Download OXIDATIVE PHOSPHORYLATION AND ... and more Study notes Medicine in PDF only on Docsity!
chapter
O
xidative phosphorylation is the culmination of energy- yielding metabolism in aerobic organisms. All oxi- dative steps in the degradation of carbohydrates, fats, and amino acids converge at this final stage of cellular respiration, in which the energy of oxidation drives the synthesis of ATP. Photophosphorylation is the means by which photosynthetic organisms capture the energy of sunlight—the ultimate source of energy in the bio- sphere—and harness it to make ATP. Together, oxida- tive phosphorylation and photophosphorylation account for most of the ATP synthesized by most organisms most of the time. In eukaryotes, oxidative phosphorylation occurs in mitochondria, photophosphorylation in chloroplasts. Oxidative phosphorylation involves the reduction of O to H 2 O with electrons donated by NADH and FADH 2 ; it occurs equally well in light or darkness. Photophosphor- ylation involves the oxidation of H 2 O to O 2 , with NADP�^ as ultimate electron acceptor; it is absolutely dependent on the energy of light. Despite their differ- ences, these two highly efficient energy-converting processes have fundamentally similar mechanisms. Our current understanding of ATP synthesis in mi- tochondria and chloroplasts is based on the hypothesis, introduced by Peter Mitchell in 1961, that transmem- brane differences in proton concentration are the reser- voir for the energy extracted from biological oxidation reactions. This chemiosmotic theory has been ac- cepted as one of the great unifying principles of twen- tieth century biology. It provides insight into the processes of oxidative phosphorylation and photophos- phorylation, and into such apparently disparate energy transductions as active transport across membranes and the motion of bacterial flagella. Oxidative phosphorylation and photophosphoryla- tion are mechanistically similar in three respects. (1) Both
OXIDATIVE PHOSPHORYLATION
AND PHOTOPHOSPHORYLATION
If an idea presents itself to us, we must not reject it
simply because it does not agree with the logical
deductions of a reigning theory.
—Claude Bernard, An Introduction to the Study of Experimental Medicine, 1813
The aspect of the present position of consensus that I
find most remarkable and admirable, is the altruism and
generosity with which former opponents of the
chemiosmotic hypothesis have not only come to accept it,
but have actively promoted it to the status of a theory.
—Peter Mitchell, Nobel Address, 1978
OXIDATIVE PHOSPHORYLATION
19.1 Electron-Transfer Reactions in Mitochondria 691
19.2 ATP Synthesis 704
19.3 Regulation of Oxidative Phosphorylation 716
19.4 Mitochondrial Genes: Their Origin and the Effects
of Mutations 719
19.5 The Role of Mitochondria in Apoptosis and
Oxidative Stress 721
PHOTOSYNTHESIS: HARVESTING LIGHT ENERGY
19.6 General Features of Photophosphorylation 723
19.7 Light Absorption 725
19.8 The Central Photochemical Event: Light-Driven
Electron Flow 730
19.9 ATP Synthesis by Photophosphorylation 740
processes involve the flow of electrons through a chain of membrane-bound carriers. (2) The free energy made available by this “downhill” (exergonic) electron flow is coupled to the “uphill” transport of protons across a proton-impermeable membrane, conserving the free energy of fuel oxidation as a transmembrane electro- chemical potential (p. 391). (3) The transmembrane flow of protons down their concentration gradient through specific protein channels provides the free energy for synthesis of ATP, catalyzed by a membrane protein complex (ATP synthase) that couples proton flow to phosphorylation of ADP. We begin this chapter with oxidative phosphoryla- tion. We first describe the components of the electron- transfer chain, their organization into large functional complexes in the inner mitochondrial membrane, the path of electron flow through them, and the proton movements that accompany this flow. We then consider the remarkable enzyme complex that, by “rotational catalysis,” captures the energy of proton flow in ATP, and the regulatory mechanisms that coordinate oxida- tive phosphorylation with the many catabolic pathways by which fuels are oxidized. With this understanding of mitochondrial oxidative phosphorylation, we turn to photophosphorylation, looking first at the absorption of light by photosynthetic pigments, then at the light- driven flow of electrons from H 2 O to NADP�^ and the molecular basis for coupling electron and proton flow. We also consider the similarities of structure and mech- anism between the ATP synthases of chloroplasts and mitochondria, and the evolutionary basis for this con- servation of mechanism.
OXIDATIVE PHOSPHORYLATION
19.1 Electron-Transfer Reactions
in Mitochondria
The discovery in 1948 by Eugene Kennedy and Albert Lehninger that mitochondria are the site of oxidative phosphorylation in eukaryotes marked the beginning of the modern phase of studies in biological energy transduc- tions. Mitochondria, like gram- negative bacteria, have two membranes (Fig. 19–1). The outer mitochondrial membrane is readily permeable to small molecules ( M r �5,000) and ions, which move freely through transmembrane chan- nels formed by a family of inte- gral membrane proteins called porins. The inner membrane is impermeable to most small
molecules and ions, including protons (H�); the only species that cross this membrane do so through specific transporters. The inner membrane bears the compo- nents of the respiratory chain and the ATP synthase. The mitochondrial matrix, enclosed by the inner membrane, contains the pyruvate dehydrogenase com- plex and the enzymes of the citric acid cycle, the fatty
19.1 Electron-Transfer Reactions in Mitochondria 691
Outer membrane Freely permeable to small molecules and ions
ATP synthase (FoF1) Cristae
Impermeable to most small molecules and ions, including H� Contains:
Contains:
Ribosomes
Porin channels
- Respiratory electron carriers (Complexes I–IV)
- ADP-ATP translocase
- ATP synthase (FoF1)
- Other membrane transporters - Pyruvate dehydrogenase complex - Citric acid cycle enzymes - Amino acid oxidation enzymes
- DNA, ribosomes
- Many other enzymes
- ATP, ADP, Pi, Mg^2 �, Ca^2 �, K�
- Many soluble metabolic intermediates
Inner membrane
Matrix
- Fatty acid -oxidation enzymes
�
Albert L. Lehninger, 1917–
FIGURE 19–1 Biochemical anatomy of a mitochondrion. The convo- lutions (cristae) of the inner membrane provide a very large surface area. The inner membrane of a single liver mitochondrion may have more than 10,000 sets of electron-transfer systems (respiratory chains) and ATP synthase molecules, distributed over the membrane surface. Heart mitochondria, which have more profuse cristae and thus a much larger area of inner membrane, contain more than three times as many sets of electron-transfer systems as liver mitochondria. The mitochon- drial pool of coenzymes and intermediates is functionally separate from the cytosolic pool. The mitochondria of invertebrates, plants, and mi- crobial eukaryotes are similar to those shown here, but with much vari- ation in size, shape, and degree of convolution of the inner membrane.
potential is therefore that of the particular flavoprotein, not that of isolated FAD or FMN. The flavin nucleotide should be considered part of the flavoprotein’s active site rather than a reactant or product in the electron- transfer reaction. Because flavoproteins can participate in either one- or two-electron transfers, they can serve as intermediates between reactions in which two elec- trons are donated (as in dehydrogenations) and those in which only one electron is accepted (as in the reduction of a quinone to a hydroquinone, described below).
