Download Calcium Signaling: The Role of Calmodulin and Ca2+/Calmodulin-Dependent Protein Kinases and more Slides Cell Biology in PDF only on Docsity!
Some G Proteins Signal Via Phospholipids
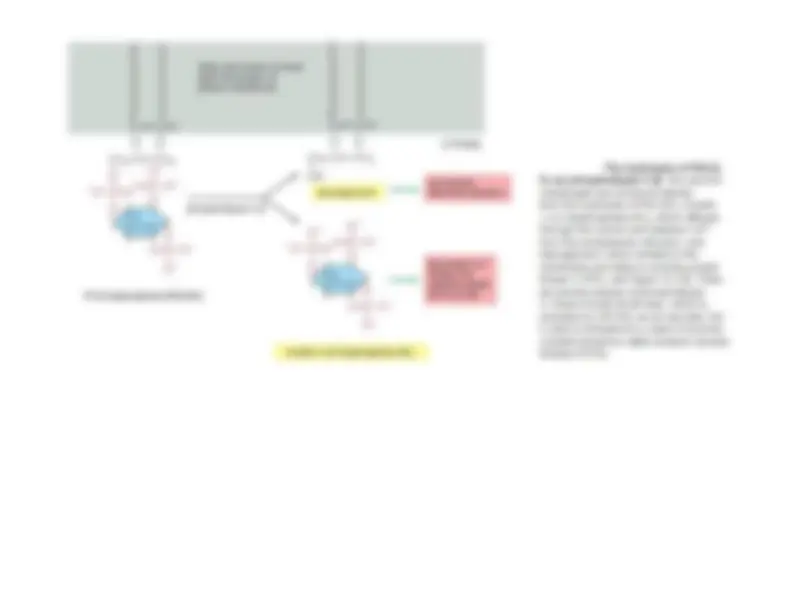
- Many GPCRs exert their effects through G proteins that activate the plasma-membrane bound enzyme phospholipase C-β (PLCβ).
- The phospholipase acts on a phosphorylated inositol phospholipid (a phospho inositide) called phosphatidyl inositol 4,5-bisphosphate [PI(4,5)P2], which is present in small amounts in the inner half of the plasma membrane lipid bilayer.
Ca2+/Calmodulin-Dependent Protein Kinases Mediate Many
Responses to Ca2+ Signals
- Various Ca2+ binding proteins help to relay the cytosolic Ca2+ signal. The most important is calmodulin, which is found in all eukaryotic cells and can constitute as much as 1% of a cell's total protein mass.
- Calmodulin functions as a multipurpose intracellular Ca2+ receptor, governing many Ca2+regulated processes.
- It consists of a highly conserved, single polypeptide chain with four high-affinity Ca2+binding sites.
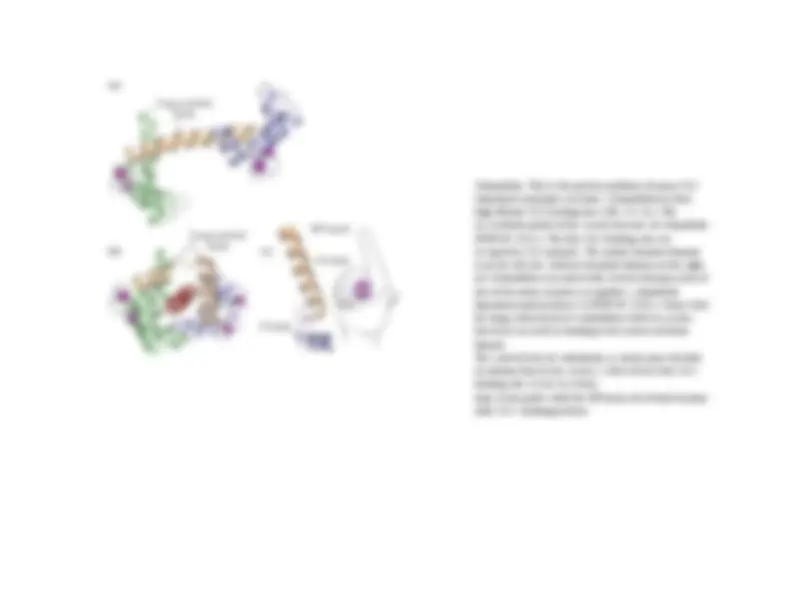
The structure of Ca2+/calmodulin. (A) The molecule has a dumbbell shape, with two globular ends, which can bind to many target proteins. The globular ends are connected by a long, exposed α helix, which allows the protein to adopt a number of different conformations, depending on the target protein it interacts with. Each globular head has two Ca2+ binding sites (B) Shown is the major structural change that occurs in Ca2+/calmodulin when it binds to a target protein (in this example, a peptide that consists of the Ca2+/calmodulin-binding domain of a Ca2+/calmodulin dependent protein kinase). Note that the
- In some cases, calmodulin serves as a permanent regulatory subunit of an enzyme complex, but usually the binding of Ca2+ instead enables calmodulin to bind to various target proteins in the cell to alter their activity.
