Download Molecular genetic techniques and more Lecture notes Remedial Biology in PDF only on Docsity!
Computer-generated model of a section of a mitochon- drion from chicken brain, based on a three-dimensional electron tomogram. [T. Frey and C. Mannella, 2000, Trends Biochem. Sci. 25: 319.]
CELLULAR
ENERGETICS
T
he most important molecule for capturing and trans- ferring free energy in biological systems is adenosine triphosphate, or ATP (see Figure 2-24). Cells use the energy released during hydrolysis of the terminal “high- energy” phosphoanhydride bond in ATP to power many energetically unfavorable processes. Examples include the synthesis of proteins from amino acids and of nucleic acids from nucleotides (Chapter 4), transport of molecules against a concentration gradient by ATP-powered pumps (Chapter 7), contraction of muscle (Chapter 19), and movement (beat- ing) of cilia (Chapter 20). Although other high-energy mole- cules occur in cells, ATP is the universal “currency” of chemical energy; it is found in all types of organisms and must have occurred in the earliest life-forms. This chapter focuses on how cells generate the high- energy phosphoanhydride bond of ATP from ADP and in- organic phosphate (HPO 42 �). This endergonic reaction, which is the reverse of ATP hydrolysis and requires an input of 7.3 kcal/mol to proceed, can be written as
Pi^2 �^ � H�^ � ADP 3 �^ ATP^4 �^ � H 2 O
where P (^) i^2 �^ represents inorganic phosphate (HPO 42 �). The energy to drive this reaction is produced primarily by two main processes: aerobic oxidation, which occurs in nearly all cells, and photosynthesis, which occurs only in leaf cells of plants and certain single-celled organisms. In aerobic oxidation, fatty acids and sugars, principally glucose, are metabolized to carbon dioxide (CO 2 ) and water (H 2 O), and the released energy is converted to the chemical energy of phosphoanhydride bonds in ATP. In animal cells and most other nonphotosynthetic cells, ATP is generated mainly by this process. The initial steps in the oxidation of
glucose, called glycolysis, occur in the cytosol in both eu- karyotes and prokaryotes and do not require oxygen (O 2 ). The final steps, which require oxygen, generate most of the ATP. In eukaryotes, these later stages of aerobic oxidation occur in mitochondria; in prokaryotes, which contain only a plasma membrane and lack internal organelles, many of the final steps occur on the plasma membrane. The final stages of fatty acid metabolism sometimes occur in mito- chondria and generate ATP; in most eukaryotic cells, how- ever, fatty acids are metabolized to CO 2 and H 2 O in peroxisomes without production of ATP. In photosynthesis, light energy is converted to the chem- ical energy of phosphoanhydride bonds in ATP and stored in the chemical bonds of carbohydrates (primarily sucrose and starch). Oxygen also is formed during photosynthesis. In
O U T L I N E
8.1 Oxidation of Glucose and Fatty Acids to CO 2
8.2 Electron Transport and Generation of
the Proton-Motive Force
8.3 Harnessing the Proton-Motive Force for Energy-
Requiring Processes
8.4 Photosynthetic Stages and Light-Absorbing
Pigments
8.5 Molecular Analysis of Photosystems
8.6 CO 2 Metabolism During Photosynthesis
plants and eukaryotic single-celled algae, photosynthesis oc- curs in chloroplasts. Several prokaryotes also carry out pho- tosynthesis on their plasma membrane or its invaginations by a mechanism similar to that in chloroplasts. The oxygen gen- erated during photosynthesis is the source of virtually all the oxygen in the air, and the carbohydrates produced are the ul- timate source of energy for virtually all nonphotosynthetic organisms. Bacteria living in deep ocean vents, where there is no sunlight, disprove the popular view that sunlight is the ul- timate source of energy for all organisms on earth. These bacteria obtain energy for converting carbon dioxide into carbohydrates and other cellular constituents by oxidation of reduced inorganic compounds in dissolved vent gas. At first glance, photosynthesis and aerobic oxidation ap- pear to have little in common. However, a revolutionary dis- covery in cell biology is that bacteria, mitochondria, and chloroplasts all use the same basic mechanism, called chemiosmosis (or chemiosmotic coupling), to generate ATP from ADP and P (^) i. In chemiosmosis, a proton (H�) concen- tration gradient and an electric potential (voltage gradient) across the membrane, collectively termed the proton-motive force, drive an energy-requiring process such as ATP synthe- sis (Figure 8-1, bottom ). Chemiosmosis can occur only in sealed, membrane- limited compartments that are impermeable to H�. The proton-motive force is generated by the stepwise movement of electrons from higher to lower energy states via membrane- bound electron carriers. In mitochondria and nonphotosyn- thetic bacterial cells, electrons from NADH (produced during the metabolism of sugars, fatty acids, and other sub- stances) are transferred to O 2 , the ultimate electron acceptor. In the thylakoid membrane of chloroplasts, energy absorbed from light strips electrons from water (forming O 2 ) and pow-
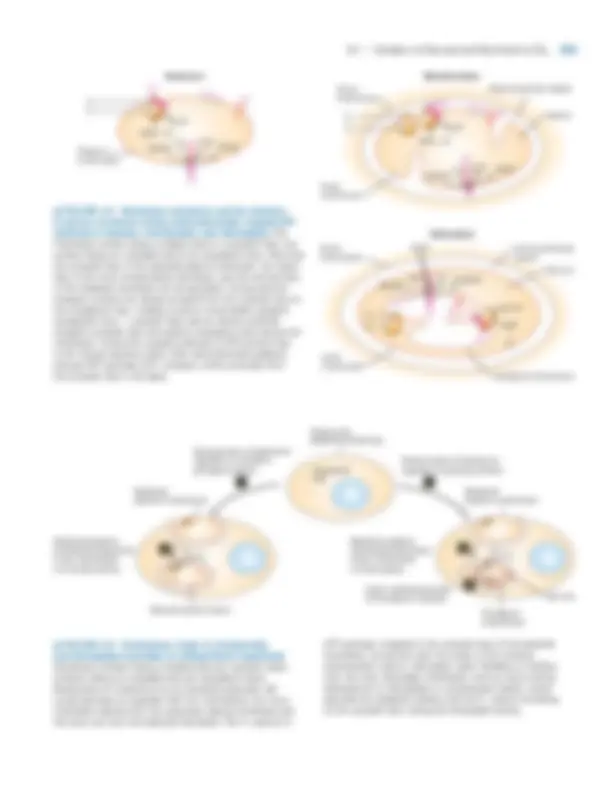
302 CHAPTER 8 •^ Cellular Energetics
ers their movement to other electron carriers, particularly NADP�; eventually these electrons are donated to CO 2 to synthesize carbohydrates. All these systems, however, con- tain some similar carriers that couple electron transport to the pumping of protons across the membrane—always from the cytosolic face to the exoplasmic face of the membrane— thereby generating the proton-motive force (Figure 8-1, top ). Invariably, the cytosolic face has a negative electric poten- tial relative to the exoplasmic face. Moreover, mitochondria, chloroplasts, and bacteria uti- lize essentially the same kind of membrane protein, the F 0 F 1 complex, to synthesize ATP. The F 0 F 1 complex, now com- monly called ATP synthase, is a member of the F class of ATP-powered proton pumps (see Figure 7-6). In all cases, ATP synthase is positioned with the globular F 1 domain, which catalyzes ATP synthesis, on the cytosolic face of the membrane, so ATP is always formed on the cytosolic face of the membrane (Figure 8-2). Protons always flow through ATP synthase from the exoplasmic to the cytosolic face of the membrane, driven by a combination of the proton concen- tration gradient ([H�]exoplasmic � [H�]cytosolic ) and the mem- brane electric potential (exoplasmic face positive with respect to the cytosolic face). These commonalities between mitochondria, chloro- plasts, and bacteria undoubtedly have an evolutionary origin. In bacteria both photosynthesis and oxidative phos- phorylation occur on the plasma membrane. Analysis of the sequences and transcription of mitochondrial and chloroplast DNAs (Chapters 10 and 11) has given rise to the popular hypothesis that these organelles arose early in the evolution of eukaryotic cells by endocytosis of bacteria capable of oxidative phosphorylation or photosynthesis, re- spectively (Figure 8-3). According to this endosymbiont
− (^) H +
X
H+^ symport H+^ H+
H+
H+^ antiport
Y
Membrane impermeable to H+
Cytosolic face
Exoplasmic face
Rotation of bacterial flagella
Transport of molecule (X or Y) against concentration gradient
Synthesis of ATP
Chemical energy Light energy
H+
ADP + P (^) i
ATP
H+
GENERATION OF PROTON-MOTIVE FORCE
Electric potential (^) H +^ concentration gradient
Sealed compartment
C
B
A
Mitochondria and aerobic bacteria
Chloroplasts and photosynthetic bacteria
CHEMIOSMOTIC COUPLING
+++ +++
- –– –––
� FIGURE 8-1^ Overview of the generation and utilization of a proton-motive force. A transmembrane proton concentration gradient and a voltage gradient, collectively called the proton-motive force, are generated during photo- synthesis and the aerobic oxidation of carbon compounds in mitochondria and aerobic bacteria. In chemiosmotic coupling, a proton-motive force powers an energy-requiring process such as ATP synthesis (A), transport of metabolites across the membrane against their concentration gradient (B), or rotation of bacterial flagella (C).