Electrons Pass through a Series
of Membrane-Bound Carriers
The mitochondrial respiratory chain consists of a series of sequentially acting electron carriers, most of which are integral proteins with prosthetic groups capable of accepting and donating either one or two electrons. Three types of electron transfers occur in oxidative phosphorylation: (1) direct transfer of electrons, as in the reduction of Fe^3 �^ to Fe^2 �; (2) transfer as a hydro- gen atom (H�^ � e �); and (3) transfer as a hydride ion ( : H�), which bears two electrons. The term reducing equivalent is used to designate a single electron equiv- alent transferred in an oxidation-reduction reaction. In addition to NAD and flavoproteins, three other types of electron-carrying molecules function in the res- piratory chain: a hydrophobic quinone (ubiquinone) and two different types of iron-containing proteins (cyto- chromes and iron-sulfur proteins). Ubiquinone (also called coenzyme Q, or simply Q ) is a lipid-soluble ben- zoquinone with a long isoprenoid side chain (Fig. 19–2). The closely related compounds plastoquinone (of plant chloroplasts) and menaquinone (of bacteria) play roles analogous to that of ubiquinone, carrying electrons in membrane-associated electron-transfer chains. Ubiqui- none can accept one electron to become the semi- quinone radical (�QH) or two electrons to form ubiquinol (QH 2 ) (Fig. 19–2) and, like flavoprotein carriers, it can act at the junction between a two-electron donor and a one-electron acceptor. Because ubiquinone is both small and hydrophobic, it is freely diffusible within the lipid bilayer of the inner mitochondrial membrane and can shuttle reducing equivalents between other, less mobile electron carriers in the membrane. And because it car- ries both electrons and protons, it plays a central role in coupling electron flow to proton movement. The cytochromes are proteins with characteristic strong absorption of visible light, due to their iron- containing heme prosthetic groups (Fig. 19–3). Mito- chondria contain three classes of cytochromes, desig- nated a, b, and c, which are distinguished by differences in their light-absorption spectra. Each type of cyto- chrome in its reduced (Fe^2 �) state has three absorp- tion bands in the visible range (Fig. 19–4). The longest- wavelength band is near 600 nm in type a cytochromes,
near 560 nm in type b, and near 550 nm in type c. To distinguish among closely related cytochromes of one type, the exact absorption maximum is sometimes used in the names, as in cytochrome b 562. The heme cofactors of a and b cytochromes are tightly, but not covalently, bound to their associated pro- teins; the hemes of c -type cytochromes are covalently attached through Cys residues (Fig. 19–3). As with the flavoproteins, the standard reduction potential of the heme iron atom of a cytochrome depends on its inter- action with protein side chains and is therefore differ- ent for each cytochrome. The cytochromes of type a and b and some of type c are integral proteins of the inner mitochondrial membrane. One striking exception is the cytochrome c of mitochondria, a soluble protein that associates through electrostatic interactions with the outer surface of the inner membrane. We encountered cytochrome c in earlier dis- cussions of protein structure (see Fig. 4–18). In iron-sulfur proteins, first discovered by Helmut Beinert, the iron is present not in heme but in association with inorganic sulfur atoms or with the sulfur atoms of Cys residues in the protein, or both. These iron-sulfur (Fe-S)
19.1 Electron-Transfer Reactions in Mitochondria 693
O^ •
CH C H
R
OH
CH 3
CH 3 O
(CH 2
O
CH 3 O
CH 3
CH 2 ) (^10) Ubiquinone (Q) (fully oxidized)
Semiquinone radical (•QH)
Ubiquinol (QH 2 ) (fully reduced)
H�^ � e �
O
CH 3 O CH 3
CH 3 O
H�^ � e �
OH
OH
R
CH 3 O CH 3
CH 3 O
FIGURE 19–2 Ubiquinone (Q, or coenzyme Q). Complete reduction of ubiquinone requires two electrons and two protons, and occurs in two steps through the semiquinone radical intermediate.
Helmut Beinert
centers range from simple structures with a single Fe atom coordinated to four Cys OSH groups to more com- plex Fe-S centers with two or four Fe atoms (Fig. 19–5). Rieske iron-sulfur proteins (named after their dis- coverer, John S. Rieske) are a variation on this theme, in which one Fe atom is coordinated to two His residues rather than two Cys residues. All iron-sulfur proteins participate in one-electron transfers in which one iron atom of the iron-sulfur cluster is oxidized or reduced. At least eight Fe-S proteins function in mitochondrial electron transfer. The reduction potential of Fe-S pro- teins varies from �0.65 V to �0.45 V, depending on the microenvironment of the iron within the protein. In the overall reaction catalyzed by the mitochon- drial respiratory chain, electrons move from NADH, suc- cinate, or some other primary electron donor through flavoproteins, ubiquinone, iron-sulfur proteins, and cy- tochromes, and finally to O 2. A look at the methods used to determine the sequence in which the carriers act is instructive, as the same general approaches have been used to study other electron-transfer chains, such as those of chloroplasts.
694 Chapter 19 Oxidative Phosphorylation and Photophosphorylation
Fe N
N N N CH 3
CH 3
CH 2 CH 2 COO�
CH 2
CH
CH 2
CH
CH (^3)
Heme A (in a -type cytochromes)
Fe N
N N N CH 3
CH 3
CH 3
CH 3
CH 2 CH 2 COO�
CH 2
CH 2
CHO
CH 2
CH 2 CH
Iron protoporphyrin IX (in b -type cytochromes)
COO�
CH 3
OH
Cys
S Cys
Fe N
N N N CH 3
CH 3
CH 3
CH 3
CH 2 CH 2 COO�
CH 2 CH (^2)
Heme C (in c -type cytochromes)
COO�
CH 3
CH CH (^2) S CH
CH CH (^2)
CH (^3) CH 3
CH 3 CH 3
COO�
FIGURE 19–3 Prosthetic groups of cytochromes. Each group consists of four five-membered, nitrogen-containing rings in a cyclic structure called a porphyrin. The four nitrogen atoms are coordinated with a central Fe ion, either Fe 2 �^ or Fe^3 �. Iron protoporphyrin IX is found in b -type cytochromes and in hemoglobin and myoglobin (see Fig. 4–17). Heme c is covalently bound to the protein of cytochrome c through thioether bonds to two Cys residues. Heme a, found in the a -type cytochromes, has a long isoprenoid tail attached to one of the five-membered rings. The conjugated double-bond system (shaded pink) of the porphyrin ring accounts for the absorption of visible light by these hemes.
100
Relative light absorption (%)
50
300 400 500 600 Wavelength (nm)
Oxidized cyt c
Reduced cyt c
�
�
�
0
FIGURE 19–4 Absorption spectra of cytochrome c (cyt c ) in its oxi- dized (red) and reduced (blue) forms. Also labeled are the character- istic � , � , and � bands of the reduced form.
of electron carriers involves reducing the entire chain of carriers experimentally by providing an electron source but no electron acceptor (no O 2 ). When O 2 is suddenly introduced into the system, the rate at which each electron carrier becomes oxidized (measured spectroscopically) reveals the order in which the car- riers function. The carrier nearest O 2 (at the end of the chain) gives up its electrons first, the second carrier from the end is oxidized next, and so on. Such exper- iments have confirmed the sequence deduced from standard reduction potentials. In a final confirmation, agents that inhibit the flow of electrons through the chain have been used in com- bination with measurements of the degree of oxidation of each carrier. In the presence of O 2 and an electron donor, carriers that function before the inhibited step become fully reduced, and those that function after this step are completely oxidized (Fig. 19–6). By using sev- eral inhibitors that block different steps in the chain, in- vestigators have determined the entire sequence; it is the same as deduced in the first two approaches.