- When an activated molecule of Ca2+/calmodulin binds to its target protein, the calmodulin further changes its conformation, the nature of which depends on the specific target protein.
- Among the many targets calmodulin regulates are enzymes and membrane transport proteins. As one example, Ca2+/ calmodulin binds to and activates the plasma membrane Ca2+pump that uses ATP hydrolysis to pump Ca2+ out of cells.
- Thus, whenever the concentration of Ca2+ in the cytosol rises, the pump is activated, which helps to return the cytosolic Ca2+ level to resting levels. Many effects of Ca2+, however, are more indirect and are mediated by protein phosphorylation catalyzed by a family of protein kinases called Ca2+/calmodulin- dependent kinases (CaM- kinases).
- It constitutes up to 2% of the total protein mass in some regions of the brain, and it is highly concentrated in synapses.
- CaM-kinase II has several remarkable properties. To begin with, it has a spectacular quaternary structure: twelve copies of the enzyme are assembled into a stacked pair of rings, with kinase domains on the outside linked to a central hub.
Calcium Is a Second Messenger That May Be Localized in Space
and Time
- There are many variations on this basic scheme for Ca2+ signalling.
- In many cell types that respond to extracellular signals, Ca2+ serves as a second messenger that triggers intracellular responses, such as exocytosis in neurons and endocrine cells, contraction in muscle, and cytoskeletal rearrangements during amoeboid movement.
- In unstimulated cells, cytosolic [Ca2+] is kept very low (107 M) by the action of Ca2+ pumps in the ER, mitochondria, and plasma membrane.
Calmodulin. This is the protein mediator of many Ca 2 stimulated enzymatic reactions. Calmodulin has four high affinity Ca 2 binding sites (Kd ≈ 0.1 to 1 M). (a) A ribbon model of the crystal structure of calmodulin (PDB ID 1CLL). The four Ca 2 binding sites are occupied by Ca 2 (purple). The amino-terminal domain is on the left; the carboxyl-terminal domain on the right. (b) Calmodulin associated with a helical domain (red) of one of the many enzymes it regulates, calmodulin dependent protein kinase II (PDB ID 1CDL). Notice that the long central helix of calmodulin visible in (a) has bent back on itself in binding to the helical substrate domain. The central helix of calmodulin is clearly more flexible in solution than in the crystal. c) Each of the four Ca 2 - binding sites occurs in a helix loop- helix motif called the EF hand, also found in many other Ca 2 - binding proteins.
Ca2+/ calmodulin-dependent protein kinases
- Calmodulin associates with a variety of proteins and, in its Ca2+ bound state, modulates their activities.
- It is a member of a family of Ca2+ binding proteins that also includes troponin , which triggers skeletal muscle contraction in response to increased [Ca2+].
- This family shares a characteristic Ca2+ binding structure, the EF hand. Calmodulin is an integral subunit of the Ca2+/calmodulin-dependent protein kinases (CaM kinases, types I through IV).
- The activity of the second messenger Ca2+, like that of cAMP, can be spatially restricted; after its release triggers a local response, Ca2+ is generally removed before it can diffuse to distant parts of the cell.
- Very commonly, Ca2+ level does not simply rise and then decrease, but rather oscillates with a period of a few seconds even when the extracellular concentration of the triggering hormone remains constant.
- The mechanism underlying [Ca2+] oscillations presumably entails feedback regulation by Ca2+ on some part of the Ca2+ release process.
- Whatever the mechanism, the effect is that one kind of signal (hormone concentration, for example) is converted into another (frequency and amplitude of intracellular [Ca2+] “spikes”).
- Another variation is the occurrence of localized Ca2+ “blips,” “puffs,” and “waves”—transient increases in [Ca2+] that are limited to specific subcellular regions
- The Ca2+ signal diminishes as Ca2+ diffuses away from the initial source (the Ca2+ channel), is sequestered in the ER, or is pumped out of the cell.
- There is significant cross-talk between the Ca2+ and cAMP signaling systems.
- In some tissues, both the enzyme that produces cAMP (adenylyl cyclase) and the enzyme that degrades cAMP (phosphodiesterase) are stimulated by Ca2+.
- Temporal and spatial changes in [Ca2+] can therefore produce transient, localized changes in [cAMP].
- It is noted that PKA, the enzyme that responds to cAMP, is often part of a highly localized supra molecular complex assembled on scaffold proteins such as AKAPs.
- This subcellular localization of target enzymes, combined with temporal and spatial gradients in [Ca2+] and [cAMP], allow a cell to respond to one or several signals with subtly nuanced metabolic changes, localized in space and time.