hypothesis, the inner mitochondrial membrane would be derived from the bacterial plasma membrane with the globular F 1 domain still on its cytosolic face pointing to- ward the matrix space of the mitochondrion. Similarly, the globular F 1 domain would be on the cytosolic face of the thylakoid membrane facing the stromal space of the chloroplast. In addition to powering ATP synthesis, the proton- motive force can supply energy for the transport of small molecules across a membrane against a concentration gra- dient (see Figure 8-1). For example, a H�/sugar symport protein catalyzes the uptake of lactose by certain bacteria, and proton-driven antiporters catalyze the accumulation of ions and sucrose by plant vacuoles (Chapter 7). The proton-motive force also powers the rotation of bacterial flagella. (The beating of eukaryotic cilia, however, is pow- ered by ATP hydrolysis.) Conversely, hydrolysis of ATP by V-class ATP-powered proton pumps, which are similar in structure to F-class pumps (see Figure 7-6), provides the en- ergy for transporting protons against a concentration gra- dient. Chemiosmotic coupling thus illustrates an important principle introduced in our discussion of active transport in Chapter 7: the membrane potential, the concentration gra- dients of protons (and other ions) across a membrane, and the phosphoanhydride bonds in ATP are equivalent and in- terconvertible forms of chemical potential energy. In this brief overview, we’ve seen that oxygen and carbohydrates are produced during photosynthesis, whereas they are consumed during aerobic oxidation. In both processes, the flow of electrons creates a H�^ electrochemical gradient, or proton-motive force, that can power ATP syn- thesis. As we examine these two processes at the molecular level, focusing first on aerobic oxidation and then on pho- tosynthesis, the striking parallels between them will become evident.
Oxidation of Glucose
and Fatty Acids to CO 2
The complete aerobic oxidation of each molecule of glucose yields 6 molecules of CO 2 and is coupled to the synthesis of as many as 30 molecules of ATP:
C 6 H 12 O 6 � 6 O 2 � 30 P (^) i^2 �^ � 30 ADP^3 �^ � 30 H� 6 CO 2 � 30 ATP^4 �^ � 36 H 2 O
Glycolysis, the initial stage of glucose metabolism, takes place in the cytosol and does not involve molecular O 2. It produces a small amount of ATP and the three-carbon compound pyruvate. In aerobic cells, pyruvate formed in glycolysis is transported into the mitochondria, where it is oxidized by O 2 to CO 2. Via chemiosmotic coupling, the oxidation of pyruvate in the mitochondria generates the
8.
304 CHAPTER 8 •^ Cellular Energetics
bulk of the ATP produced during the conversion of glu- cose to CO 2. In this section, we discuss the biochemical pathways that oxidize glucose and fatty acids to CO 2 and H 2 O; the fate of the released electrons is described in the next section.
Cytosolic Enzymes Convert Glucose to Pyruvate During Glycolysis A set of 10 water-soluble cytosolic enzymes catalyze the re- actions constituting the glycolytic pathway, in which one molecule of glucose is converted to two molecules of pyru- vate (Figure 8-4). All the metabolic intermediates between glucose and pyruvate are water-soluble phosphorylated compounds. Four molecules of ATP are formed from ADP during glycolysis via substrate-level phosphorylation, which is catalyzed by enzymes in the cytosol (reactions 7 and 10). Unlike ATP formation in mitochondria and chloroplasts, a proton-motive force is not involved in substrate-level phosphorylation. Early in the glycolytic pathway, two ATP molecules are consumed: one by the addition of a phos- phate residue to glucose in the reaction catalyzed by hex- okinase (reaction 1), and another by the addition of a second phosphate to fructose 6-phosphate in the reaction catalyzed by phosphofructokinase-1 (reaction 3). Thus glycolysis yields a net of only two ATP molecules per glu- cose molecule. The balanced chemical equation for the conversion of glucose to pyruvate shows that four hydrogen atoms (four protons and four electrons) are also formed:
(For convenience, we show pyruvate here in its un-ionized form, pyruvic acid, although at physiological pH it would be largely dissociated.) All four electrons and two of the four protons are transferred to two molecules of the oxidized form of the electron carrier nicotinamide adenine dinu- cleotide ( NAD �) to produce the reduced form, NADH (see Figure 2-26):
2 H�^ � 4 e�^ � 2 NAD�^ 2 NADH
The reaction that generates these hydrogen atoms and transfers them to NAD�^ is catalyzed by glyceraldehyde 3-phosphate dehydrogenase (see Figure 8-4, reaction 6). The overall chemical equation for this first stage of glucose metabolism is
C 6 H 12 O 6 � 2 NAD�^ � 2 ADP^3 �^ � 2 P (^) i^2 � ¡ 2 C 3 H 4 O 3 � 2 NADH � 2 ATP^4 �
Glucose Pyruvate
C 6 H 12 O 6 2 CH 3 C OH
O O C � 4 H��4 e�
Anaerobic Metabolism of Each Glucose Molecule
Yields Only Two ATP Molecules
Many eukaryotes are obligate aerobes: they grow only in the presence of oxygen and metabolize glucose (or related sug- ars) completely to CO 2 , with the concomitant production of a large amount of ATP. Most eukaryotes, however, can gen- erate some ATP by anaerobic metabolism. A few eukaryotes are facultative anaerobes: they grow in either the presence or the absence of oxygen. For example, annelids, mollusks, and some yeasts can live and grow for days without oxygen. In the absence of oxygen, facultative anaerobes convert glucose to one or more two- or three-carbon compounds, which are generally released into the surrounding medium. For instance, yeasts degrade glucose to two pyruvate mole- cules via glycolysis, generating a net of two ATP. In this process two NADH molecules are formed from NAD�^ per glucose molecule. In the absence of oxygen, yeasts convert pyruvate to one molecule each of ethanol and CO 2 ; in these
8.1 •^ Oxidation of Glucose and Fatty Acids to CO 2 305
reactions two NADH molecules are oxidized to NAD�^ for each two pyruvates converted to ethanol, thereby regenerat- ing the supply of NAD�^ (Figure 8-5a, left ). This anaerobic degradation of glucose, called fermentation, is the basis of beer and wine production. During the prolonged contraction of mammalian skeletal muscle cells, when oxygen becomes limited, muscle cells
CH 2 OH H
H
OH
OH
H H H
O
HO OH
H
H
OH
OH
H H H
O
HO OH
CH 2 OPO 32 �
CH 2 OH
CH 2 OPO 32 �
H
HO
OH
H
O
H OH
CH 2 OPO 32 � CH 2 OPO 32 �
H
HO
OH
H
O
H OH
Glucose
Glucose 6-phosphate
Fructose 6-phosphate
Dihydroxyacetone phosphate
Fructose 1,6-bisphosphate
1,3-Bisphosphoglycerate (2 molecules)
3-Phosphoglycerate (2 molecules)
2-Phosphoglycerate (2 molecules)
Phosphoenolpyruvate (2 molecules)
Pyruvate (2 molecules)
Hexokinase
Phosphoglucose isomerase
Phosphofructo- kinase-
Aldolase
Triose phosphate isomerase Glyceraldehyde 3-phosphate (2 molecules )
ATP ADP
Glyceraldehyde 3-phosphate dehydrogenase
2 NAD�� 2 Pi 2 NADH � 2 H�
ATP ADP
Phosphoglycerate kinase
Phosphoglycero- mutase
Enolase
Pyruvate kinase
2 ADP 2 ATP
2 ADP 2 ATP
2 H 2 O
1 2 3 4 5 6 7 8 9
10
H O OH C C H H
�HO 3 PO C
O H H HC C C H OHOPO 32 �
O H H
OH
C C C H OPO 32 �
2 �O 3 PO
O H H
OH
�OC C C H OPO 32 �
� O
O H
HO
C C H
H C 2 �O 3 PO
�O
O H C C C H 2 �O 3 PO
�O
O H
H
C C C H
O
� FIGURE 8-4^ The glycolytic pathway by which glucose is degraded to pyruvic acid. Two reactions consume ATP, forming ADP and phosphorylated sugars (red); two generate ATP from ADP by substrate-level phosphorylation (green); and one yields NADH by reduction of NAD�^ (yellow). Note that all the intermedi- ates between glucose and pyruvate are phosphorylated com- pounds. Reactions 1, 3, and 10, with single arrows, are essentially irreversible (large negative ∆ G values) under conditions ordinarily obtaining in cells.