Electron Carriers Function in Multienzyme Complexes
The electron carriers of the respiratory chain are or- ganized into membrane-embedded supramolecular
complexes that can be physically separated. Gentle treatment of the inner mitochondrial membrane with detergents allows the resolution of four unique electron- carrier complexes, each capable of catalyzing electron transfer through a portion of the chain (Table 19–3; Fig. 19–7). Complexes I and II catalyze electron transfer to ubiquinone from two different electron donors: NADH (Complex I) and succinate (Complex II). Complex III carries electrons from reduced ubiquinone to cyto- chrome c, and Complex IV completes the sequence by transferring electrons from cytochrome c to O 2. We now look in more detail at the structure and function of each complex of the mitochondrial respira- tory chain.
Complex I: NADH to Ubiquinone Figure 19–8 illustrates the relationship between Complexes I and II and ubiquinone. Complex I, also called NADH:ubiquinone oxidore- ductase or NADH dehydrogenase, is a large enzyme composed of 42 different polypeptide chains, including an FMN-containing flavoprotein and at least six iron- sulfur centers. High-resolution electron microscopy shows Complex I to be L-shaped, with one arm of the L in the membrane and the other extending into the ma- trix. As shown in Figure 19–9, Complex I catalyzes two simultaneous and obligately coupled processes: (1) the
696 Chapter 19 Oxidative Phosphorylation and Photophosphorylation
NADH Q
Cyt c 1
Cyt ( a � a 3 ) O (^2)
rotenone
antimycin A
CN�^ or CO
NADH Q Cyt b Cyt c O (^2)
NADH Q O (^2)
Cyt b Cyt c 1 Cyt c
Cyt b Cyt c 1 Cyt c Cyt
Cyt ( a � a 3 )
( a � a 3 )
FIGURE 19–6 Method for determining the sequence of electron carriers. This method measures the effects of inhibitors of electron transfer on the oxidation state of each carrier. In the presence of an electron donor and O 2 , each inhibitor causes a characteristic pattern of oxidized/reduced carriers: those before the block become reduced (blue), and those after the block become oxidized (pink).
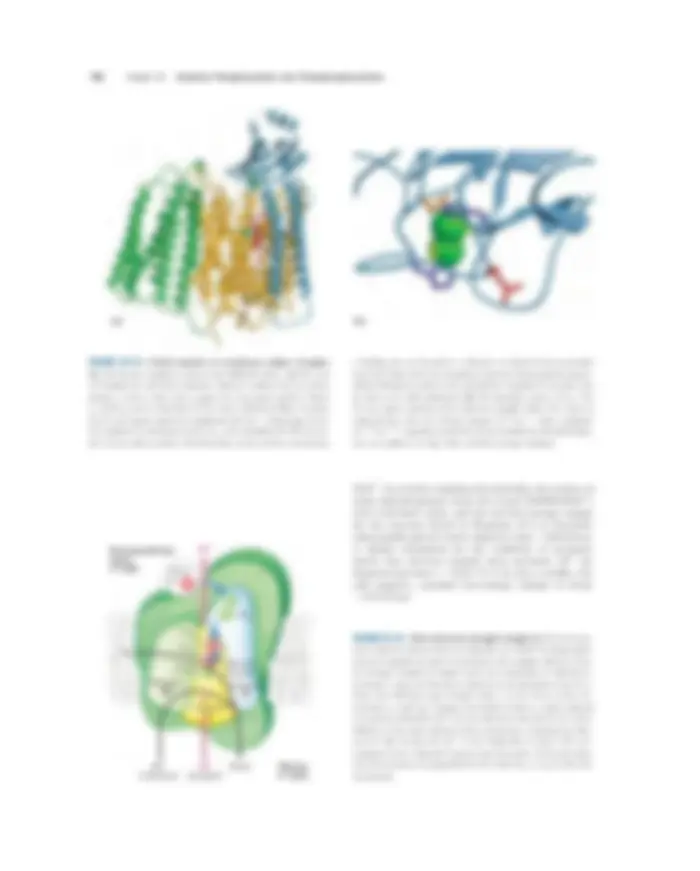
TABLE 19–3 The Protein Components of the Mitochondrial Electron-Transfer Chain
Enzyme complex/protein Mass (kDa) Number of subunits *^ Prosthetic group(s)
I NADH dehydrogenase 850 43 (14) FMN, Fe-S II Succinate dehydrogenase 140 4 FAD, Fe-S III Ubiquinone cytochrome c oxidoreductase 250 11 Hemes, Fe-S Cytochrome c†^13 1 Heme IV Cytochrome oxidase 160 13 (3–4) Hemes; Cu (^) A, CuB
*Numbers of subunits in the bacterial equivalents in parentheses. †Cytochrome c is not part of an enzyme complex; it moves between Complexes III and IV as a freely soluble protein.
exergonic transfer to ubiquinone of a hydride ion from NADH and a proton from the matrix, expressed by NADH � H�^ � Q (^) On NAD�^ � QH 2 (19–1) and (2) the endergonic transfer of four protons from the matrix to the intermembrane space. Complex I is there- fore a proton pump driven by the energy of electron transfer, and the reaction it catalyzes is vectorial: it moves protons in a specific direction from one location (the matrix, which becomes negatively charged with the departure of protons) to another (the intermembrane space, which becomes positively charged). To empha- size the vectorial nature of the process, the overall re- action is often written with subscripts that indicate the location of the protons: P for the positive side of the in- ner membrane (the intermembrane space), N for the negative side (the matrix): NADH � 5H� N � Q (^) On NAD�^ � QH 2 � 4H� P (19–2)
19.1 Electron-Transfer Reactions in Mitochondria 697
Osmotic rupture
Inner membrane fragments Outer membrane fragments discarded ATP synthase
IV III
II
I
I II III IV ATP synthase
NADH Q Suc- cinate
Q Q Cyt c Cyt c O 2 ATP ADP � Pi Reactions catalyzed by isolated fractions in vitro
Solubilization with detergent followed by ion-exchange chromatography
Treatment with digitonin
FIGURE 19–7 Separation of functional complexes of the respiratory chain. The outer mitochondrial membrane is first removed by treat- ment with the detergent digitonin. Fragments of inner membrane are then obtained by osmotic rupture of the mitochondria, and the frag- ments are gently dissolved in a second detergent. The resulting mix- ture of inner membrane proteins is resolved by ion-exchange chro- matography into different complexes (I through IV) of the respiratory chain, each with its unique protein composition (see Table 19–3), and the enzyme ATP synthase (sometimes called Complex V). The isolated Complexes I through IV catalyze transfers between donors (NADH and succinate), intermediate carriers (Q and cytochrome c ), and O 2 , as shown. In vitro, isolated ATP synthase has only ATP-hydrolyzing (ATPase), not ATP-synthesizing, activity.
