Download cell biology chapter 11 and more Summaries Cell Biology in PDF only on Docsity!
THE LIPID BILAYER
MEMBRANE PROTEINS
A living cell is a self-reproducing system of molecules held inside a con- tainer. That container is the plasma membrane—a protein-studded, fatty film so thin that it cannot be seen directly in the light microscope. Every cell on Earth uses such a membrane to separate and protect its chemical components from the outside environment. Without membranes, there would be no cells, and thus no life. The structure of the plasma membrane is simple: it consists of a two-ply sheet of lipid molecules about 5 nm—or 50 atoms—thick, into which pro- teins have been inserted. Its properties, however, are unlike those of any sheet of material we are familiar with in the everyday world. Although it serves as a barrier to prevent the contents of the cell from escaping
and mixing with molecules in the surrounding environment (Figure
11! 1 ), the plasma membrane does much more than that. If a cell is to
survive and grow, nutrients must pass inward across the plasma mem- brane, and waste products must make their way out. To facilitate this plasma membrane internal membrane (A) BACTERIAL CELL (B) EUKARYOTIC CELL Figure 11–1 Cell membranes act as selective barriers. The plasma membrane separates a cell from its surroundings, enabling the molecular composition of a cell to differ from that of its environment. (A) In some bacteria, the plasma membrane is the only membrane. (B) In addition to a plasma membrane, eukaryotic cells also have internal membranes that enclose individual organelles. All cell membranes prevent molecules on one side from freely mixing with those on the other, as indicated schematically by the colored dots. Membrane Structure CHAPTER ELEVEN
366 CHAPTER 11 Membrane Structure exchange, the membrane is penetrated by highly selective channels and transporters—proteins that allow specific, small molecules and ions to be imported and exported. Other proteins in the membrane act as sensors, or receptors, that enable the cell to receive information about changes in its environment and respond to them in appropriate ways. The mechani- cal properties of the plasma membrane are equally impressive. When a cell grows, so does its membrane: this remarkable structure enlarges in area by adding new membrane without ever losing its continuity, and it can deform without tearing, allowing the cell to move or change shape
(Figure 11! 2 ). The membrane is also self-healing: if it is pierced, it nei-
ther collapses like a balloon nor remains torn; instead, the membrane quickly reseals. As shown in Figure 11–1, many bacteria have only a single membrane— the plasma membrane—whereas eukaryotic cells also contain internal membranes that enclose intracellular compartments. The internal mem- branes form various organelles, including the endoplasmic reticulum,
Golgi apparatus, endosomes, and mitochondria (Figure 11–3). Although
these internal membranes are constructed on the same principles as the plasma membrane, they differ subtly in composition, especially in their resident proteins. Regardless of their location, all cell membranes are composed of lipids
and proteins and share a common general structure (Figure 11–4). The
lipids are arranged in two closely apposed sheets, forming a lipid bilayer (see Figure 11–4B). This lipid bilayer serves as a permeability barrier to most water-soluble molecules, while the proteins embedded within it carry out the other functions of the membrane and give different mem- branes their individual characteristics. In this chapter, we consider the structure of biological membranes and the organization of their two main constituents: lipids and proteins. Although we focus mainly on the plasma membrane, most of the con- cepts we discuss also apply to internal membranes. The functions of cell membranes, including their role in cell communication, the transport of small molecules, and energy generation, are considered in later chapters. receiving information 1 import and export of small molecules 2 capacity for movement and expansion 3 ECB5 e11.02/11. Figure 11–2 The plasma membrane is involved in cell communication, import and export of molecules, and cell growth and motility. (1) Receptor proteins in the plasma membrane enable the cell to receive signals from the environment; (2) channels and transporters in the membrane enable the import and export of small molecules; (3) the flexibility of the membrane and its capacity for expansion allow the cell to grow, change shape, and move. endoplasmic reticulum lysosome nucleus peroxisome endosome transport vesicle mitochondrion Golgi apparatus plasma membrane Figure 11–3 Internal membranes form many different compartments in a eukaryotic cell. Some of the main membrane-enclosed organelles in a typical animal cell are shown here. Note that the nucleus and mitochondria are each enclosed by two membranes.