ferment glucose to two molecules of lactic acid—again, with the net production of only two molecules of ATP per glucose molecule (Figure 8-5a, right ). The lactic acid causes muscle and joint aches. It is largely secreted into the blood; some passes into the liver, where it is reoxidized to pyruvate and ei- ther further metabolized to CO 2 aerobically or converted to glucose. Much lactate is metabolized to CO 2 by the heart, which is highly perfused by blood and can continue aerobic metabolism at times when exercising skeletal muscles secrete lactate. Lactic acid bacteria (the organisms that “spoil” milk) and other prokaryotes also generate ATP by the fermentation of glucose to lactate. In the presence of oxygen, however, pyruvate formed in glycolysis is transported into mitochondria, where it is oxi- dized by O 2 to CO 2 in a series of oxidation reactions collec- tively termed cellular respiration (Figure 8-5b). These reactions generate an estimated 28 additional ATP molecules per glucose molecule, far outstripping the ATP yield from anaerobic glucose metabolism. To understand how mito- chondria operate as ATP-generating factories, we first de- scribe their structure and then the reactions they employ to degrade pyruvate.
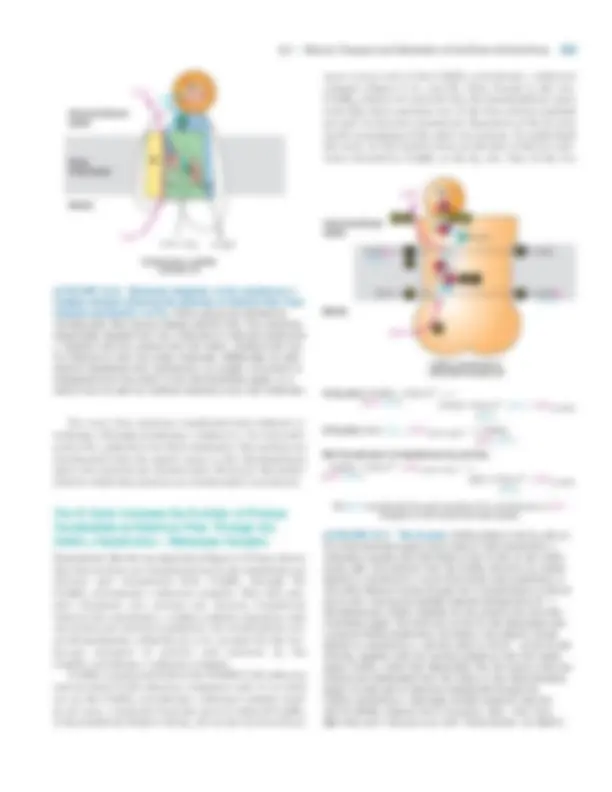
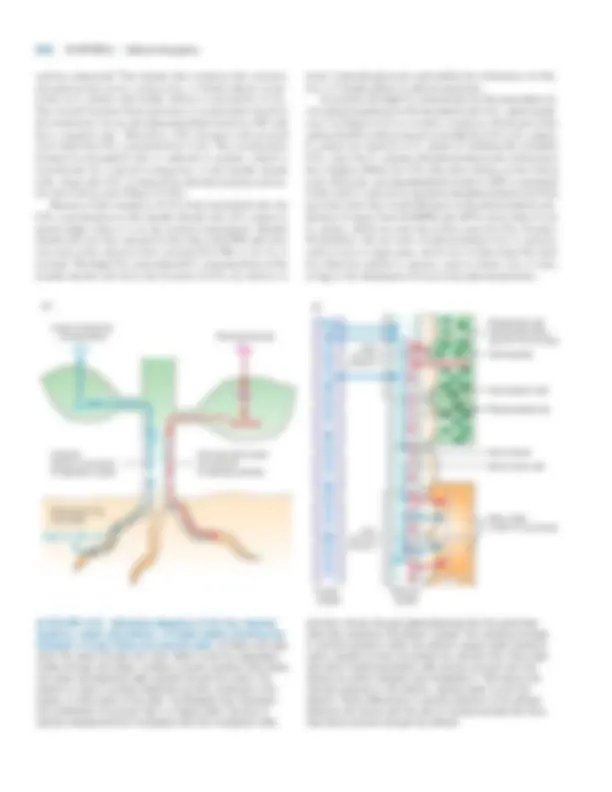
Mitochondria Possess Two Structurally and Functionally Distinct Membranes
Mitochondria are among the larger organelles in the cell, each one being about the size of an E. coli bacterium. Most eukaryotic cells contain many mitochondria, which collec- tively can occupy as much as 25 percent of the volume of the cytoplasm. They are large enough to be seen under a light microscope, but the details of their structure can be viewed only with the electron microscope (see Figure 5-26). The outer membrane defines the smooth outer perimeter of the mitochondrion. In contrast, the inner membrane has numer- ous invaginations called cristae. These membranes define two submitochondrial compartments: the intermembrane space between the outer membrane and the inner membrane with its cristae, and the matrix, or central compartment (Figure 8-6). The fractionation and purification of these membranes and compartments have made it possible to determine their protein and phospholipid compositions and to localize each enzyme-catalyzed reaction to a specific membrane or space. The outer membrane contains mitochondrial porin, a transmembrane channel protein similar in structure to
8.1 •^ Oxidation of Glucose and Fatty Acids to CO 2 307
(b)
▲ FIGURE 8-6 Internal structure of a mitochondrion. (a) Schematic diagram showing the principal membranes and compartments. The cristae form sheets and tubes by invagination of the inner membrane and connect to the inner membrane through relatively small uniform tubular structures called crista junctions. The intermembrane space appears continuous with the lumen of each crista. The F 0 F 1 complexes (small red spheres), which synthesize ATP, are intramembrane particles that protrude from the cristae and inner membrane into the matrix. The matrix contains the mitochondrial DNA (blue strand), ribosomes (small blue spheres), and granules (large yellow spheres).
(b) Computer-generated model of a section of a mitochondrion from chicken brain. This model is based on a three-dimensional electron tomogram calculated from a series of two-dimensional electron micrographs recorded at regular angular intervals. This technique is analogous to a three-dimensional X-ray tomogram or CAT scan. Note the tightly packed cristae (yellow-green), the inner membrane (light blue), and the outer membrane (dark blue). [Part (a) courtesy of T. Frey; part (b) from T. Frey and C. Mannella, 2000, Trends Biochem. Sci. 25 :319.]