I II
Intermembrane space
Matrix
Fe-S
Fe-S
FAD
Glycerol 3-phosphate (cytosolic)
glycerol 3-phosphate dehydrogenase
FAD
FMN
NADH NAD+^ Succinate ETF:Qoxidoreductase
acyl-CoA dehydrogenase
ETF (FAD)
Fe-S (FAD)
Fatty acyl–CoA
FAD
Q
FIGURE 19–8 Path of electrons from NADH, succinate, fatty acyl–CoA, and glycerol 3-phosphate to ubiquinone. Electrons from NADH pass through a flavoprotein to a series of iron-sulfur proteins (in Complex I) and then to Q. Electrons from succinate pass through a flavoprotein and several Fe-S centers (in Complex II) on the way to Q. Glycerol 3-phosphate donates electrons to a flavoprotein (glycerol 3-phosphate dehydrogenase) on the outer face of the inner mito- chondrial membrane, from which they pass to Q. Acyl-CoA dehydro- genase (the first enzyme of � oxidation) transfers electrons to electron- transferring flavoprotein (ETF), from which they pass to Q via ETF:ubiquinone oxidoreductase.
flavoprotein acyl-CoA dehydrogenase (see Fig. 17–8), involves transfer of electrons from the substrate to the FAD of the dehydrogenase, then to electron-transferring flavoprotein (ETF), which in turn passes its electrons to ETF : ubiquinone oxidoreductase (Fig. 19–8). This enzyme transfers electrons into the respiratory chain by reducing ubiquinone. Glycerol 3-phosphate, formed ei- ther from glycerol released by triacylglycerol breakdown or by the reduction of dihydroxyacetone phosphate from glycolysis, is oxidized by glycerol 3-phosphate dehydrogenase (see Fig. 17–4). This enzyme is a flavo- protein located on the outer face of the inner mito- chondrial membrane, and like succinate dehydrogenase and acyl-CoA dehydrogenase it channels electrons into the respiratory chain by reducing ubiquinone (Fig. 19–8). The important role of glycerol 3-phosphate de- hydrogenase in shuttling reducing equivalents from cytosolic NADH into the mitochondrial matrix is de- scribed in Section 19.2 (see Fig. 19–28). The effect of each of these electron-transferring enzymes is to con- tribute to the pool of reduced ubiquinone. QH 2 from all these reactions is reoxidized by Complex III.
Complex III: Ubiquinone to Cytochrome c The next respi- ratory complex, Complex III, also called cytochrome bc 1 complex or ubiquinone:cytochrome c oxidore- ductase, couples the transfer of electrons from ubiquinol (QH 2 ) to cytochrome c with the vectorial transport of protons from the matrix to the intermem- brane space. The determination of the complete struc- ture of this huge complex (Fig. 19–11) and of Complex IV (below) by x-ray crystallography, achieved between 1995 and 1998, were landmarks in the study of mito- chondrial electron transfer, providing the structural framework to integrate the many biochemical observa- tions on the functions of the respiratory complexes. Based on the structure of Complex III and detailed biochemical studies of the redox reactions, a reasonable model has been proposed for the passage of electrons
19.1 Electron-Transfer Reactions in Mitochondria 699
The heme b of Complex II is apparently not in the direct path of electron transfer; it may serve instead to reduce the frequency with which electrons “leak” out of the system, moving from succinate to mo- lecular oxygen to produce the reactive oxygen species (ROS) hydrogen peroxide (H 2 O 2 ) and the superoxide radical (�O 2 �) described in Section 19.5. Humans with point mutations in Complex II subunits near heme b or the quinone-binding site suffer from hereditary para- ganglioma. This inherited condition is characterized by benign tumors of the head and neck, commonly in the carotid body, an organ that senses O 2 levels in the blood. These mutations result in greater production of ROS and perhaps greater tissue damage during succinate oxidation. ■ Other substrates for mitochondrial dehydrogenases pass electrons into the respiratory chain at the level of ubiquinone, but not through Complex II. The first step in the � oxidation of fatty acyl–CoA, catalyzed by the
FIGURE 19–10 Structure of Complex II (succinate dehydrogenase) of E. coli (PDB ID 1NEK). The enzyme has two transmembrane sub- units, C (green) and D (blue); the cytoplasmic extensions contain sub- units B (orange) and A (purple). Just behind the FAD in subunit A (gold) is the binding site for succinate (occupied in this crystal structure by the inhibitor oxaloacetate, green). Subunit B has three sets of Fe-S cen- ters (yellow and red); ubiquinone (yellow) is bound to subunit C; and heme b (purple) is sandwiched between subunits C and D. A cardi- olipin molecule is so tightly bound to subunit C that it shows up in the crystal structure (gray spacefilling). Electrons move (blue arrows) from succinate to FAD, then through the three Fe-S centers to ubiquinone. The heme b is not on the main path of electron transfer but protects against the formation of reactive oxygen species (ROS) by electrons that go astray.
Substrate binding site
Cytoplasm ( N side)
C
B
D
A
FAD
Fe-S centers
Periplasm ( P side)
Cardiolipin
Ubiquinone
QH 2 Heme b
and protons through the complex. The net equation for the redox reactions of this Q cycle (Fig. 19–12) is
QH 2 � 2 cyt c 1 (oxidized) � 2H� N On Q � 2 cyt c 1 (reduced) � 4H� P (19–3)
The Q cycle accommodates the switch between the two- electron carrier ubiquinone and the one-electron carri- ers—cytochromes b 562 , b 566 , c 1 , and c —and explains the measured stoichiometry of four protons translocated per pair of electrons passing through the Complex III to cytochrome c. Although the path of electrons through this segment of the respiratory chain is complicated, the net effect of the transfer is simple: QH 2 is oxidized to Q and two molecules of cytochrome c are reduced. Cytochrome c (see Fig. 4–18) is a soluble protein of the intermembrane space. After its single heme accepts an electron from Complex III, cytochrome c moves to Complex IV to donate the electron to a binuclear cop- per center.
Complex IV: Cytochrome c to O 2 In the final step of the respiratory chain, Complex IV, also called cytochrome oxidase, carries electrons from cytochrome c to mo- lecular oxygen, reducing it to H 2 O. Complex IV is a large enzyme (13 subunits; M r 204,000) of the inner mito- chondrial membrane. Bacteria contain a form that is much simpler, with only three or four subunits, but still capable of catalyzing both electron transfer and proton pumping. Comparison of the mitochondrial and bacter- ial complexes suggests that three subunits are critical to the function (Fig. 19–13). Mitochondrial subunit II contains two Cu ions com- plexed with the OSH groups of two Cys residues in a binuclear center (Cu (^) A; Fig. 19–13b) that resembles the 2Fe-2S centers of iron-sulfur proteins. Subunit I con- tains two heme groups, designated a and a 3 , and an- other copper ion (CuB). Heme a 3 and CuB form a sec- ond binuclear center that accepts electrons from heme a and transfers them to O 2 bound to heme a 3.
