Download Autoshaping in Pigeons: The Role of Visual and Oral Stimuli and more Lecture notes Topography in PDF only on Docsity!
JOURNAL OF THE EXPERIMENTAL ANALYSIS OF BEHAVIOR
THE ASSOCIATIVE RELATION UNDERLYING
AUTOSHAPING IN THE^ PIGEON'
GuY WOODRUFF AND D. R. WILLIAMS
UNIVERSITY OF PENNSYLVANIA
Fifteen pigeons were exposed to either response-independent or response-dependent sched- ules of water reinforcement, whereby water was injected directly into the unrestrained pigeons' mandibles. Key-contact responses were released by a lighted key correlated-with water, but not by a lighted key uncorrelated with water. A negative response-reinforcer contingency suppressed autoshaped^ key-contact^ responses,^ resulting^ in^ responding^ directed away from the^ lighted key.^ In^ all^ pigeons,^ water^ injected^ directly^ into^ the mandibles elicited a consummatory fixed-action pattern of^ "mumbling" and^ swallowing.^ The^ lighted key correlated with water released a^ broader^ set of both^ appetitive^ and^ consummatory responses: approach to the lighted key, "bowing", "rooting", "mumbling", and swallowing. Key-contact responses were "rooting" and "mumbling" motions of the beak on^ the^ sur- face of the key. Views of autoshaping based on stimulus substitution or stimulus sur- rogation do not fully explain the origin of autoshaped responses not previously elicited by the reinforcer. The present findings are consonant with views of conditioning that em- phasize the large degree of biological pre-organization in conditioned response patterns, and the importance of associative factors in the control of such patterns. Key words: autoshaping, respondent conditioning, stimulus substitution, stimulus^ sur- rogation, "learned release", water reinforcer, species-specific behavior, key peck, pigeon
The phenomenon of^ autoshaping entails conditioning the pigeon's key peck with a re- spondent conditioning procedure. Correlation of a lighted key with grain presentations gen- erally produces key pecking (Brown and Jen- kins, 1968; Gamzu and Williams, 1971). The stimulus (lighted key)-reinforcer (grain) rela- tionship exerts^ powerful^ control^ over^ the de- velopment of^ key pecking when^ reinforcers are independent of responding (Gamzu and Schwartz, 1973; Gamzu and Williams, 1973), or dependent on not responding (Barrera, 1974; Griffin and Rashotte, 1973; Schwartz, 1972; Williams and Williams, 1969). The na- ture of the stimulus-reinforcer relation in au- toshaping was the concern of the present study. One view of autoshaping attributes the ori-
1This research was supported by NSF Grant GB 35319X to D. R. Williams. Some of the data were re- ported by the first author at the meeting of the Eastern Psychological Association, 1974. Thanks are due Doug- las Fenner, Robert Morrison, Jan (^) Moselsky, Mark Starr, and James Stellar for helpful discussions of the data. Reprints may be obtained from Guy Woodruff, De- partment of^ Psychology, 3815 Walnut^ Street, University of (^) Pennsylvania, Philadelphia, Pennsylvania 19174.
gin of key pecking to stimulus substitution
(Jenkins and^ Moore, 1973; Moore,^ 1971;^ Stad-
don and Simmelhag, 1971). According to a
stimulus. substitution principle, stimulus con-
trol of some type of response is transferred
from an unconditioned stimulus (US) to a con- ditioned stimulus (CS). In autoshaping, the particular conditioned response generally re- sembles the response exhibited during rein-
forcement. This correspondence between con-
ditioned and unconditioned responding holds
across several species (pigeons, chickens, quail,
fish, rats, dogs), responses, and reinforcers
used in autoshaping situations (Hearst and
Jenkins, 1974; Moore, 1973; Schwartz^ and
Gamzu, in^ press).