M E D I A
C O N N E C T I O N S
Video: Mitochondrion Reconstructed by Electron Tomography
Intermembrane space Outer membrane Inner membrane Matrix
Cristae junctions
Cristae
1~2 �m
0.1~0.5 �m
(a)
bacterial porins (see Figure 5-14). Ions and most small mole- cules (up to about 5000 Da) can readily pass through these channel proteins. Although the flow of metabolites across the outer membrane may limit their rate of mitochondrial oxida- tion, the inner membrane and cristae are the major permeabil- ity barriers between the cytosol and the mitochondrial matrix. Freeze-fracture studies indicate that mitochondrial cristae contain many protein-rich intramembrane particles. Some are the F 0 F 1 complexes that synthesize ATP; others function in transporting electrons to O 2 from NADH or other elec-
308 CHAPTER 8 •^ Cellular Energetics
tron carriers. Various transport proteins located in the inner membrane and cristae allow otherwise impermeable mole- cules, such as ADP and Pi, to pass from the cytosol to the ma- trix, and other molecules, such as ATP, to move from the matrix into the cytosol. Protein constitutes 76 percent of the total weight of the inner membrane—a higher fraction than in any other cellular membrane. Cardiolipin (diphosphatidyl glycerol), a lipid concentrated in the inner membrane, suffi- ciently reduces the membrane’s permeability to protons that a proton-motive force can be established across it.
1 Pyruvate dehydrogenase, citric acid cycle, and fatty acid metabolism Electron transport from NADH and FADH 2 to oxygen; generation of proton-motive force (^3) ATP synthesis by F 0 F 1 using proton-motive force
2
1
2 3
Pyruvate
Fatty acid
Fatty acyl CoA
Acetyl CoA
Fatty acyl CoA
NADH
NADH
NADH
ATP HSCoA
HSCoA
HSCoA CH 3 C C OH CH 3 C SCoA
�
AMP PP (^) i
ATP ATP
GDP GTP
ADP + ADP
�
P (^) i
Pi
Electron shuttle
NAD+
H+^ H+^ H+^ 3 H+
3 H+
OH−
Transporters
NAD+
NAD+^ NADH 3 NAD+^ 3 NADH
NAD+^ NAD
Electron transport chain
H 2 O
CO 2
O (^2) H 2 O
2 CO 2
CO 2
CO 2
F 0 F 1 complex
2 e−^ 2 H+ O 2
FADH 2
FAD FADH 2
FAD
1 2
HSCoA
Transporter
Transporter
O O O
Citric acid cycle
Intermembrane space
Inner mitochondrial membrane and cristae
Pi
Mitochondrial matrix
Outer mitochondrial membrane (permeable to metabolites)
I III IV
Succinate
FAD II
Fumarate
▲ FIGURE 8-7 Summary of the aerobic oxidation of pyruvate and fatty acids in mitochondria. The outer membrane is freely permeable to all metabolites, but specific transport proteins (colored ovals) in the inner membrane are required to import pyruvate (yellow), ADP (green), and Pi (purple) into the matrix and to export ATP (green). NADH generated in the cytosol is not transported directly to the matrix because the inner membrane is impermeable to NAD�^ and NADH; instead, a shuttle system (red) transports electrons from cytosolic NADH to NAD� in the matrix. O 2 diffuses into the matrix and CO 2 diffuses out. Stage 1: Fatty acyl groups are transferred from fatty acyl CoA and transported across the inner membrane via a special carrier (blue oval) and then reattached to CoA on the matrix side.
Pyruvate is converted to acetyl CoA with the formation of NADH, and fatty acids (attached to CoA) are also converted to acetyl CoA with formation of NADH and FADH. Oxidation of acetyl CoA in the citric acid cycle generates NADH and FADH 2. Stage 2: Electrons from these reduced coenzymes are transferred via electron-transport complexes (blue boxes) to O 2 concomitant with transport of H�^ ions from the matrix to the intermembrane space, generating the proton-motive force. Electrons from NADH flow directly from complex I to complex III, bypassing complex II. Stage 3: ATP synthase, the F 0 F 1 complex (orange), harnesses the proton-motive force to synthesize ATP. Blue arrows indicate electron flow; red arrows transmembrane movement of protons; and green arrows transport of metabolites.
to FAD, yielding the reduced form of this coenzyme, FADH 2. In reaction 6, hydrolysis of the high-energy thioester bond in succinyl CoA is coupled to synthesis of one GTP by substrate- level phosphorylation (GTP and ATP are interconvertible). Reaction 9, the final one, also regenerates oxaloacetate, so the cycle can begin again. Note that molecular O 2 does not par- ticipate in the citric acid cycle. Most enzymes and small molecules involved in the citric acid cycle are soluble in aqueous solution and are localized to the mitochondrial matrix. These include CoA, acetyl CoA, suc- cinyl CoA, NAD�, and NADH, as well as six of the eight cycle enzymes. Succinate dehydrogenase, (reaction 7) and �-ketoglu- tarate dehydrogenase (reaction 5) are integral proteins in the inner membrane, with their active sites facing the matrix. When mitochondria are disrupted by gentle ultrasonic vibration or os- motic lysis, the six non-membrane-bound enzymes in the citric acid cycle are released as a very large multiprotein complex. The reaction product of one enzyme is thought to pass directly to the next enzyme without diffusing through the solution. However, much work is needed to determine the structure of this enzyme complex as it exists in the cell.
310 CHAPTER 8 •^ Cellular Energetics
Since glycolysis of one glucose molecule generates two acetyl CoA molecules, the reactions in the glycolytic pathway and citric acid cycle produce six CO 2 molecules, ten NADH molecules, and two FADH 2 molecules per glucose molecule (Table 8-1). Although these reactions also generate four high- energy phosphoanhydride bonds in the form of two ATP and two GTP molecules, this represents only a small fraction of the available energy released in the complete aerobic oxida- tion of glucose. The remaining energy is stored in the re- duced coenzymes NADH and FADH 2. Synthesis of most of the ATP generated in aerobic oxi- dation is coupled to the reoxidation of NADH and FADH (^2) by O 2 in a stepwise process involving the respiratory chain, also called the electron transport chain. Even though mo- lecular O 2 is not involved in any reaction of the citric acid cycle, in the absence of O 2 the cycle soon stops operating as the supply of NAD�^ and FAD dwindles. Before consid- ering electron transport and the coupled formation of ATP in detail, we discuss first how the supply of NAD�^ in the cytosol is regenerated and then the oxidation of fatty acids to CO 2.
CH 2
HO C
COO�
COO�
Citrate
CH 2
COO�
HC
C COO�
COO� cis-Aconitate
CH 2
COO�
C C
H HO H
COO�
COO� Isocitrate
CH 2
COO�
CH 2 COO� Succinate
CH 2
COO� COO�
HC
Fumarate
CH
COO�
CH 2 COO�
COO� C O
CH^ Oxaloacetate 2 COO�
COO� HO C H
Malate
C O
CH 2
COO� -Keto- glutarate
�
CH 2
COO�
CH 3 C SCoA HSCoA
O
Acetyl CoA
� H 2 O
1
3
2
4 6 5
7
8
9
H 2 O
H 2 O
NAD�
�^ CO^2 �^ NADH�^ H�
NAD� HSCoA CO 2 � (^) NADH �H�
GDP � Pi � H 2 O GTP �HSCoA
FADH 2
FAD
H 2 O
� H�
NAD�
NADH
CH 2
CH 2
C SCoA O
COO�
Succinyl CoA
▲ FIGURE 8-9 The citric acid cycle, in which acetyl groups transferred from acetyl CoA are oxidized to CO 2. In reaction 1, a two-carbon acetyl residue from acetyl CoA condenses with the four-carbon molecule oxaloacetate to form the six-carbon molecule citrate. In the remaining reactions (2–9) each molecule of citrate is eventually converted back to oxaloacetate, losing two CO 2 molecules in the process. In each turn of the cycle, four pairs of electrons are removed from carbon atoms, forming
three molecules of NADH and one molecule of FADH 2. The two carbon atoms that enter the cycle with acetyl CoA are highlighted in blue through succinyl CoA. In succinate and fumarate, which are symmetric molecules, they can no longer be specifically denoted. Isotope labeling studies have shown that these carbon atoms are not lost in the turn of the cycle in which they enter; on average one will be lost as CO 2 during the next turn of the cycle and the other in subsequent turns.