700 Chapter 19 Oxidative Phosphorylation and Photophosphorylation
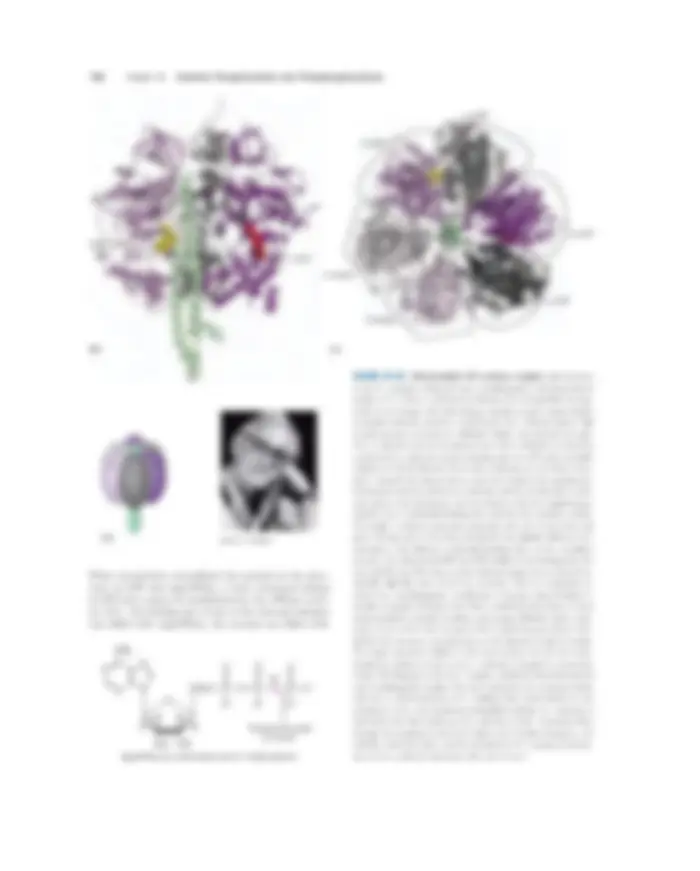
FIGURE 19–11 Cytochrome bc 1 complex (Complex III). The com- plex is a dimer of identical monomers, each with 11 different sub- units. (a) Structure of a monomer. The functional core is three sub- units: cytochrome b (green) with its two hemes ( b H and b L, light red); the Rieske iron-sulfur protein (purple) with its 2Fe-2S centers (yellow); and cytochrome c 1 (blue) with its heme (red) (PDB ID 1BGY). (b) The dimeric functional unit. Cytochrome c 1 and the Rieske iron-sulfur pro- tein project from the P surface and can interact with cytochrome c (not part of the functional complex) in the intermembrane space. The complex has two distinct binding sites for ubiquinone, QN and QP, which correspond to the sites of inhibition by two drugs that block oxidative phosphorylation. Antimycin A, which blocks electron flow from heme b H to Q, binds at QN, close to heme b H on the N (matrix) side of the membrane. Myxothiazol, which prevents electron flow from
QH 2 to the Rieske iron-sulfur protein, binds at QP, near the 2Fe-2S center and heme b L on the P side. The dimeric structure is essential to the function of Complex III. The interface between monomers forms two pockets, each containing a Q (^) P site from one monomer and a QN site from the other. The ubiquinone intermediates move within these sheltered pockets. Complex III crystallizes in two distinct conformations (not shown). In one, the Rieske Fe-S center is close to its electron acceptor, the heme of cytochrome c 1 , but relatively distant from cytochrome b and the QH 2 -binding site at which the Rieske Fe-S center receives elec- trons. In the other, the Fe-S center has moved away from cytochrome c 1 and toward cytochrome b. The Rieske protein is thought to oscil- late between these two conformations as it is reduced, then oxidized.
(a)
Intermembrane space (P side)
Matrix ( N side)
Cytochrome c 1
Cytochrome b
Rieske iron- sulfur protein
2Fe-2S
(b)
Cytochrome c 1
Cytochrome c
Rieske iron- sulfur protein
2Fe-2S center
Cytochrome b
(P side)
(N side)
b L
QPQP
QNQN b H
c 1
Heme
NAD�. In actively respiring mitochondria, the actions of many dehydrogenases keep the actual [NADH]/[NAD�] ratio well above unity, and the real free-energy change for the reaction shown in Equation 19–5 is therefore substantially greater (more negative) than �220 kJ/mol. A similar calculation for the oxidation of succinate shows that electron transfer from succinate ( E � for fumarate/succinate 0.031 V) to O 2 has a smaller, but still negative, standard free-energy change of about �150 kJ/mol.
702 Chapter 19 Oxidative Phosphorylation and Photophosphorylation
(a) (b)
FIGURE 19–13 Critical subunits of cytochrome oxidase (Complex IV). The bovine complex is shown here (PDB ID 1OCC). (a) The core of Complex IV, with three subunits. Subunit I (yellow) has two heme groups, a and a 3 (red), and a copper ion, CuB (green sphere). Heme a 3 and Cu (^) B form a binuclear Fe-Cu center. Subunit II (blue) contains two Cu ions (green spheres) complexed with the OSH groups of two Cys residues in a binuclear center, Cu (^) A, that resembles the 2Fe-2S cen- ters of iron-sulfur proteins. This binuclear center and the cytochrome
c –binding site are located in a domain of subunit II that protrudes from the P side of the inner membrane (into the intermembrane space). Subunit III (green) seems to be essential for Complex IV function, but its role is not well understood. (b) The binuclear center of Cu (^) A. The Cu ions (green spheres) share electrons equally. When the center is reduced they have the formal charges Cu 1 �Cu^1 �; when oxidized, Cu 1.5�Cu1.5�. Ligands around the Cu ions include two His (dark blue), two Cys (yellow), an Asp (red), and Met (orange) residues.
Subunit II Subunit I
4H +
4H+ (pumped)
Subunit III
4H+ (substrate)
4 e –
2H2O
O
4Cyt c
CuB
CuA
Fe-Cu center
a 3
a
Intermembrane space (P side)
Matrix ( N side)
FIGURE 19–14 Path of electrons through Complex IV. The three pro- teins critical to electron flow are subunits I, II, and III. The larger green structure includes the other ten proteins in the complex. Electron trans- fer through Complex IV begins when two molecules of reduced cy- tochrome c (top) each donate an electron to the binuclear center Cu (^) A. From here electrons pass through heme a to the Fe-Cu center (cy- tochrome a 3 and Cu (^) B). Oxygen now binds to heme a 3 and is reduced to its peroxy derivative (O 22 �) by two electrons from the Fe-Cu center. Delivery of two more electrons from cytochrome c (making four elec- trons in all) converts the O 22 �^ to two molecules of water, with con- sumption of four “substrate” protons from the matrix. At the same time, four more protons are pumped from the matrix by an as yet unknown mechanism.