An experiment by Jenkins and Moore (1973)
is the clearest support for a stimulus substitu- tion account of autoshaped key pecking with pigeons. During autoshaping with grain rein-
forcement, pigeons directed pecks to the grain
(US) in the food magazine and toward a
lighted key (CS) paired^ with^ grain.^ Likewise,
during autoshaping with^ water^ reinforcement pigeons directed drinking responses to the
water (US) in the magazine and toward a
lighted key (CS) paired with water. Jenkins
1976, 26,^ 1-13^ NUMBER^ I^ (JULY)
GUY WOODRUFF and D. R. WILLIAMS
and Moore interpreted these results in terms of stimulus substitution, suggesting that the CS gained control of the type of response (peck or drink) found to be elicited by the grain or water US. These authors offered the term "stimulus surrogation" as an even better description of the results. Their pigeons re- sponded to the CS as though it were the US, i.e., the lighted key became a surrogate for either grain or water in the magazine. Stimulus substitution or surrogation sug- gests that approach and contact responses to the CS develop because the reinforcer elicits such responses. However, approach and con- tact responses are also inherent requirements for acquisition of the reinforcer in the con- ventional procedure of delivering grain or water in a magazine or dipper. These re- sponses are typically well established during
"magazine training" before conditioning ses-
sions are initiated. Is the occurrence of these responses necessary for the development of autoshaped approach and contact responses to the lighted key, as substitution or surrogation principles would suggest?
Williams' (cf. Schwartz and Gamzu, in press)
"learned release" account of autoshaped key
pecking suggests not. In this view, the associa-
tive relation underlying autoshaping involves the visual stimulus of the lighted key and the interoceptive oral stimuli arising from grain or water in the mandibles. Approach and con- tact responses to the sight of grain, or water, or the lighted key paired with either grain or water, are controlled by the correlation be- tween each visual stimulus and an oral rein-
forcing stimulus. Normally, approach and
pecking or drinking responses arrange and
maintain a correlation between visual and
oral stimuli during feeding in the home cage
as well as during reinforcement in the condi- tioning apparatus. However, any other "arti- ficial" way of arranging a correlation between
visual and oral stimuli should also^ produce
and maintain approach and^ contact responses to the visual stimulus. By this view, autoshap- ing with grain or water introduced directly into the mandibles, bypassing approach and
contact responses to the reinforcer, should be
just as effective as autoshaping with grain or
water presented in a magazine. Since the
lighted key acquires its^ releasing properties
solely from its^ correlation^ with^ oral^ reinforc-
ing stimuli, the associative relation established
by the autoshaping procedure may be said to instigate, rather than transfer, control of re- sponding to the lighted key. The following questions about this issue over the nature of the role of the stimulus-re- inforcer relationship in autoshaping were ad- dressed by the present experiment. (1) Can water introduced directly into the mandibles of a^ pigeon be a^ reinforcer in^ the autoshaping procedure? (2) If so, what responses are elicited by the oral stimulus of water, and by the lighted key with which it is paired? (3) If au-
toslhaped responding occurs, may its develop-
ment be described as stimulus substitution? That is, does the visual stimulus gain control of responses previously controlled by the oral stimulus? May the results be described as stim- ulus surrogation? Does the pigeon respond to the visual stimulus as though it were the oral stimulus? Autoshaping, omission training, and ran- dom control procedures were investigated with a (^) new method of delivering water (^) to pi- geons. Water was injected through a cannula directly into the mandibles of water-deprived pigeons. The data of primary interest were the effects of this method of correlating visual and oral stimuli on the development of condi- tioned responding to the visual stimulus.
METHOD
Subjects Eleven White King and four White Car- neaux adult pigeons, experimentally naive at the start of the experiment, were maintained at 80% of their free-feeding body weights by restriction of daily water rations. Grain and grit were always available in the home (^) cages.