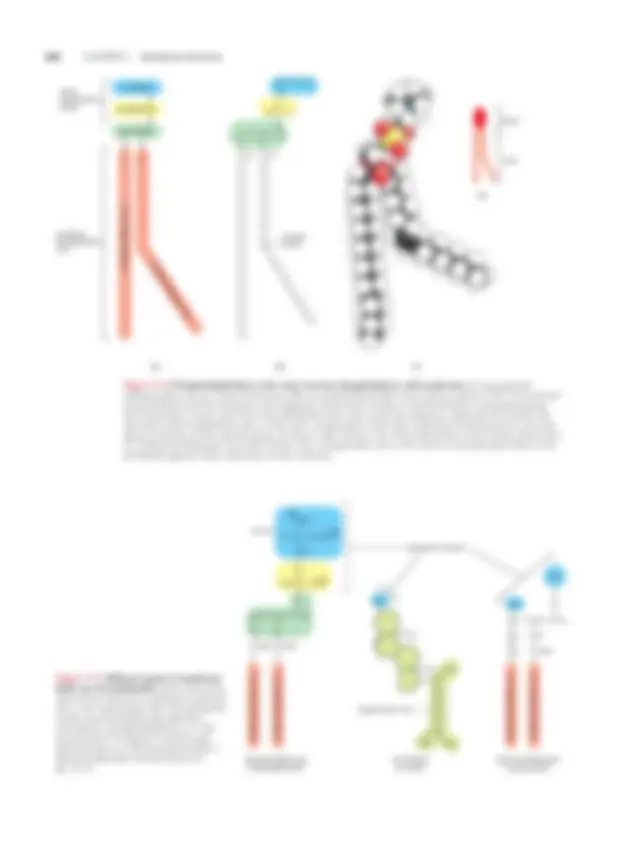
368 CHAPTER 11 Membrane Structure CHOLINE PHOSPHATE GLYCEROL HYDROCARBON TAIL HYDROCARBON TAIL polar (hydrophilic) head nonpolar (hydrophobic) tails (A) (B) CH 2 CH 2 CH 2 CH 2 CH 2 CH 2 CH 2 CH 2 CH 2 CH 2 CH 2 CH 2 CH 2 CH 2 CH 2 CH 2 CH 3 C O O CH 2 CH 2 CH 2 CH 2 CH 2 CH 2 CH 2 CH 2 CH C O O CH CH 2 O P O O_ CH 2 CH 2 N+(CH 3 ) 3 O CH double bond head tails (D) (C) 1 2 ECB5 e11.06/11. CH 2 CH 2 CH 2 CH 2 CH 2 CH 2 CH 2 CH 3 C CH 2 O NH 3
O P O O CH 2 CH CH 2 O O C O C O HYDROCARBON T AIL HYDROCARBON T AIL hydrocarbon tail HYDROCARBON T AIL HYDROCARBON T CH AIL CH 3 CH 3 CH 2 CH 2 CH 2 CH 3 CH 3 CH 3 CH OH Gal O CH CH 2 CH CH CH NH C O OH phosphatidylserine (a phospholipid) cholesterol (a sterol) galactocerebroside (a glycolipid) H COO serine hydrophilic heads Figure 11–7 Different types of membrane lipids are all amphipathic. Each of the three types shown here has a hydrophilic head and one or two hydrophobic tails. The hydrophilic head is serine phosphate (shaded blue and yellow ) in phosphatidylserine, an –OH group ( blue ) in cholesterol, and the sugar galactose plus an –OH group (both blue ) in galactocerebroside. See also Panel 2–4, pp. 72–73. Figure 11–6 Phosphatidylcholine is the most common phospholipid in cell membranes. It is represented schematically in (A), as a chemical formula in (B), as a space-filling model in (C), and as a symbol in (D). This particular phospholipid is built from five parts: the hydrophilic head, which consists of choline linked to a phosphate group; two hydrocarbon chains , which form the hydrophobic tails; and a molecule of glycerol, which links the head to the tails. Each of the hydrophobic tails is a fatty acid —a hydrocarbon chain with a carboxyl (–COOH) group at one end; glycerol attaches via this carboxyl group, as shown in (B). A kink in one of the hydrocarbon chains occurs where there is a double bond between two carbon atoms. (The “phosphatidyl” part of the name of a phospholipid refers to the phosphate–glycerol–fatty acid portion of the molecule.)
As discussed in Chapter 2 (see Panel 2–2, pp. 68–69), hydrophilic molecules dissolve readily in water because they contain either charged groups or uncharged polar groups that can form electrostatic attractions or hydro-
gen bonds with water molecules (Figure 11–8). Hydrophobic molecules,
by contrast, are insoluble in water because all—or almost all—of their atoms are uncharged and nonpolar; they therefore cannot form favora- ble interactions with water molecules. Instead, they force adjacent water
molecules to reorganize into a cagelike structure around them (Figure
11–9). Because this cagelike structure is more highly ordered than the
rest of the water, its formation requires free energy. This energy cost is minimized when the hydrophobic molecules cluster together, limiting their contacts with the surrounding water molecules. Thus, purely hydro- phobic molecules, like the fats found in the oils of plant seeds and the
adipocytes (fat cells) of animals (Figure 11–10), coalesce into large fat
droplets when dispersed in water. Figure 11–8 A hydrophilic molecule attracts water molecules. Both acetone and water are polar molecules: thus acetone readily dissolves
in water. Polar atoms are shown in red and blue , with δ–^ indicating a
partial negative charge, and δ+^ indicating a partial positive charge.
Hydrogen bonds ( red ) and an electrostatic attraction ( yellow ) form between acetone and the surrounding water molecules. Nonpolar groups are shown in gray. Figure 11–9 A hydrophobic molecule tends to avoid water. Because the 2-methylpropane molecule is entirely hydrophobic, it cannot form favorable interactions with water. This causes the adjacent water molecules to reorganize into a cagelike structure around it, to maximize their hydrogen bonds with each other. QUESTION 11– Water molecules are said “to reorganize into a cagelike structure” around hydrophobic compounds (e.g., see Figure 11–9). This seems paradoxical because water molecules do not interact with the hydrophobic compound. So how could they “know” about its presence and change their behavior to interact differently with one another? Discuss this argument and, in doing so, develop a clear concept of what is meant by a “cagelike” structure. How does it compare to ice? Why would this cagelike structure be energetically unfavorable? water δ+^ δ+ δ _ HC CH 3 CH 3 CH 3 HC CH 3 CH 3 CH 3 2-methylpropane in water 2-methylpropane H (^) H O C^ O CH 3 CH 3 C O CH 3 CH 3 water acetone in water acetone δ+ δ+^ δ+ δ _ δ _ ECB5 e11.08/11. H (^) H O hydrogen bonds The Lipid Bilayer
sets a lower limit of about 25 nm to the vesicle diameter that cell mem- branes can form. The fluidity of lipid bilayers can be studied using synthetic lipid bilayers, which are easily produced by the spontaneous aggregation of amphi- pathic lipid molecules in water. Pure phospholipids, for example, will form closed, spherical vesicles, called liposomes , when added to water; these vesicles vary in size from about 25 nm to 1 mm in diameter
(Figure 11–13).