Much of this energy is used to pump protons out of the matrix. For each pair of electrons transferred to O 2 , four protons are pumped out by Complex I, four by Com- plex III, and two by Complex IV (Fig. 19–15). The vec- torial equation for the process is therefore
NADH � 11H� N � 12 O 2 On NAD�^ � 10H� P � H 2 O (19–7)
The electrochemical energy inherent in this difference in proton concentration and separation of charge rep- resents a temporary conservation of much of the energy of electron transfer. The energy stored in such a gradi- ent, termed the proton-motive force, has two com- ponents: (1) the chemical potential energy due to the difference in concentration of a chemical species (H�) in the two regions separated by the membrane, and (2) the electrical potential energy that results from the separation of charge when a proton moves across the membrane without a counterion (Fig. 19–16). As we showed in Chapter 11, the free-energy change for the creation of an electrochemical gradient by an ion pump is
G RT ln (^) � � � Z � (19–8)
where C 2 and C 1 are the concentrations of an ion in two regions, and C 2 C 1 ; Z is the absolute value of its elec- trical charge (1 for a proton), and � is the transmem- brane difference in electrical potential, measured in volts. For protons at 25 C,
ln (^) � � 2.3(log [H�]P � log [H�]N ) 2.3(pHN � pH (^) P ) 2.3 pH
and Equation 19–8 reduces to
G 2.3 RT pH � � (19–9) (5.70 kJ/mol) pH � (96.5 kJ/V (^) � mol)∆ �
In actively respiring mitochondria, the measured ∆ � is 0.15 to 0.20 V and the pH of the matrix is about 0.
C 2 C 1
C 2 C 1
units more alkaline than that of the intermembrane space, so the calculated free-energy change for pump- ing protons outward is about 20 kJ/mol (of H�), most of which is contributed by the electrical portion of the electrochemical potential. Because the transfer of two electrons from NADH to O 2 is accompanied by the out- ward pumping of 10 H�^ (Eqn 19–7), roughly 200 kJ of the 220 kJ released by oxidation of a mole of NADH is conserved in the proton gradient. When protons flow spontaneously down their elec- trochemical gradient, energy is made available to do work. In mitochondria, chloroplasts, and aerobic bacte- ria, the electrochemical energy in the proton gradient drives the synthesis of ATP from ADP and Pi. We return to the energetics and stoichiometry of ATP synthesis driven by the electrochemical potential of the proton gradient in Section 19.2.
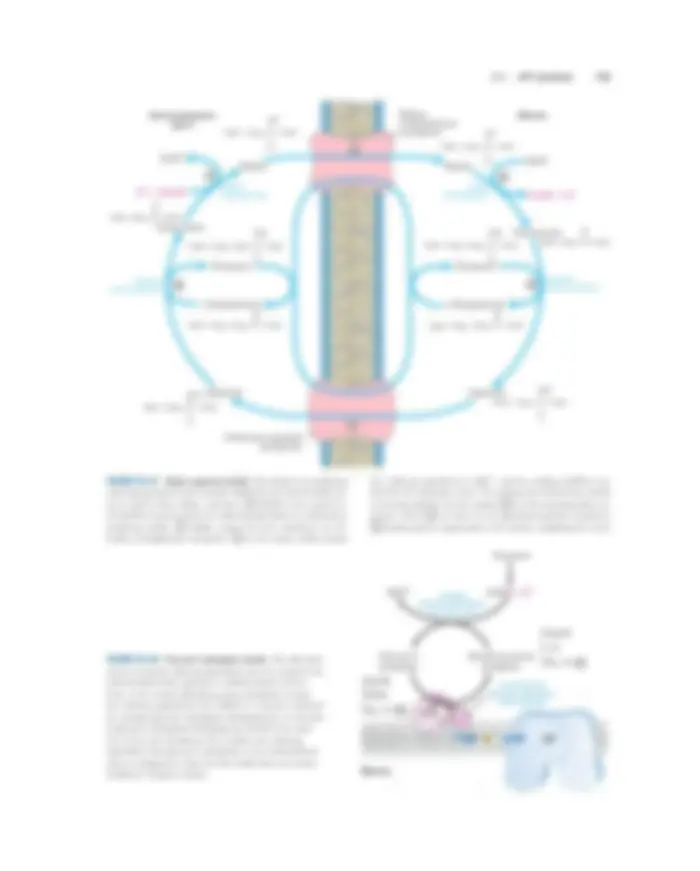
19.1 Electron-Transfer Reactions in Mitochondria 703
Intermembrane space (P side)
Matrix (N side)
4H+
(^1) – 2 O 2 +^ 2H
- (^) H Succinate Fumarate^2 O
4H+
II
NADH + H+^ NAD+
Cyt c
IV
2H +
III I
Q
FIGURE 19–15 Summary of the flow of electrons and protons through the four complexes of the respiratory chain. Electrons reach Q through Complexes I and II. QH 2 serves as a mobile carrier of elec- trons and protons. It passes electrons to Complex III, which passes them to another mobile connecting link, cytochrome c. Complex IV
then transfers electrons from reduced cytochrome c to O 2. Electron flow through Complexes I, III, and IV is accompanied by proton flow from the matrix to the intermembrane space. Recall that electrons from � oxidation of fatty acids can also enter the respiratory chain through Q (see Fig. 19–8).
N side [H�]N C 1 OH� OH� OH� OH� OH� OH� OH�
H� H� H� H� H� H� H�
P side [H�]P C 2
G RT ln ( C 2 / C 1 ) � Z ℑ �� 2.3 RT pH � ℑ ∆��
H�
Proton pump
� �
FIGURE 19–16 Proton-motive force. The inner mitochondrial mem- brane separates two compartments of different [H�], resulting in dif- ferences in chemical concentration ( pH) and charge distribution ( � ) across the membrane. The net effect is the proton-motive force ( G ), which can be calculated as shown here. This is explained more fully in the text.
Because the energy of substrate oxidation drives ATP synthesis in mitochondria, we would expect in- hibitors of the passage of electrons to O 2 (such as cyanide, carbon monoxide, and antimycin A) to block ATP synthesis (Fig. 19–18a). More surprising is the find- ing that the converse is also true: inhibition of ATP syn- thesis blocks electron transfer in intact mitochondria. This obligatory coupling can be demonstrated in isolated mitochondria by providing O 2 and oxidizable substrates, but not ADP (Fig. 19–18b). Under these conditions, no ATP synthesis can occur and electron transfer to O 2 does not proceed. Coupling of oxidation and phosphor- ylation can also be demonstrated using oligomycin or venturicidin, toxic antibiotics that bind to the ATP syn- thase in mitochondria. These compounds are potent in-
hibitors of both ATP synthesis and the transfer of elec- trons through the chain of carriers to O 2 (Fig. 19–18b). Because oligomycin is known to interact not directly with the electron carriers but with ATP synthase, it follows that electron transfer and ATP synthesis are obligately coupled; neither reaction occurs without the other. Chemiosmotic theory readily explains the depend- ence of electron transfer on ATP synthesis in mitochon- dria. When the flow of protons into the matrix through the proton channel of ATP synthase is blocked (with oligomycin, for example), no path exists for the return of protons to the matrix, and the continued extrusion of protons driven by the activity of the respiratory chain generates a large proton gradient. The proton-motive force builds up until the cost (free energy) of pumping
19.2 ATP Synthesis 705
NADH + H+^ NAD+
Succinate
Fumarate
Cyt c
ADP + Pi
ATP
4H+
4H+^
2H+
H+ Chemical potential ∆pΗ (inside alkaline)
ATP synthesis driven by proton-motive force
Electrical potential ∆w (inside negative)
O 2 + –
H 2 2H+
O 2
1
II
IV
I
III
Fo
F
Intermembrane space
Matrix
Q
FIGURE 19–17 Chemiosmotic model. In this simple representation of the chemiosmotic theory applied to mitochondria, electrons from NADH and other oxidizable substrates pass through a chain of carriers arranged asymmet- rically in the inner membrane. Electron flow is accompanied by proton transfer across the membrane, producing both a chemical gradient ( pH) and an electrical gradient ( � ). The inner mitochondrial membrane is imper- meable to protons; protons can reenter the matrix only through proton-specific channels (F (^) o). The proton-motive force that drives protons back into the matrix provides the energy for ATP synthesis, catalyzed by the F (^1) complex associated with F (^) o.
