobine (okruglo i žuto zrno) i linije sa dve recesivne osobine (smežurano i zeleno), Mendel je u F 1 generaciji dobio hibridne biljke isključivo sa okruglim i žutim zrnima (uniformne po genotipu i fenotipu).
Kada je dozvolio samooplodnju kod F 1 hibrida, Mendel je u F 2 generaciji zapazio da pored roditeljskih kombinacija fenotipova (okruglo i žuto, kao i smežurano i zeleno) postoje i dva nova fenotipa (okruglo i zeleno; smežurano i žuto), koja predstavljaju nove kombinacije gena. Analizirajući veći broj semena, utvrdio je da je brojni odnos fenotipova u F 2 generaciji 9:3:3:1 (okruglo i žuto: okruglo i zeleno: smežurano i žuto: smežurano i zeleno) Ako okruglo seme određuje dominantan alel predstavljen simbolom A, a smežurano seme određuje recesivan alel predstavljen simbolom a, i ako žutu boju određuje dominantni alel predstavljen simbolom B, a zelenu boju recesivni alel predstavljen simbolom b, nasleđivanje boje i oblika graška se može predstaviti na sledeći način (Punetova mreža) :
A-Žuta boja a-zelena B- Okruglo b- smežurano
♀/ ♂ AB Ab aB ab AB AABB AABb AaBB AaBb Ab AABb AAbb AaBb Aabb aB AaBB AaBb aaBB aaBb ab AaBb Aabb aaBb aabb
U dihibridnom ukrštanju Mendel je dobio rekombinovane fenotipove (fenotipove koji se razlikuju od roditelja po jednoj osobini - okruglo i zeleno zrno; smežurano i žuto zrno). Zaključio je da je ovde došlo i do rastavljanja (segregacije) pojedinačnih parova genskih alela, ali i do nezavisnog kombinovanja alelnih parova različitih gena. Pravilo slobodnog kombinovanja (treće Mendelovo pravilo) ukazuje da se različiti parovi genskih alela slobodno kombinuju tokom gametogeneze. Mendel nije mogao u potpunosti da objasni ovu
P: ♀ AABB x ♂ aabb F1: ♀/ ♂ ab
AB AaBb
F 1 : ♀ AaBb x ♂ AaBb