Using such simple synthetic bilayers, investigators can measure the movements of the lipid molecules in a lipid bilayer. These measure- ments reveal that some types of movement are rare, while others are frequent and rapid. Thus, in synthetic lipid bilayers, phospholipid mole- cules very rarely tumble from one half of the bilayer, or monolayer, to the other. Without proteins to facilitate the process, it is estimated that this event, called “flip-flop,” occurs less than once a month for any individual lipid molecule under conditions similar to those in a cell. On the other hand, as the result of random thermal motions, lipid molecules continu- ously exchange places with their neighbors within the same monolayer. This exchange leads to rapid lateral diffusion of lipid molecules within the plane of each monolayer, so that, for example, a lipid in an artifi- cial bilayer may diffuse a length equal to that of an entire bacterial cell
(~2 μm) in about one second.
Similar studies show that individual lipid molecules not only flex their hydrocarbon tails, but they also rotate rapidly about their long axis— some reaching speeds of 500 revolutions per second. Studies of whole cells—and of isolated cell membranes—indicate that lipid molecules in cell membranes undergo the same movements as they do in synthetic bilayers. The movements of membrane phospholipid molecules are sum-
marized in Figure 11–14.
The Fluidity of a Lipid Bilayer Depends on Its Composition The fluidity of a cell membrane—the ease with which its lipid molecules move within the plane of the bilayer—is important for membrane func- tion and has to be maintained within certain limits. Just how fluid a lipid bilayer is at a given temperature depends on its phospholipid composi- tion and, in particular, on the nature of the hydrocarbon tails: the closer and more regular the packing of the tails, the more viscous and less fluid the bilayer will be. Two major properties of hydrocarbon tails affect how tightly they pack in the bilayer: their length and the number of double bonds they con- tain. A shorter chain length reduces the tendency of the hydrocarbon tails to interact with one another and therefore increases the fluidity of the bilayer. The hydrocarbon tails of membrane phospholipids vary in length between 14 and 24 carbon atoms, with 18 or 20 atoms being the most common. For most phospholipids, one of these hydrocarbon tails contains only single bonds between its adjacent carbon atoms, whereas the other tail includes one or more double bonds (see Figure 11–6). The chain that harbors a double bond does not contain the maximum num- ber of hydrogen atoms that could, in principle, be attached to its carbon backbone; it is thus said to be unsaturated with respect to hydrogen. The Figure 11–12 Phospholipid bilayers spontaneously close in on themselves to form sealed compartments. The closed structure is stable because it avoids the exposure of the hydrophobic hydrocarbon tails to water, which would be energetically unfavorable. Figure 11–13 Pure phospholipids can form closed, spherical liposomes. (A) An electron micrograph of phospholipid vesicles, or liposomes. (B) A drawing of a small, spherical liposome seen in cross section. (A, courtesy of Jean Lepault.) (A) (B) water water 25 nm 50 nm The Lipid Bilayer ENERGETICALLY UNFAVORABLE in a planar phospholipid bilayer, hydrophobic tails (white layer) are exposed to water along the edges formation of a sealed compartment shields hydrophobic tails from water ENERGETICALLY FAVORABLE ECB5 e11.12-11.
372 CHAPTER 11 Membrane Structure hydrocarbon tail with no double bonds has a full complement of hydrogen atoms and is said to be saturated. Each double bond in an unsaturated tail creates a small kink in the tail (see Figure 11–6), which makes it more difficult for the tails to pack against one another. For this reason, lipid bilayers that contain a large proportion of unsaturated hydrocarbon tails are more fluid than those with lower proportions. In bacterial and yeast cells, which have to adapt to varying temperatures, both the lengths and the degree of saturation of the hydrocarbon tails in the bilayer are adjusted constantly to maintain a membrane with a relatively consistent fluidity: at higher temperatures, for example, the cell makes membrane lipids with tails that are longer and that contain fewer double bonds. A similar trick is used in the manufacture of margarine from vegetable oils. The fats produced by plants are generally unsatu- rated and therefore liquid at room temperature, unlike animal fats such as butter or lard, which are generally saturated and therefore solid at room temperature. To produce margarine, vegetable oils are “hydrogen- ated”: this addition of hydrogen removes their double bonds, making the oils more solid and butterlike at room temperature. In animal cells, membrane fluidity is modulated by the inclusion of the sterol cholesterol. This molecule is present in especially large amounts in the plasma membrane, where it constitutes approximately 20% of the lipids in the membrane by weight. With its short and rigid steroid ring structure, cholesterol can fill the spaces between neighboring phospho- lipid molecules left by the kinks in their unsaturated hydrocarbon tails
(Figure 11–15). In this way, cholesterol tends to stiffen the bilayer, mak-
ing it less flexible, as well as less permeable. The chemical properties of membrane lipids—and how they affect membrane fluidity—are reviewed
in Movie 11.3 and Movie 11.4.