O 2 consumed
Add ADP � Pi Add succinate
(b) Time
ATP synthesized
Add venturicidin or oligomycin
Add DNP (^) Uncoupled
O 2 consumedAdd ADP � Pi
Add succinate
(a) Time
ATP synthesized
Add CN�
FIGURE 19–18 Coupling of electron transfer and ATP synthesis in mitochondria. In experiments to demonstrate coupling, mitochondria are suspended in a buffered medium and an O 2 electrode monitors O 2 consumption. At intervals, samples are removed and assayed for the presence of ATP. (a) Addition of ADP and Pi alone results in little or no increase in either respiration (O 2 consumption; black) or ATP synthe- sis (red). When succinate is added, respiration begins immediately and
ATP is synthesized. Addition of cyanide (CN�), which blocks electron transfer between cytochrome oxidase and O 2 , inhibits both respiration and ATP synthesis. (b) Mitochondria provided with succinate respire and synthesize ATP only when ADP and P (^) i are added. Subsequent ad- dition of venturicidin or oligomycin, inhibitors of ATP synthase, blocks both ATP synthesis and respiration. Dinitrophenol (DNP) is an un- coupler, allowing respiration to continue without ATP synthesis.
706 Chapter 19 Oxidative Phosphorylation and Photophosphorylation
BOX 19–1 THE WORLD OF BIOCHEMISTRY
Hot, Stinking Plants and Alternative Respiratory Pathways Many flowering plants attract insect pollinators by re- leasing odorant molecules that mimic an insect’s nat- ural food sources or potential egg-laying sites. Plants pollinated by flies or beetles that normally feed on or lay their eggs in dung or carrion sometimes use foul- smelling compounds to attract these insects. One family of stinking plants is the Araceae, which includes philodendrons, arum lilies, and skunk cab- bages. These plants have tiny flowers densely packed on an erect structure, the spadix, surrounded by a modified leaf, the spathe. The spadix releases odors of rotting flesh or dung. Before pollination the spadix also heats up, in some species to as much as 20 to 40 �C above the ambient temperature. Heat produc- tion (thermogenesis) helps evaporate odorant mole- cules for better dispersal, and because rotting flesh and dung are usually warm from the hyperactive me- tabolism of scavenging microbes, the heat itself might also attract insects. In the case of the eastern skunk cabbage (Fig. 1), which flowers in late winter or early spring when snow still covers the ground, thermogen- esis allows the spadix to grow up through the snow. How does a skunk cabbage heat its spadix? The mitochondria of plants, fungi, and unicellular eukary- otes have electron-transfer systems that are essen- tially the same as those in animals, but they also have an alternative respiratory pathway. A cyanide- resistant QH 2 oxidase transfers electrons from the ubiquinone pool directly to oxygen, bypassing the two proton-translocating steps of Complexes III and IV (Fig. 2). Energy that might have been conserved as
ATP is instead released as heat. Plant mitochondria also have an alternative NADH dehydrogenase, insen- sitive to the Complex I inhibitor rotenone (see Table 19–4), that transfers electrons from NADH in the ma- trix directly to ubiquinone, bypassing Complex I and its associated proton pumping. And plant mitochon- dria have yet another NADH dehydrogenase, on the external face of the inner membrane, that transfers electrons from NADPH or NADH in the intermem- brane space to ubiquinone, again bypassing Complex I. Thus when electrons enter the alternative respira- tory pathway through the rotenone-insensitive NADH dehydrogenase, the external NADH dehydrogenase, or succinate dehydrogenase (Complex II), and pass to O 2 via the cyanide-resistant alternative oxidase, en- ergy is not conserved as ATP but is released as heat. A skunk cabbage can use the heat to melt snow, pro- duce a foul stench, or attract beetles or flies.
FIGURE 1 Eastern skunk cabbage.
FIGURE 2 Electron carriers of the inner membrane of plant mitochondria. Electrons can flow through Complexes I, III, and IV, as in animal mitochondria, or through plant-specific alterna- tive carriers by the paths shown with blue arrows.
Heat
I IV
Intermembrane space
Q
Matrix
Cyt c
NAD+
NAD(P)+^
External NAD(P)H dehydrogenase
Alternative oxidase
Alternative NADH dehydrogenase
III
NADH^ H 2 O
NAD(P)H
(^1) – 2 O 2
ATP Synthase Has Two Functional Domains, Fo and F (^1)
Mitochondrial ATP synthase is an F-type ATPase (see Fig. 11–39; Table 11–3) similar in structure and mech- anism to the ATP synthases of chloroplasts and eubac- teria. This large enzyme complex of the inner mito- chondrial membrane catalyzes the formation of ATP from ADP and Pi, accompanied by the flow of protons from the P to the N side of the mem- brane (Eqn 19–10). ATP syn- thase, also called Complex V, has two distinct components: F 1 , a peripheral membrane protein, and Fo ( o denoting oligomycin-sensitive), which is integral to the membrane. F 1 , the first factor recognized as essential for oxidative phos- phorylation, was identified and purified by Efraim Racker and his colleagues in the early 1960s. In the laboratory, small membrane vesicles formed from inner mitochondrial membranes carry out ATP syn- thesis coupled to electron transfer. When F 1 is gently extracted, the “stripped” vesicles still contain intact res- piratory chains and the Fo portion of ATP synthase. The vesicles can catalyze electron transfer from NADH to O 2
but cannot produce a proton gradient: Fo has a proton pore through which protons leak as fast as they are pumped by electron transfer, and without a proton gra- dient the F 1 -depleted vesicles cannot make ATP. Iso- lated F 1 catalyzes ATP hydrolysis (the reversal of syn- thesis) and was therefore originally called F 1 ATPase. When purified F 1 is added back to the depleted vesicles, it reassociates with Fo , plugging its proton pore and restoring the membrane’s capacity to couple electron transfer and ATP synthesis.