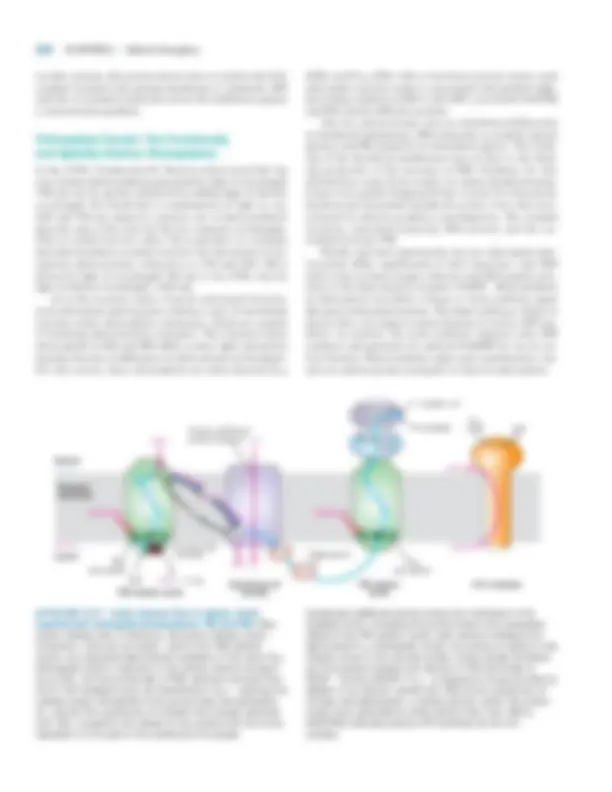
Transporters in the Inner Mitochondrial
Membrane Allow the Uptake of Electrons
from Cytosolic NADH
For aerobic oxidation to continue, the NADH produced dur- ing glycolysis in the cytosol must be oxidized to NAD�. As with NADH generated in the mitochondrial matrix, elec- trons from cytosolic NADH are ultimately transferred to O 2 via the respiratory chain, concomitant with the generation of
8.1 •^ Oxidation of Glucose and Fatty Acids to CO 2 311
a proton-motive force. Although the inner mitochondrial membrane is impermeable to NADH itself, several electron shuttles can transfer electrons from cytosolic NADH to the matrix. Operation of the most widespread shuttle—the malate- aspartate shuttle —is depicted in Figure 8-10. Critical to the shuttle are two antiport proteins in the inner mitochondrial membrane, a malate/ � -ketoglutarate antiporter and a glutamate/aspartate antiporter, that permit transport of their
TABLE 8-1 Net Result of the Glycolytic Pathway and the Citric Acid Cycle
CO 2 Molecules NAD �^ Molecules FAD Molecules Reaction Produced Reduced to NADH Reduced to FADH 2
1 glucose molecule to 2 0 2 0 pyruvate molecules
2 pyruvates to 2 acetyl CoA 2 2 0 molecules
2 acetyl CoA to 4 CO 2 4 6 2 molecules
Total 6 10 2
Malate
Aspartate Malate
Aspartate
Oxaloacetate
NADHcytosol NAD� cytosol
Malate dehydrogenase
Malate dehydrogenase
NADHmatrix NAD� matrix
Oxaloacetate
-Ketoglutarate Glutamate
Transaminase
Transaminase
-KetoglutarateGlutamate
Cytosol
Mitochondrial inner membrane
Matrix
5
6
1
2
3
4
▲ FIGURE 8-10 The malate shuttle. This cyclical series of reactions transfers electrons from NADH in the cytosol (intermembrane space) across the inner mitochondrial membrane, which is impermeable to NADH itself. Step 1 : Cytosolic malate dehydrogenase transfers electrons from cytosolic NADH to oxaloacetate, forming malate. Step 2 : An antiporter (blue oval) in the inner mitochondrial membrane transports malate into the matrix in exchange for -ketoglutarate. Step 3 : Mitochondrial malate dehydrogenase converts malate back to oxaloacetate, reducing NAD�^ in the matrix to NADH in the process. Step 4 : Oxaloacetate, which cannot directly cross the inner membrane, is converted to
aspartate by addition of an amino group from glutamate. In this transaminase-catalyzed reaction in the matrix, glutamate is converted to -ketoglutarate. Step 5 : A second antiporter (red oval) exports aspartate to the cytosol in exchange for glutamate. Step 6 : A cytosolic transaminase converts aspartate to oxaloacetate, completing the cycle. The blue and red arrows reflect the movement of the -ketoglutarate and glutamate, respectively. In step 4 glutamate is deaminated to -ketoglutarate, which is transported to the cytosol by an antiporter (step 2 ; in step 6 , the -ketoglutarate is aminated, converting it back to glutamate, which is transported to the matrix by the antiporter in step 5.
1 g of triacylglycerol to CO 2 generates about six times as much ATP as does the oxidation of 1 g of hydrated glycogen, the polymeric storage form of glucose in muscle and liver. Triglycerides are more efficient for storage of energy because they are stored in anhydrous form and are much more re- duced (have more hydrogens) than carbohydrates and there- fore yield more energy when oxidized. In the cytosol, free fatty acids are esterified to coenzyme A to form a fatty acyl CoA in an exergonic reaction coupled to the hydrolysis of ATP to AMP and PP (^) i (inorganic pyrophosphate):
Subsequent hydrolysis of PPi to two molecules of phosphate (P (^) i ) drives this reaction to completion. Then the fatty acyl group is transferred to carnitine and moved across the inner mitochondrial membrane by an acylcarnitine transporter protein (see Figure 8-7, blue oval); on the matrix side, the fatty acyl group is released from carnitine and reattached to another CoA molecule. Each molecule of a fatty acyl CoA in the mitochondrion is oxidized in a cyclical sequence of four reactions in which all the carbon atoms are converted to acetyl CoA with generation of NADH and FADH 2 (Figure 8-11a). For example, mitochon- drial oxidation of each molecule of the 18-carbon stearic acid, CH 3 (CH 2 ) 16 COOH, yields nine molecules of acetyl CoA and eight molecules each of NADH and FADH 2. As with acetyl CoA generated from pyruvate, these acetyl groups enter the cit- ric acid cycle and are oxidized to CO 2. Electrons from the re- duced coenzymes produced in the oxidation of fatty acyl CoA to acetyl CoA and in the subsequent oxidation of acetyl CoA in the citric acid cycle move via the respiratory chain to O 2. This electron movement is coupled to generation of a proton-motive force that is used to power ATP synthesis as described previ- ously for the oxidation of pyruvate (see Figure 8-7).
Peroxisomal Oxidation of Fatty
Acids Generates No ATP
Mitochondrial oxidation of fatty acids is the major source of ATP in mammalian liver cells, and biochemists at one time believed this was true in all cell types. However, rats treated with clofibrate, a drug used to reduce the level of blood lipoproteins, were found to exhibit an increased rate of fatty acid oxidation and a large increase in the number of peroxi- somes in their liver cells. This finding suggested that peroxi- somes, as well as mitochondria, can oxidize fatty acids. These small organelles, ≈0.2–1 μm in diameter, are lined by a single membrane (see Figure 5-21). They are present in all
8.1 •^ Oxidation of Glucose and Fatty Acids to CO 2 313
mammalian cells except erythrocytes and are also found in plant cells, yeasts, and probably most other eukaryotic cells. The peroxisome is now recognized as the principal or- ganelle in which fatty acids are oxidized in most cell types. Indeed, very long chain fatty acids containing more than about 20 CH 2 groups are degraded only in peroxisomes; in mammalian cells, mid-length fatty acids containing 10– CH 2 groups can be degraded in both peroxisomes and mi- tochondria. In contrast to mitochondrial oxidation of fatty acids, which is coupled to generation of ATP, peroxisomal oxidation of fatty acids is not linked to ATP formation, and the released energy is converted to heat. The reaction pathway by which fatty acids are degraded to acetyl CoA in peroxisomes is similar to that used in liver mitochondria (Figure 8-11b). However, peroxisomes lack a respiratory chain, and electrons from the FADH 2 produced during the oxidation of fatty acids are immediately trans- ferred to O 2 by oxidases , regenerating FAD and forming hydrogen peroxide (H 2 O 2 ). In addition to oxidases, peroxisomes contain abundant catalase , which quickly de- composes the H 2 O 2 , a highly cytotoxic metabolite. NADH produced during oxidation of fatty acids is exported and re- oxidized in the cytosol. Peroxisomes also lack the citric acid cycle, so acetyl CoA generated during peroxisomal degrada- tion of fatty acids cannot be oxidized further; instead it is transported into the cytosol for use in the synthesis of cho- lesterol and other metabolites.