For all cells, membrane fluidity is important for a number of reasons. It enables many membrane proteins to diffuse rapidly in the plane of the bilayer and to interact with one another, as is crucial, for example, in cell signaling (discussed in Chapter 16). It permits membrane lipids and proteins to diffuse from sites where they are inserted into the bilayer after their synthesis to other regions of the cell. It ensures that membrane mol- ecules are distributed evenly between daughter cells when a cell divides. And, under appropriate conditions, it allows membranes to fuse with one another and mix their molecules (discussed in Chapter 15). If biological flexion rotation lateral diffusion flip-flop (rarely occurs) ECB5 e11.14/11. Figure 11–14 Membrane phospholipids move within the lipid bilayer. Because of these motions, the bilayer behaves as a two- dimensional fluid, in which the individual lipid molecules are able to move in their own monolayer. Note that lipid molecules do not move spontaneously from one monolayer to the other. Figure 11–15 Cholesterol tends to stiffen cell membranes. (A) The shape of a cholesterol molecule. The chemical formula of cholesterol is shown in Figure 11–7. (B)!How cholesterol fits into the gaps between phospholipid molecules in a lipid bilayer. (C) Space-filling model of the bilayer, with cholesterol molecules in green. Although the nonpolar hydrocarbon tail of cholesterol is shown in green —to visually distinguish it from the hydrocarbon tails of the membrane phospholipids—in reality, the hydrophobic tail of cholesterol is chemically equivalent to the hydrophobic tails of the phospholipids. (C, from H.L. Scott, Curr. Opin. Struct. Biol. 12:495–502, 2002.) QUESTION 11– Five students in your class always sit together in the front row. This could be because (A) they really like each other or (B) nobody else in your class wants to sit next to them. Which explanation holds for the assembly of a lipid bilayer? Explain. Suppose, instead, that the other explanation held for lipid molecules. How would the properties of the lipid bilayer be different? 6 nm (A) (B) rigid planar steroid ring structure nonpolar hydrocarbon tail polar head group phospholipid cholesterol polar head cholesterol- stiffened region more fluid region (C)
374 CHAPTER 11 Membrane Structure The action of these flippases—and of similar transporters in the plasma membrane—initiates and maintains the asymmetric arrangement of phospholipids that is characteristic of the membranes of animal cells. This asymmetry is preserved as membranes bud from one organelle and fuse with another—or with the plasma membrane. This means that all cell membranes have distinct “inside” and “outside” faces: the cytosolic monolayer always faces the cytosol, while the noncytosolic monolayer is exposed to either the cell exterior—in the case of the plasma mem- brane—or the interior space ( lumen ) of an organelle. This conservation of orientation applies not only to the phospholipids that make up the mem- brane, but also to any proteins that might be inserted in the membrane
(Figure 11–18). This positioning is very important, as a protein’s orienta-
tion within the lipid bilayer is crucial for its function (see Figure 11–20). Among lipids, those that show the most dramatically lopsided distribu- tion in cell membranes are the glycolipids, which are located mainly in the plasma membrane, and only in the noncytosolic half of the bilayer
(Figure 11–19). The sugar groups of these membrane lipids face the cell
exterior, where they form part of a continuous coat of carbohydrate that surrounds and protects animal cells. Glycolipid molecules acquire their sugar groups in the Golgi apparatus, where the enzymes that engineer this chemical modification are confined. These enzymes are oriented such that sugars are added only to lipid molecules in the noncytosolic half of the bilayer. Once a glycolipid molecule has been created in this way, it remains trapped in this monolayer, as there are no flippases that transfer glycolipids to the cytosolic side. Thus, when a glycolipid mol- ecule is finally delivered to the plasma membrane, it displays its sugars to the exterior of the cell. Other lipid molecules show different types of asymmetric distributions, which relate to their specific functions. For example, the inositol phos- pholipids—a minor component of the plasma membrane—have a special role in relaying signals from the cell surface into the cell interior (dis- cussed in Chapter 16); thus they are concentrated in the cytosolic half of the lipid bilayer.
ECB5 E11.17/11.
membrane of Golgi apparatus plasma membrane CYTOSOL LUMEN noncytosolic face^ EXTRACELLULAR FLUID cytosolic face transport vesicle membrane glycoprotein Figure 11–18 Membranes retain their orientation during transfer between cell compartments. Membranes are transported by a process of vesicle budding and fusing. Here, a vesicle is shown budding from the Golgi apparatus and fusing with the plasma membrane. Note that the orientations of both the membrane lipids and proteins are preserved during the process: the original cytosolic surface of the lipid bilayer ( pink ) remains facing the cytosol, and the noncytosolic surface ( red ) continues to face away from the cytosol, toward the lumen of the Golgi and the transport vesicle—or toward the extracellular fluid. Similarly, the glycoprotein shown here ( blue and green ) remains in the same orientation, with its attached sugar facing the noncytosolic side. EXTRACELLULAR SPACE CYTOSOL plasma membrane Figure 11–19 Phospholipids and glycolipids are distributed asymmetrically in the lipid bilayer of an animal cell plasma membrane. Phosphatidylcholine ( red ) and sphingomyelin ( brown ) are concentrated in the noncytosolic monolayer, whereas phosphatidylserine ( light green ) and phosphatidylethanolamine ( yellow ) are found mainly on the cytosolic side. In addition to these phospholipids, phosphatidylinositols ( dark green head group), a minor constituent of the plasma membrane, are shown in the cytosolic monolayer, where they participate in cell signaling. Glycolipids are drawn with hexagonal blue head groups to represent sugars; these are found exclusively in the noncytosolic monolayer of the membrane. Within the bilayer, cholesterol ( green ) is distributed almost equally in both monolayers. QUESTION 11– It seems paradoxical that a lipid bilayer can be fluid yet asymmetrical. Explain.