ATP Is Stabilized Relative to ADP on the Surface of F 1 Isotope exchange experiments with purified F 1 reveal a remarkable fact about the enzyme’s catalytic mecha- nism: on the enzyme surface, the reaction ADP � P (^) i ATP � H 2 O is readily reversible—the free-energy change for ATP synthesis is close to zero! When ATP is hydrolyzed by F 1 in the presence of 18 O-labeled water, the P (^) i released contains an 18 O atom. Careful meas- urement of the 18 O content of P (^) i formed in vitro by F 1 - catalyzed hydrolysis of ATP reveals that the P (^) i has not one, but three or four 18 O atoms (Fig. 19–21). This in- dicates that the terminal pyrophosphate bond in ATP is cleaved and re-formed repeatedly before P (^) i leaves the enzyme surface. With P (^) i free to tumble in its binding site, each hydrolysis inserts 18 O randomly at one of the
y^ z
708 Chapter 19 Oxidative Phosphorylation and Photophosphorylation
Efraim Racker, 1913–
(^18) O
(^18) O
ADP
ATP � H 218 O
�
(^18) O P (^18) O
Enzyme (F 1 )
(a)
ADP
a-Arg^376
b-Arg^182
b-Glu^181 b-Lys 155
Mg2+
FIGURE 19–21 Catalytic mechanism of F 1. (a)^18 O-exchange exper- iment. F 1 solubilized from mitochondrial membranes is incubated with ATP in the presence of 18 O-labeled water. At intervals, a sample of the solution is withdrawn and analyzed for the incorporation of 18 O into the Pi produced from ATP hydrolysis. In minutes, the Pi contains three or four 18 O atoms, indicating that both ATP hydrolysis and ATP synthesis have occurred several times during the incubation. (b) The likely transition state complex for ATP hydrolysis and synthesis in ATP
synthase (derived from PDB ID 1BMF). The � subunit is shown in green, � in gray. The positively charged residues � -Arg 182 and � -Arg 376 coordinate two oxygens of the pentavalent phosphate intermediate; � - Lys^155 interacts with a third oxygen, and the Mg 2 �^ ion (green sphere) further stabilizes the intermediate. The blue sphere represents the leav- ing group (H 2 O). These interactions result in the ready equilibration of ATP and ADP � Pi in the active site.
(b)
four positions in the molecule. This exchange reaction occurs in unenergized Fo F 1 complexes (with no proton gradient) and with isolated F 1 —the exchange does not require the input of energy. Kinetic studies of the initial rates of ATP synthesis and hydrolysis confirm the conclusion that G � for ATP synthesis on the enzyme is near zero. From the meas- ured rates of hydrolysis ( k 1 10 s�^1 ) and synthesis ( k � 1 24 s�^1 ), the calculated equilibrium constant for the reaction
Enz-ATP Enz–(ADP � Pi )
is
K �eq 2.
From this K �eq, the calculated apparent G � is close to zero. This is much different from the K eq� of about 10 5 ( G � �30.5 kJ/mol) for the hydrolysis of ATP free in solution (not on the enzyme surface). What accounts for the huge difference? ATP syn- thase stabilizes ATP relative to ADP � Pi by binding ATP more tightly, releasing enough energy to counterbalance the cost of making ATP. Careful measurements of the binding constants show that Fo F 1 binds ATP with very high affinity ( K d ≤ 10 �^12 M) and ADP with much lower affinity ( K d ≈ 10 �^5 M). The difference in K d corresponds to a difference of about 40 kJ/mol in binding energy, and this binding energy drives the equilibrium toward for- mation of the product ATP.
The Proton Gradient Drives the Release of ATP
from the Enzyme Surface
Although ATP synthase equilibrates ATP with ADP � Pi, in the absence of a proton gradient the newly syn- thesized ATP does not leave the surface of the enzyme.
24 s�^1 10 s�^1
k � 1 k 1
zy
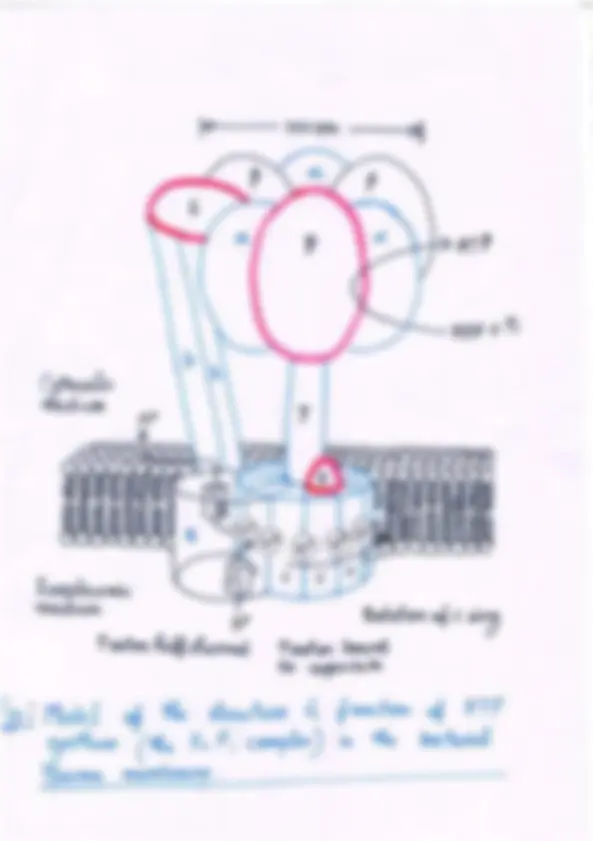
It is the proton gradient that causes the enzyme to re- lease the ATP formed on its surface. The reaction co- ordinate diagram of the process (Fig. 19–22) illustrates the difference between the mechanism of ATP synthase and that of many other enzymes that catalyze ender- gonic reactions. For the continued synthesis of ATP, the enzyme must cycle between a form that binds ATP very tightly and a form that releases ATP. Chemical and crystallo- graphic studies of the ATP synthase have revealed the structural basis for this alternation in function.
Each � Subunit of ATP Synthase Can Assume Three Different Conformations Mitochondrial F 1 has nine subunits of five different types, with the composition � 3 � 3 ���. Each of the three � subunits has one catalytic site for ATP synthesis. The crystallographic determination of the F 1 structure by John E. Walker and colleagues revealed structural de- tails very helpful in explaining the catalytic mechanism of the enzyme. The knoblike portion of F 1 is a flattened sphere, 8 nm high and 10 nm across, consisting of al- ternating � and � subunits arranged like the sections of an orange (Fig. 19–23a–c). The polypeptides that make up the stalk in the F 1 crystal structure are asymmetri- cally arranged, with one domain of the single � subunit making up a central shaft that passes through F 1 , and another domain of � associated primarily with one of the three � subunits, designated � -empty (Fig. 19–23c). Although the amino acid sequences of the three � sub- units are identical, their conformations differ, in part because of the association of the � subunit with just one of the three. The structures of the � and � subunits are not revealed in these crystallographic studies. The conformational differences among � subunits extend to differences in their ATP/ADP-binding sites.
19.2 ATP Synthesis 709
G
(kJ/mol)
Reaction coordinate
80
60
40
20
0
‡
P ADP�Pi ES
E � S
E ADP�Pi
[E ATP]
Typical enzyme ATP synthase
ATP (in solution)
FIGURE 19–22 Reaction coordinate diagrams for ATP synthase and for a more typical enzyme. In a typical enzyme-catalyzed reaction (left), reaching the transition state (‡) between substrate and product is the major energy barrier to overcome. In the reaction catalyzed by ATP synthase (right), release of ATP from the enzyme, not formation of ATP, is the major energy barrier. The free-energy change for the formation of ATP from ADP and Pi in aqueous solution is large and positive, but on the enzyme surface, the very tight binding of ATP provides sufficient binding energy to bring the free energy of the enzyme-bound ATP close to that of ADP � Pi, so the reaction is readily reversible. The equilibrium constant is near 1. The free energy required for the release of ATP is provided by the proton-motive force.