Before fatty acids can be degraded in the peroxi- some, they must first be transported into the or- ganelle from the cytosol. Mid-length fatty acids are esterified to coenzyme A in the cytosol; the resulting fatty acyl CoAs are then transported into the peroxisome by a spe- cific transporter. However, very long chain fatty acids enter the peroxisome by another transporter and then are esteri- fied to CoA once inside. In the human genetic disease X-linked adrenoleukodystrophy (ALD), peroxisomal oxidation of very long chain fatty acids is specifically defective, while the oxidation of mid-length fatty acids is normal. The most common peroxisomal disorder, ALD is marked by elevated levels of very long chain fatty acids in the plasma and tissues. Patients with the most severe form of ALD are unaffected until mid-childhood, when severe neurological disorders ap- pear, followed by death within a few years. In recent years, the gene that is defective in ALD patients has been identified and cloned by techniques described in Chapter 9. Sequence analysis shows that the gene encodes an ABC transport pro- tein (ABCD1) that is localized to peroxisomal membranes and is thought to mediate the import of very long chain fatty acids into the organelle. ❚
The Rate of Glucose Oxidation Is Adjusted to Meet the Cell’s Need for ATP All enzyme-catalyzed reactions and metabolic pathways are regulated by cells so as to produce the needed amounts of
PPi
Fatty acid
O R C �^ HSCoA �ATP
� AMP �
O�
Fatty acyl CoA
O R C SCoA
metabolites but not an excess. The primary function of the oxidation of glucose to CO 2 via the glycolytic pathway, the pyruvate dehydrogenase reaction, and the citric acid cycle is to produce NADH and FADH 2 , whose oxidation in mito- chondria generates ATP. Operation of the glycolytic pathway and citric acid cycle is continuously regulated, primarily by allosteric mechanisms, to meet the cell’s need for ATP (see Chapter 3 for general principles of allosteric control). Three allosterically controlled glycolytic enzymes play a key role in regulating the entire glycolytic pathway (see Fig- ure 8-4). Hexokinase (step 1 ) is inhibited by its reaction product, glucose 6-phosphate. Pyruvate kinase (step 10) is inhibited by ATP, so glycolysis slows down if too much ATP is present. The third enzyme, phosphofructokinase-1, which converts fructose 6-phosphate to fructose 1,6-bisphosphate (step 3 ), is the principal rate-limiting enzyme of the glycolytic pathway. Emblematic of its critical role in regulating the rate of glycolysis, this enzyme is allosterically controlled by sev- eral molecules (Figure 8-12). Phosphofructokinase-1 is allosterically inhibited by ATP and allosterically activated by AMP. As a result, the rate of glycolysis is very sensitive to the cell’s energy charge, re- flected in the ATP:AMP ratio. The allosteric inhibition of phosphofructokinase-1 by ATP may seem unusual, since ATP is also a substrate of this enzyme. But the affinity of the substrate-binding site for ATP is much higher (has a lower K m ) than that of the allosteric site. Thus at low concentra- tions, ATP binds to the catalytic but not to the inhibitory al- losteric site, and enzymatic catalysis proceeds at near maximal rates. At high concentrations, ATP also binds to the allosteric site, inducing a conformational change that reduces the affinity of the enzyme for the other substrate, fructose 6-phosphate, and thus inhibits the rate of this reaction and the overall rate of glycolysis.
314 CHAPTER 8 •^ Cellular Energetics
Another important allosteric activator of phosphofructo- kinase-1 is fructose 2,6-bisphosphate. This metabolite is formed from fructose 6-phosphate by phosphofructokinase- 2 , an enzyme different from phosphofructokinase-1. Fruc- tose 6-phosphate accelerates the formation of fructose 2,6-bisphosphate, which, in turn, activates phosphofructo- kinase-1. This type of control, by analogy with feedback con- trol, is known as feed-forward activation, in which the abundance of a metabolite (here, fructose 6-phosphate) induces an acceleration in its metabolism. Fructose 2,6- bisphosphate allosterically activates phosphofructokinase- in liver cells by decreasing the inhibitory effect of high ATP and by increasing the affinity of phosphofructokinase-1 for one of its substrates, fructose 6-phosphate. The three glycolytic enzymes that are regulated by al- losteric molecules catalyze reactions with large negative � G �� values—reactions that are essentially irreversible under or- dinary conditions. These enzymes thus are particularly suit- able for regulating the entire glycolytic pathway. Additional control is exerted by glyceraldehyde 3-phosphate dehydro- genase, which catalyzes the reduction of NAD�^ to NADH (see Figure 8-4, step 6). If cytosolic NADH builds up owing to a slowdown in mitochondrial oxidation, this reaction will be slowed by mass action. As we discuss later, mitochon- drial oxidation of NADH and FADH 2 , produced in the glycolytic pathway and citric acid cycle, also is tightly controlled to produce the appropriate amount of ATP re- quired by the cell. Glucose metabolism is controlled differently in various mammalian tissues to meet the metabolic needs of the organ- ism as a whole. During periods of carbohydrate starvation, for instance, glycogen in the liver is converted directly to glucose 6-phosphate (without involvement of hexokinase). Under these conditions, there is a reduction in fructose 2,6-bisphos-
Glucose
High [ATP] High [AMP]
To pyruvate
Fructose 6-phosphate
Fructose 1,6-bisphosphate
Phosphofructo- kinase-
Fructose 2,6-bisphosphate
Phospho- fructokinase-
ATP (^) ADP ATP
ADP
High [citrate]
Insulin
- -
+
+
+
▲ FIGURE 8-12 Allosteric control of glucose metabolism in the cytosol at the level of fructose 6-phosphate. The key regulatory enzyme in glycolysis, phosphofructokinase-1, is allosterically activated by AMP and fructose 2,6-bisphosphate, which are elevated when the cell’s energy stores are low. The enzyme is inhibited by ATP and citrate, which are elevated when the cell is actively oxidizing glucose to CO 2. Phosphofructokinase- (PFK2) is a bifunctional enzyme: its kinase activity forms fructose
2,6-bisphosphate from fructose 6-phosphate, and its phosphatase activity catalyzes the reverse reaction. Insulin, which is released by the pancreas when blood glucose levels are high, promotes PFK2 kinase activity and thus stimulates glycolysis. At low blood glucose, glucagon is released by the pancreas and promotes PFK2 phosphatase activity in the liver, indirectly slowing down glycolysis. We describe the role of insulin and glucagon in the integrated control of blood glucose levels in Chapter 15.
results from the uphill pumping of H�^ outward from the ma- trix, which becomes negative with respect to the intermem- brane space. Thus free energy released during the oxidation of NADH or FADH 2 is stored both as an electric potential and a proton concentration gradient—collectively, the proton-motive force—across the inner membrane. The movement of protons back across the inner membrane, driven by this force, is coupled to the synthesis of ATP from ADP and Pi by ATP synthase (see Figure 8-7). The synthesis of ATP from ADP and P (^) i , driven by the transfer of electrons from NADH or FADH 2 to O 2 , is the major source of ATP in aerobic nonphotosynthetic cells. Much evidence shows that in mitochondria and bacteria this
316 CHAPTER 8 •^ Cellular Energetics
process, called oxidative phosphorylation, depends on gen- eration of a proton-motive force across the inner membrane, with electron transport, proton pumping, and ATP forma- tion occurring simultaneously. In the laboratory, for instance, addition of O 2 and an oxidizable substrate such as pyruvate or succinate to isolated intact mitochondria results in a net synthesis of ATP if the inner mitochondrial membrane is in- tact. In the presence of minute amounts of detergents that make the membrane leaky, electron transport and the oxi- dation of these metabolites by O 2 still occurs, but no ATP is made. Under these conditions, no transmembrane proton concentration gradient or membrane electric potential can be maintained.