MEMBRANE PROTEINS Although the lipid bilayer provides the basic structure of all cell mem- branes and serves as a permeability barrier to the hydrophilic molecules on either side of it, most membrane functions are carried out by mem- brane proteins. In animals, proteins constitute about 50% of the mass of most plasma membranes, the remainder being lipid plus the relatively small amounts of carbohydrate found on some of the lipids (glycolipids) and many of the proteins (glycoproteins). Because lipid molecules are much smaller than proteins, however, a cell membrane typically contains about 50 times the number of lipid molecules compared to protein mol- ecules (see Figure 11–4B). Membrane proteins serve many functions. Some transport particular nutrients, metabolites, and ions across the lipid bilayer. Others anchor the membrane to macromolecules on either side. Still others function as receptors that detect chemical signals in the cell’s environment and relay them into the cell interior, or work as enzymes to catalyze specific
reactions at the membrane (Figure 11–20 and Table 11–1). Each type of
cell membrane contains a different set of proteins, reflecting the special- ized functions of the particular membrane. In this section, we discuss the structure of membrane proteins and how they associate with the lipid bilayer. X Y TRANSPORTERS AND CHANNELS ANCHORS RECEPTORS ENZYMES CYTOSOL ECB5 e11.19/11. EXTRACELLULAR SPACE Figure 11–20 Plasma membrane proteins have a variety of functions. They transport molecules and ions, act as anchors, detect signals, or catalyze reactions. TABLE 11–1 SOME EXAMPLES OF PLASMA MEMBRANE PROTEINS AND THEIR FUNCTIONS Functional Class Protein Example Specific Function Transporters Na+^ pump actively pumps Na+^ out of cells and K+ in (discussed in Chapter 12) Ion channels K+^ leak channel allows K+^ ions to leave cells, thereby influencing cell excitability (discussed in Chapter 12) Anchors integrins link intracellular actin filaments to extracellular matrix proteins (discussed in Chapter 20) Receptors platelet-derived growth factor (PDGF) receptor binds extracellular PDGF and, as a consequence, generates intracellular signals that direct the cell to grow and divide (discussed in Chapters 16 and 18) Enzymes adenylyl cyclase catalyzes the production of the small intracellular signaling molecule cyclic AMP in response to extracellular signals (discussed in Chapter 16) Membrane Proteins
A Polypeptide Chain Usually Crosses the Lipid Bilayer as an α Helix All membrane proteins have a unique orientation in the lipid bilayer, which is essential for their function. For a transmembrane receptor pro- tein, for example, the part of the protein that receives a signal from the environment must be on the outside of the cell, whereas the part that passes along the signal must be in the cytosol (see Figure 11–20). This orientation is a consequence of the way in which membrane proteins are synthesized (discussed in Chapter 15). The portions of a transmem- brane protein located on either side of the lipid bilayer are connected by specialized membrane-spanning segments of the polypeptide chain (see Figure 11–21A). These segments, which run through the hydropho- bic environment of the interior of the lipid bilayer, are composed largely of amino acids with hydrophobic side chains. Because these side chains cannot form favorable interactions with water molecules, they prefer to interact with the hydrophobic tails of the lipid molecules, where no water is present. In contrast to the hydrophobic side chains, however, the peptide bonds that join the successive amino acids in a protein are normally polar, mak-
ing the polypeptide backbone itself hydrophilic (Figure 11–22). Because
water is absent from the interior of the bilayer, atoms that are part of the polypeptide backbone are thus driven to form hydrogen bonds with one another. Hydrogen-bonding is maximized if the polypeptide chain forms
a regular α helix, and so the great majority of the membrane-spanning
segments of polypeptide chains traverse the bilayer as α helices (see
Figure 4!12). In these membrane-spanning α helices, the hydrophobic
side chains are exposed on the outside of the helix, where they contact the hydrophobic lipid tails, while the atoms of the hydrophilic polypep- tide backbone form hydrogen bonds with one another within the helix
(Figure 11–23).
For many transmembrane proteins, the polypeptide chain crosses the membrane only once (see Figure 11–21A, left ). Many of these single- pass transmembrane proteins are receptors for extracellular signals. Other transmembrane proteins function as channels, forming aqueous pores across the lipid bilayer to allow small, water-soluble molecules to cross the membrane. Such channels cannot be formed by proteins with
a single transmembrane α helix. Instead, they usually consist of a series
of α helices that cross the bilayer a number of times (see Figure 11–21A,
center ). For many of these multipass transmembrane proteins, one or more of the membrane-spanning regions are amphipathic—formed from
α helices that contain both hydrophobic and hydrophilic amino acid side
chains. These amino acids tend to be arranged so that the hydrophobic side chains fall on one side of the helix, while the hydrophilic side chains are concentrated on the other side. In the hydrophobic environment of
the lipid bilayer, α helices of this type pack side by side in a ring, with
the hydrophobic side chains exposed to the hydrophobic lipid tails and the hydrophilic side chains forming the lining of a hydrophilic pore R C O C O C O N H N H N H C H C H R C H R δ _ δ _ δ _ δ _ δ _ δ _ peptide bonds ECB5 E11.21/11. δ+^ δ+ δ+ δ+ δ+ δ+ Figure 11–22 The backbone of a polypeptide chain is hydrophilic. The atoms on either side of a peptide bond ( red line) are polar and carry partial positive or
negative charges (δ+^ or δ–). These charges
allow these atoms to hydrogen-bond with one another when the polypeptide folds
into an α helix that spans the lipid bilayer
(see Figure 11–23). hydrophobic amino acid side chain hydrogen bond hydrophobic tails of membrane phospholipids α helix Figure 11–23 A transmembrane polypeptide chain usually crosses
the lipid bilayer as an α helix. In this segment of a transmembrane
protein, the hydrophobic side chains ( light green ) of the amino acids
forming the α helix contact the hydrophobic hydrocarbon tails of the
phospholipid molecules, while the hydrophilic parts of the polypeptide backbone form hydrogen bonds with one another (dashed red lines)
along the interior of the helix. An α helix containing about 20 amino
acids is required to completely traverse a cell membrane. Membrane Proteins
378 CHAPTER 11 Membrane Structure
through the membrane (Figure 11–24). How such channels function
in the selective transport of small, water-soluble molecules, especially inorganic ions, is discussed in Chapter 12.