0
200
400
600
800
50
60
40
30
20
10
0
2 e−
2 e−^2 e−
Fumarate + 2 H+
FMN
NADH NAD+^ + H+
FAD + 2 H+
FAD
Succinate FADH (^2)
CoQ
NADH-CoQ reductase (complex I)
Succinate-CoQ reductase (complex II)
CoQH 2 –cytochromec reductase (complex III)
Cytochromec oxidase (complex IV)
Cytc (^1)
Cytb
Cyta
Cu (^) a
Cu (^) b
2 e−
Cytc
Cyta (^3)
Fe-S Fe-S
− 400
− 200
Fe-S
(^1) / 2 O 2 + 2H+^ H 2 O
H+in H +out
H +in
H +out
H +out
Redox potential (mV)
Free energy (kcal/mol)
H +in
▲ FIGURE 8-13 Changes in redox potential and free energy during stepwise flow of electrons through the respiratory chain. Blue arrows indicate electron flow; red arrows, translocation of protons across the inner mitochondrial membrane. Four large multiprotein complexes located in the inner membrane contain several electron-carrying prosthetic groups. Coenzyme Q (CoQ) and cytochrome c transport
electrons between the complexes. Electrons pass through the multiprotein complexes from those at a lower reduction potential to those with a higher (more positive) potential (left scale), with a corresponding reduction in free energy (right scale). The energy released as electrons flow through three of the complexes is sufficient to power the pumping of H�^ ions across the membrane, establishing a proton-motive force.
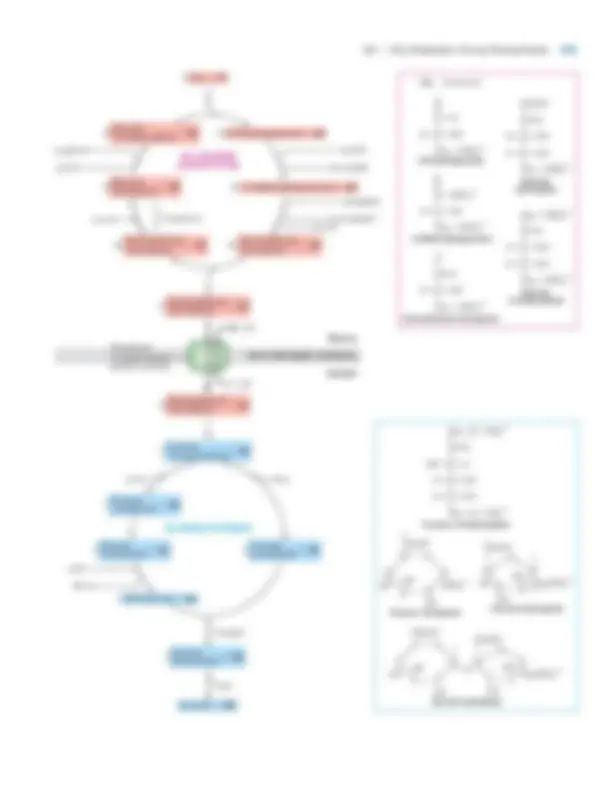
In this section we first discuss the magnitude of the proton-motive force, then the components of the respiratory chain and the pumping of protons across the inner mem- brane. In the following section we describe the structure of the ATP synthase and how it uses the proton-motive force to synthesize ATP. We also consider how mitochondrial oxi- dation of NADH and FADH 2 is controlled to meet the cell’s need for ATP.
The Proton-Motive Force in Mitochondria Is Due
Largely to a Voltage Gradient Across the Inner
Membrane
As we’ve seen, the proton-motive force (pmf) is the sum of a transmembrane proton concentration (pH) gradient and electric potential, or voltage gradient. The relative contribu- tion of the two components to the total pmf depends on the permeability of the membrane to ions other than H�. A sig- nificant voltage gradient can develop only if the membrane is poorly permeable to other cations and to anions, as is the inner mitochondrial membrane. In this case, the developing voltage gradient (i.e., excess H�^ ions on the intermembrane face and excess anions on the matrix face) soon prevents fur- ther proton movement, so only a small pH gradient is gen- erated. In contrast, a significant pH gradient can develop only if the membrane is also permeable to a major anion (e.g., Cl�) or if the H�^ ions are exchanged for another cation (e.g., K�). In either case, proton movement does not lead to a voltage gradient across the membrane because there is al- ways an equal concentration of positive and negative ions on each side of the membrane. This is the situation in the chloroplast thylakoid membrane during photosynthesis, as we discuss later. Compared with chloroplasts, then, a greater portion of the pmf in mitochondria is due to the membrane electric potential, and the actual pH gradient is smaller. Since a difference of one pH unit represents a tenfold dif- ference in H�^ concentration, a pH gradient of one unit across a membrane is equivalent to an electric potential of 59 mV at 20 �C according to the Nernst equation (Chap- ter 7). Thus we can define the proton-motive force, pmf, as
where R is the gas constant of 1.987 cal/(degree mol), T is the temperature (in degrees Kelvin), F is the Faraday constant [23,062 cal/(V mol)], and is the transmembrane electric potential; and pmf are measured in millivolts. Measure- ments on respiring mitochondria have shown that the electric potential across the inner membrane is �160 mV (nega- tive inside matrix) and that �pH is ≈1.0 (equivalent to ≈60 mV). Thus the total pmf is �220 mV, with the trans- membrane electric potential responsible for about 73 percent. Because mitochondria are much too small to be impaled with electrodes, the electric potential and pH gradient across the inner mitochondrial membrane cannot be deter-
pmf ° � a
RT
F
� ¢pHb ° � 59 ¢pH
8.2 •^ Electron Transport and Generation of the Proton-Motive Force 317
mined by direct measurement. However, researchers can measure the inside pH by trapping fluorescent pH-sensitive dyes inside vesicles formed from the inner mitochondrial membrane. They also can determine the electric potential by adding radioactive 42 K�^ ions and a trace amount of valino- mycin to a suspension of respiring mitochondria. Although the inner membrane is normally impermeable to K�, vali- nomycin is an ionophore , a small lipid-soluble molecule that selectively binds a specific ion (in this case, K�^ ) in its hy- drophilic interior and carries it across otherwise imperme- able membranes. In the presence of valinomycin, 42 K� equilibrates across the inner membrane of isolated mito- chondria in accordance with the electric potential; the more negative the matrix side of the membrane, the more 42 K� will accumulate in the matrix. Addition of small amounts of valinomycin and radioac- tive K�^ has little effect on oxidative phosphorylation by a suspension of respiring mitochondria. At equilibrium, the measured concentration of radioactive K�^ ions in the matrix, [Kin], is about 500 times greater than that in the surrounding medium, [K (^) out ]. Substitution of this value into the Nernst equation shows that the electric potential E (in mV) across the inner membrane in respiring mitochondria is �160 mV, with the inside negative:
Electron Transport in Mitochondria Is Coupled to Proton Translocation The coupling between electron transport from NADH (or FADH 2 ) to O 2 and proton transport across the inner mito- chondrial membrane, which generates the proton-motive force, also can be demonstrated experimentally with isolated mitochondria (Figure 8-14). As soon as O 2 is added to a sus- pension of mitochondria, the medium outside the mitochon- dria becomes acidic. During electron transport from NADH to O 2 , protons translocate from the matrix to the intermem- brane space; since the outer membrane is freely permeable to protons, the pH of the outside medium is lowered briefly. The measured change in pH indicates that about 10 protons are transported out of the matrix for every electron pair transferred from NADH to O 2. When this experiment is repeated with succinate rather than NADH as the reduced substrate, the medium outside the mitochondria again becomes acidic, but less so. Recall that oxidation of succinate to fumarate in the citric acid cycle generates FADH 2 (see Figure 8-9). Because electrons in FADH 2 have less potential energy (43. kcal/mol) than electrons in NADH (52.6 kcal/mole), FADH 2 transfers electrons to the respiratory chain at a later point than NADH does. As a result, electron transport from FADH 2 (or succinate) results in translocation of fewer pro- tons from the matrix, and thus a smaller change in pH (see Figure 8-13).
E � 59 log
3 Kin 4 3 Kout 4
� 59 log 500 � 160 mV
In the respiratory chain, electrons move through the cy- tochromes in the following order: b, c 1 , c, a, and a 3 (see Figure 8-13). The various cytochromes have slightly different heme groups and axial ligands, which generate different environments for the Fe ion. Therefore, each cy- tochrome has a different reduction potential, or tendency to accept an electron—an important property dictating the unidirectional electron flow along the chain. Because the heme ring in cytochromes consists of alternating double- and single-bonded atoms, a large number of resonance forms exist, and the extra electron is delocalized to the heme carbon and nitrogen atoms as well as to the Fe ion. All the cytochromes, except cytochrome c , are components of multiprotein complexes in the inner mitochondrial membrane. Although cytochrome c comprises a heme- protein complex, it moves freely by diffusion in the inter- membrane space. Iron-sulfur clusters are nonheme, iron-containing pros- thetic groups consisting of Fe atoms bonded both to inor- ganic S atoms and to S atoms on cysteine residues on a protein (Figure 8-15b). Some Fe atoms in the cluster bear a �2 charge; others have a �3 charge. However, the net charge of each Fe atom is actually between �2 and �3 be- cause electrons in the outermost orbits are dispersed among the Fe atoms and move rapidly from one atom to another. Iron-sulfur clusters accept and release electrons one at a time; the additional electron is also dispersed over all the Fe atoms in the cluster. Coenzyme Q (CoQ), also called ubiquinone, is the only electron carrier in the respiratory chain that is not a protein- bound prosthetic group. It is a carrier of hydrogen atoms, that is, protons plus electrons. The oxidized quinone form of CoQ can accept a single electron to form a semiquinone, a charged free radical denoted by CoQ� ·. Addition of a second electron
8.2 •^ Electron Transport and Generation of the Proton-Motive Force 319
and two protons to CoQ� · forms dihydroubiquinone (CoQH 2 ), the fully reduced form (Figure 8-16). Both CoQ and CoQH 2 are soluble in phospholipids and diffuse freely in the inner mitochondrial membrane.
H 2 C
H 3 C CH
CH C C C C^ C C
C
N
N
C
C C C
C C
C
C C
C C
C N
C
CH 3
N
CH 2
CH 3
Fe^2 +
CH 2 H 2 C −O 2 C CH 2 H 2 C CO 2 −
H 3 C
(a)
Fe Fe
S
Cys-S S
Cys-S S-Cys
S-Cys
Protein
(b)
▲ FIGURE 8-15 Heme and iron-sulfur prosthetic groups in the respiratory (electron-transport) chain. (a) Heme portion of cytochromes b L and b H , which are components of the CoQH 2 – cytochrome c reductase complex. The same porphyrin ring (yellow) is present in all hemes. The chemical substituents attached to the porphyrin ring differ in the other cytochromes in
the respiratory chain. All hemes accept and release one electron at a time. (b) Dimeric iron-sulfur cluster (2Fe-2S). Each Fe atom is bonded to four S atoms: two are inorganic sulfur and two are in cysteine side chains of the associated protein. (Note that only the two inorganic S atoms are counted in the chemical formula.) All Fe-S clusters accept and release one electron at a time.
H 3 CO CH 3 CH^3 H 3 CO (CH 2 CH C CH 2 ) 10 H Ubiquinone (CoQ) (oxidized form)
O
O
Semiquinone (CoQ � ) (free radical)
Dihydroquinone (CoQH 2 ) (fully reduced form)
e�
H 3 CO CH 3 CH^3 H 3 CO (CH 2 CH C CH 2 ) 10 H
O�
O 2H�^ � e�
H 3 CO CH 3 CH^3 H 3 CO (CH 2 CH C CH 2 ) 10 H
OH
OH
▲ FIGURE 8-16 Oxidized and reduced forms of coenzyme Q (CoQ), which carries two protons and two electrons. Because of its long hydrocarbon “tail” of isoprene units, CoQ is soluble in the hydrophobic core of phospholipid bilayers and is very mobile. Reduction of CoQ to the fully reduced form, QH 2 , occurs in two steps with a half-reduced free-radical intermediate, called semiquinone.
As shown in Figure 8-17, CoQ accepts electrons released from the NADH-CoQ reductase complex (I) and the succinate-CoQ reductase complex (II) and donates them to the CoQH 2 –cytochrome c reductase complex (III). Impor- tantly, reduction and oxidation of CoQ are coupled to pump- ing of protons. Whenever CoQ accepts electrons, it does so at a binding site on the cytosolic (matrix) face of the protein complex, always picking up protons from the medium fac- ing the cytosolic face. Whenever CoQH 2 releases its electrons, it does so at a binding site on the exoplasmic face of the pro- tein complex, releasing protons into the exoplasmic medium (intermembrane space). Thus transport of each pair of elec- trons by CoQ is obligatorily coupled to movement of two protons from the cytosolic to the exoplasmic medium.
NADH-CoQ Reductase (Complex I) Electrons are carried
from NADH to CoQ by the NADH-CoQ reductase com- plex. NAD�^ is exclusively a two-electron carrier: it accepts or releases a pair of electrons at a time. In the NADH-CoQ reductase complex, electrons first flow from NADH to FMN (flavin mononucleotide), a cofactor related to FAD, then to an iron-sulfur cluster, and finally to CoQ (see Figure 8-17). FMN, like FAD, can accept two electrons, but does so one electron at a time. The overall reaction catalyzed by this complex is
NADH � CoQ � 2 H�^ NAD�^ � H�^ � CoQH 2 (Reduced) (Oxidized) (Oxidized) (Reduced)
Each transported electron undergoes a drop in potential of ≈360 mV, equivalent to a � G �� of �16.6 kcal/mol for the
320 CHAPTER 8 •^ Cellular Energetics
two electrons transported (see Figure 8-13). Much of this re- leased energy is used to transport four protons across the inner membrane per molecule of NADH oxidized by the NADH-CoQ reductase complex.
Succinate-CoQ Reductase (Complex II) Succinate dehydro-
genase, the enzyme that oxidizes a molecule of succinate to fumarate in the citric acid cycle, is an integral component of the succinate-CoQ reductase complex. The two electrons re- leased in conversion of succinate to fumarate are transferred first to FAD, then to an iron-sulfur cluster, and finally to CoQ (see Figure 8-17). The overall reaction catalyzed by this complex is
Succinate � CoQ fumarate � CoQH (^2) (Reduced) (Oxidized) (Oxidized) (Reduced) Although the � G �� for this reaction is negative, the released energy is insufficient for proton pumping. Thus no protons are translocated across the membrane by the succinate-CoQ reductase complex, and no proton-motive force is generated in this part of the respiratory chain.
CoQH 2 –Cytochrome c Reductase (Complex III) A CoQH 2
generated either by complex I or complex II donates two electrons to the CoQH 2 –cytochrome c reductase complex, regenerating oxidized CoQ. Concomitantly it releases two protons picked up on the cytosolic face into the intermem- brane space, generating part of the proton-motive force. Within complex III, the released electrons first are trans- ferred to an iron-sulfur cluster within complex III and then to
Fe-S
2 e−
FMN
NADH NAD+^ + H+
4 H+
4 H+
2 H+
Fe-S
(^1) / 2 O 2 + 2 H+
NADH-CoQ reductase (complex I)
CoQH 2 –cytochromec reductase (complex III)
Cytochromec oxidase (complex IV)
2 H+
2 H+
Cytc (^1)
H 2 O
+++
- – –
Intermembrane space
Matrix 2 H+
Cytc Cytc
Succinate-CoQ reductase (complex II)
Fe-S
FAD
2 H+ e−
Cu (^) b•Cyta (^3)
Cyta
Fe-S Cu^ a
CoQH (^2)
CoQH (^2)
CoQ Cytb (^) H
Cytb (^) L
e−
Cytc
e−
Succinate Fumarate + 2 H+
Fe-S
Fe-S
CoQH (^2)
CoQH (^2)
CoQ
2 H+^ 2 H+
2 H+
▲ FIGURE 8-17 Overview of multiprotein complexes, bound prosthetic groups, and associated mobile carriers in the respiratory chain. Blue arrows indicate electron flow; red arrows indicate proton translocation. ( Left ) Pathway from NADH. A total of 10 protons are translocated per pair of electrons that flow from NADH to O 2. The protons released into the matrix space during oxidation of NADH by NADH-CoQ reductase are consumed in the formation of water from O 2 by cytochrome c oxidase, resulting in no net proton translocation from these reactions. ( Right ) Pathway
from succinate. During oxidation of succinate to fumarate and reduction of CoQ by the succinate-CoQ reductase complex, no protons are translocated across the membrane. The remainder of electron transport from CoQH 2 proceeds by the same pathway as in the left diagram. Thus for every pair of electrons transported from succinate to O 2 , six protons are translocated from the matrix into the intermembrane space. Coenzyme Q and cytochrome c function as mobile carriers in electron transport from both NADH and succinate. See the text for details.
M E D I A
C O N N E C T I O N S
Focus Animation: Electron Transport