Although the α helix is by far the most common form in which a poly-
peptide chain crosses a lipid bilayer, the polypeptide chain of some
transmembrane proteins crosses the lipid bilayer as a β sheet that is rolled
into a cylinder, forming a keglike structure called a β barrel (see Figure
11–21A, right ). As expected, the amino acid side chains that face the inside of the barrel, and therefore line the aqueous channel, are mostly hydrophilic, while those on the outside of the barrel, which contact the hydrophobic core of the lipid bilayer, are exclusively hydrophobic. A strik-
ing example of a β-barrel structure is found in the porin proteins, which
form large, water-filled pores in mitochondrial and bacterial outer mem-
branes (Figure 11–25). Mitochondria and some bacteria are surrounded
by a double membrane, and porins allow the passage of small nutrients, metabolites, and inorganic ions across their outer membranes, while preventing unwanted larger molecules from crossing. Membrane Proteins Can Be Solubilized in Detergents To understand a protein fully, one needs to know its structure in detail. For membrane proteins, this presents special problems. Most biochemi- cal procedures are designed for studying molecules in aqueous solution. Membrane proteins, however, are built to operate in an environment that is partly aqueous and partly fatty, and taking them out of this environ- ment to study in isolation—while preserving their essential structure—is no easy task. Before an individual protein can be examined in detail, it must be sepa- rated from all the other cell proteins. For most membrane proteins, the first step in this purification process involves solubilizing the membrane with agents that destroy the lipid bilayer by disrupting hydrophobic associations. The most widely used disruptive agents are detergents
(Movie 11.5). These small, amphipathic, lipidlike molecules differ from
membrane phospholipids in that they have only a single hydrophobic
tail (Figure 11–26). Because they have one tail, detergent molecules are
shaped like cones; in water, these conical molecules tend to aggregate into small clusters called micelles , rather than forming a bilayer as do the phospholipids, which—with their two tails—are more cylindrical in shape. When mixed in great excess with membranes, the hydrophobic ends of detergent molecules interact with the membrane-spanning hydro- phobic regions of the transmembrane proteins, as well as with the hydrophobic tails of the phospholipid molecules, thereby disrupting the lipid bilayer and separating the proteins from most of the phospho- lipids. Because the other end of the detergent molecule is hydrophilic, these interactions draw the membrane proteins into the aqueous solu- tion as protein–detergent complexes; at the same time, the detergent hydrophilic side chains form an aqueous pore amphipathic α helix ECB5 E11.23/11. lipid bilayer hydrophobic side chains interact with phospholipid tails Figure 11–24 A transmembrane hydrophilic pore can be formed by multiple amphipathic α helices. In this example, five amphipathic
transmembrane α helices form a water-filled channel across the lipid
bilayer. The hydrophobic amino acid side chains on one side of each helix ( green ) come in contact with the hydrophobic lipid tails of the lipid bilayer, while the hydrophilic side chains on the opposite side of the helices ( red ) form a water-filled pore. N C 2 nm ECB5 e11.24/11. Figure 11–25 Porin proteins form water- filled channels in the outer membrane of a bacterium. The protein illustrated is from E. coli , and it consists of a 16-stranded
β sheet curved around on itself to form a
transmembrane water-filled channel. The three-dimensional structure was determined by x-ray crystallography. Although not shown in the drawing, three porin proteins associate to form a trimer with three separate channels. QUESTION 11– Explain why the polypeptide chain of most transmembrane proteins
crosses the lipid bilayer as an α helix
or a β barrel.
380 CHAPTER 11 Membrane Structure the entire organism—a deep purple color. When retinal, which is cova-
lently attached to one of bacteriorhodopsin’s transmembrane α helices,
absorbs a photon of light, it changes shape. This shape change causes the surrounding helices to undergo a series of small conformational changes, which pump one proton from the retinal to the outside of the organism
(Figure 11–28).
In the presence of sunlight, thousands of bacteriorhodopsin mol- ecules pump H+^ out of the cell, generating a concentration gradient of H+^ across the plasma membrane. The cell uses this proton gradient to store energy and convert it into ATP, as we discuss in detail in Chapter
- Bacteriorhodopsin is a pump , a class of transmembrane protein that actively moves small organic molecules and inorganic ions into and out of cells. We will discuss the action of other important transmembrane pumps in Chapter 12. The Plasma Membrane Is Reinforced by the Underlying Cell Cortex A cell membrane by itself is extremely thin and fragile. It would require nearly 10,000 cell membranes laid on top of one another to achieve the thickness of this paper. Most cell membranes are therefore strengthened and supported by a framework of proteins, attached to the membrane via transmembrane proteins. For plants, yeasts, and bacteria, the cell’s shape and mechanical properties are conferred by a rigid cell wall —a fibrous layer of proteins, sugars, and other macromolecules that encases the plasma membrane. By contrast, the plasma membrane of animal cells is stabilized by a meshwork of filamentous proteins, called the cell cortex, that is attached to the underside of the membrane. The cortex of the human red blood cell has a relatively simple and regular structure and has been especially well studied. Red blood cells are small
and have a distinctive flattened shape (Figure 11!29A). The main com-
ponent of their cortex is the dimeric protein spectrin , a long, thin, flexible rod about 100 nm in length. Spectrin forms a lattice that provides sup- port for the plasma membrane and maintains the cell’s biconcave shape. The spectrin network is connected to the membrane through intracellular Figure 11–28 Bacteriorhodopsin acts as a proton pump. The polypeptide chain of this small protein (about 250 amino acids in
length) crosses the lipid bilayer as seven α
helices. The location of the retinal ( purple ) and the probable pathway taken by protons during the light-activated pumping cycle ( red arrows) are highlighted. Strategically placed polar amino acid side chains— shown in red , yellow , and blue —guide the movement of the proton (H+) across the bilayer, allowing it to avoid contact with the lipid environment. The retinal is then regenerated by taking up a H+^ from the cytosol, returning the protein to its original conformation—a cycle shown in Movie 11.6. Retinal is also used to detect light in our own eyes, where it is attached to a protein with a structure very similar to that of bacteriorhodopsin. (Adapted from H.!Luecke et al., Science 286:5438 255–260, 1999.) ECB5 e11.27/11. EXTRACELLULAR SPACE lipid bilayer CYTOSOL retinal HOOC NH 2 H+ H+ transmembrane helices
attachment proteins that link spectrin to specific transmembrane pro-
teins (Figure 11!29B and Movie 11.7). The importance of this meshwork
is seen in mice and humans that, due to genetic alterations, produce a form of spectrin with an abnormal structure. These individuals are ane- mic: they have fewer red blood cells than normal. The red cells they do have are spherical instead of flattened and are abnormally fragile. Proteins similar to spectrin, and to its associated attachment proteins, are present in the cortex of most animal cells. But the cortex in these cells is especially rich in actin and the motor protein myosin , and it is much more complex than that of red blood cells. Whereas red blood cells need their cortex mainly to provide mechanical strength as they are pumped through blood vessels, other cells also use their cortex to selectively take up materials from their environment, to change their shape, and to move, as we discuss in Chapter 17. In addition, cells also use their cortex to restrain the diffusion of proteins within the plasma membrane, as we see next. A Cell Can Restrict the Movement of Its Membrane Proteins Because a membrane is a two-dimensional fluid, many of its proteins, like its lipids, can move freely within the plane of the bilayer. This lateral diffusion was initially demonstrated by experimentally fusing a mouse cell with a human cell to form a double-sized hybrid cell and then moni- toring the distribution of certain mouse and human plasma membrane proteins. At first, the mouse and human proteins are confined to their own halves of the newly formed hybrid cell, but within half an hour or so the two sets of proteins become evenly mixed over the entire cell sur-
face (Figure 11–30). We describe some other techniques for studying the
movement of membrane proteins in How We Know, pp. 384–385.
The picture of a cell membrane as a sea of lipid in which all proteins float freely is too simple, however. Cells have ways of confining particular proteins to localized areas within the bilayer, thereby creating function- ally specialized regions, or membrane domains, on the surface of the cell or organelle. 5 μm ECB5 e11.28-29/11. (A) (B) spectrin dimer actin 100 nm attachment proteins transmembrane proteins Figure 11–29 A cortex made largely of spectrin gives human red blood cells their characteristic shape. (A) Scanning electron micrograph showing human red blood cells, which have a flattened, biconcave shape. These cells lack a nucleus and other intracellular organelles. (B) In the cortex of a red blood cell, spectrin dimers ( red ) are linked end-to-end to form longer tetramers. The spectrin tetramers, together with a smaller number of actin molecules, are linked together into a mesh. This network is attached to the plasma membrane by the binding of at least two types of attachment proteins (shown here in yellow and blue ) to two kinds of transmembrane proteins (shown here in green and brown ). (A, courtesy of Bernadette Chailley.) QUESTION 11– Look carefully at the transmembrane proteins shown in Figure 11!29B. What can you say about their mobility in the membrane? Membrane Proteins
the carbohydrate on the glycoproteins, proteoglycans, and glycolipids is located on the outside of the plasma membrane, where it forms a sugar
coating called the carbohydrate layer or glycocalyx (Figure 11–33).
This layer of carbohydrate helps protect the cell surface from mechanical damage. And because the oligosaccharides and polysaccharides attract water molecules, they also give the cell a slimy surface, which helps motile cells such as white blood cells squeeze through narrow spaces and prevents blood cells from sticking to one another or to the walls of blood vessels. Cell-surface carbohydrates do more than just protect and lubricate the cell, however. They have an important role in cell–cell recognition and adhesion. Transmembrane proteins called lectins are specialized to bind to particular oligosaccharide side chains. The oligosaccharide side chains of glycoproteins and glycolipids, although short (typically fewer than 15 sugar units), are enormously diverse. Unlike proteins, in which the amino acids are all joined together in a linear chain by identical peptide bonds, sugars can be joined together in many different arrangements, often forming elaborate branched structures (see Panel 2–4, pp. 72–73). Using a variety of covalent linkages, even three different sugars can form hun- dreds of different trisaccharides. The carbohydrate layer on the surface of cells in a multicellular organism serves as a kind of distinctive clothing, like a police officer’s uniform. It is characteristic of each cell type and is recognized by other cell types that basal lamina protein B tight junction protein A apical plasma membrane lateral plasma membrane basal plasma membrane ECB5 e11.32/11. Figure 11–32 Membrane proteins are restricted to particular domains of the plasma membrane of epithelial cells in the gut. Protein A ( green ) and protein B ( red ) can diffuse laterally in their own membrane domains but are prevented from entering the other domain by a specialized cell junction called a tight junction. The basal lamina ( yellow ) is a mat of extracellular matrix that supports all epithelial sheets (discussed in Chapter 20). lipid bilayer carbohydrate- rich layer = sugar unit transmembrane glycoprotein adsorbed glycoprotein glycolipid transmembrane proteoglycan CYTOSOL EXTRA- CELLULAR SPACE Figure 11–33 Eukaryotic cells are coated with sugars. This carbohydrate-rich layer is made of the oligosaccharide side chains attached to membrane glycolipids and glycoproteins, and of the polysaccharide chains on membrane proteoglycans. As shown, glycoproteins that have been secreted by the cell and then adsorbed back onto its surface can also contribute. Note that all the carbohydrate is on the external (noncytosolic) surface of the plasma membrane. Membrane Proteins
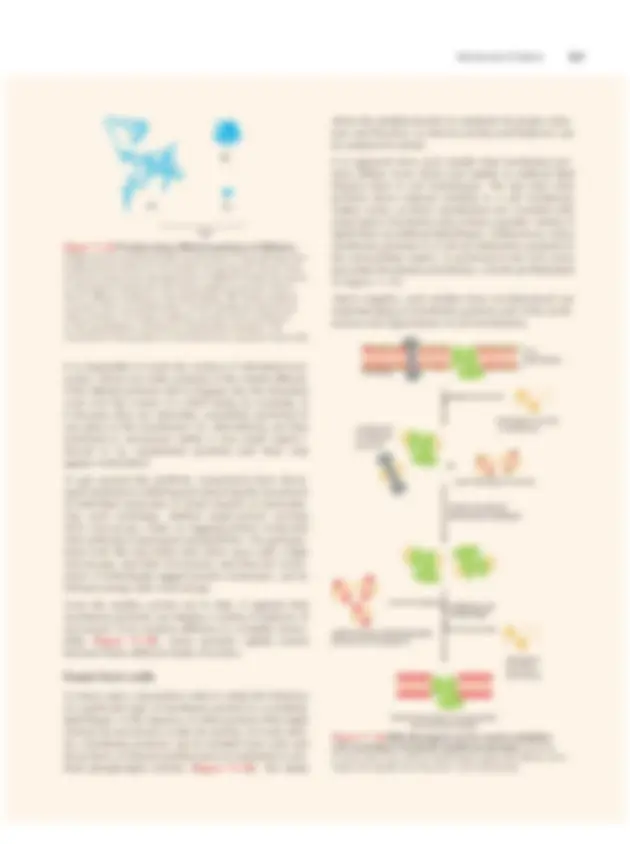
An essential feature of the lipid bilayer is its fluidity, which is crucial for cell membrane integrity and func- tion. This property allows many membrane-embedded proteins to move laterally in the plane of the bilayer, so that they can engage in the various protein–protein interactions on which cells depend. The fluid nature of cell membranes is so central to their proper function that it may seem surprising that this property was not recognized until the early 1970s. Given its importance for membrane structure and func- tion, how do we measure and study the fluidity of cell membranes? The most common methods are visual: simply label some of the molecules native to the mem- brane and then watch where they go. Such an approach first demonstrated the lateral movement of membrane proteins that had been tagged with labeled antibodies (see Figure 11–30). This experiment seemed to suggest that membrane proteins diffuse freely, without restric- tion, in an open sea of lipids. We now know that this image is not entirely accurate. To probe membrane flu- idity more thoroughly, researchers had to invent more precise methods for tracking the movement of proteins within a membrane such as the plasma membrane of a living cell. The FRAP attack One such technique, called fluorescence recovery after photobleaching ( FRAP ), involves uniformly labeling the components of the cell membrane—its lipids or, more often, its proteins—with some sort of fluorescent marker. Labeling membrane proteins can be accom- plished by incubating cells with a fluorescent antibody or by covalently attaching a fluorescent protein such as green fluorescent protein (GFP) to a membrane protein using the DNA techniques discussed in Chapter 10. Once a protein has been labeled, a small patch of mem- brane is irradiated with an intense pulse of light from a sharply focused laser beam. This treatment irreversibly “bleaches” the fluorescence from the labeled proteins in that small patch of membrane, typically an area
about 1 μm square. The fluorescence of this irradiated
membrane is monitored in a fluorescence microscope, and the amount of time it takes for the neighboring, unbleached fluorescent proteins to migrate into the
bleached region of the membrane is measured (Figure
11–34). The rate of this “fluorescence recovery” is a
direct measure of the rate at which the protein mol-
ecules can diffuse within the membrane (Movie 11.8).
Such experiments have revealed that, generally speak- ing, cell membranes are about as viscous as olive oil. One-by-one One drawback to the FRAP approach is that the tech- nique monitors the movement of fairly large populations of proteins—hundreds or thousands—across a rela- tively large area of the membrane. With this technique MEASURING MEMBRANE FLOW HOW WE KNOW BLEACH PATCH WITH LASER BEAM FLUORESCENCE RETURNED TO BLEACHED PATCH LABELED PROTEINS DIFFUSE RANDOMLY THROUGHOUT MEMBRANE BLEACH RECOVERY time fluorescence in bleached area bleached area FRAP lipid bilayer fluorescently labeled membrane proteins Figure 11–34 Photobleaching techniques such as FRAP can be used to measure the rate of lateral diffusion of a membrane protein. A specific type of protein can be labeled with a fluorescent antibody (as shown here) or tagged with a fluorescent protein, such as GFP. A small area of the membrane containing these fluorescent protein molecules is then bleached using a laser beam. As the bleached molecules diffuse away, and unbleached, fluorescent molecules diffuse into the area, the intensity of the fluorescence is recovered (shown here in side and top views). The diffusion coefficient is then calculated from a graph of the rate of fluorescence recovery: the greater the diffusion coefficient of the membrane protein, the faster the recovery.