Apparatus A modified Lehigh Valley operant condi- tioning chamber (31 by 35 by 33 cm) for pi- geons was used. Two 2.4-cm diameter response
keys, 7.5 cm apart and 25.5 cm above the floor,
protruded 2 mm^ into^ the chamber from^ the
stimulus panel. A 7-W red Westinghouse
Christmas (^) Bulb (D (^) 18-71/2) transilluminated the right-hand key, the only one used. The re- sponse key required 15 g force (0.15 N) to op- erate a photocell attached to the back of the
stimulus panel and record a key-contact re-
sponse. Horizontal key displacement during
GUY WOODRUFF and D. R. WILLIAMS
currence of a darkened key during water injec- tions was eliminated by illuminating the key during US presentations. The procedure was otherwise identical to that for the first auto- shaping group. Thus, each illumination of the
key lasted 12 sec and overlapped the reinforcer
(Schwartz, 1972 used a similar autoshaping procedure). Two birds (P9 and^ P1) received
10 sessions; the other two (PlO and P12) re-
ceived five sessions of the stimulus-overlap procedure, followed by five sessions of the first autoshaping procedure. (3) Omission training: three pigeons (P13, P14, and P15) were exposed to 20 or 25 sessions of an omission-training procedure (Schwartz, 1972). The procedure was the same as that for the first group, except for the addition of a negative response-reinforcer contingency. A re- sponse during illumination of the key omitted the US scheduled to occur at the end of that CS. (^) Key-contact responses had no other sched- uled (^) consequences. (4) Associative control: four pigeons (P5,
P6, P7, P8) were exposed to 10 sessions of an
associative-control procedure (Gamzu and
Williams, 1971) in which illumination of the
key and reinforcement (US) were uncorre- lated. Key illumination occurred according to the same temporal schedule as that described for the first autoshaping group, but reinforc- ers occurred randomly throughout sessions. Water injections were delivered by the output of a probability generator set at p =^ 0.09 and sampled once every 6 sec. About 50 reinforcers were delivered each session. Lighted key and reinforcer could, and did, occasionally occUr together. Each pigeon in all four groups was observed by the first author^ during each of the first^ two or three sessions, to observe what responses were elicited by water in the mandibles.
RESULTS
Autoshaping Group
Figure 1 shows the proportion of CSs in which there was a key-contact response during
each acquisition, extinction, and reacquisition
session for each bird in the first group. Key- contact responses were autoshaped in all four birds. Mean number of CSs to the first re- sponse was 39 (range: 15 to 75) and each bird's
first response occurred during a key illumina-
tion. Responding steadily increased to more than 0.8 of the CSs by the third or fourth ses- sion. Responses per minute in Session 4 dur- ing the CS and the ISI were, respectively: P (31.0, 41.9), P2(92.2, 11.7), P3(70.6, 11.3), and P4(131.0, 34.9). Extinction began with Session 5, and re- sponding decreased quickly and ceased within eight extinction^ sessions^ for^ three^ birds^ (cf. Figure 1).^ P4^ continued^ to^ respond^ during^ sev-
eral CSs per session for many sessions, but
eventually met the extinction criterion after 25 sessions. Responding was quickly regained in P3 and
P4 dturing reacquisition, but in P3 remained
at only a moderate level of 0.5 of the CSs by the fourth reacquisition session (cf. Figure 1). Rates of response during the ISI were un- usually high for autoshaping in the first group of birds. Indeed, for P1, responding was greater during the ISI than during the CS. Observation of the birds showed an unantici- pated outcome of the present technique of re- inforcer delivery, and provided a clue to the source of the high ISI response rates. Key-con- tact responses first occurred during key illumi- nations, but soon began to occur during injec- tions of water. If responding was initiated during a CS, it tended to persist after the light was extinguished while water was injected into the mandibles. By the fourth session, each pi- geon consistently consumed reinforcers while operating the darkened key. This behavior was not anticipated by the experimenters, and
responses were unfortunately partitioned into
only two categories by the recording equip- ment, those occurring during the illuminated key (CS) and those occurring during the dark- ened key (ISI). Possibly, high rates of response in the ISI were an artifact produced by includ- ing responses during the US in calculating ISI response rates. However, the occurrence of responding to the dark key during water injections only par- tially accounts for the present findings. Each bird also tended to respond throughout the ISI by Session 4. Perhaps consumption of water while responding to the darkened key increased the tendency to respond to the dark key at^ all times. The second autoshaping pro- cedure was designed to test this hypothesis. The occurrence of a dark key and consump- tion of water was eliminated by illuminating the key during water injections.
ASSOCIATIVE RELATION OF AUTOSHAPING
1.0.
X.6-
.4.
.2-
On
1.0-
.8-
.6-
.4a
.2'
O
Pi P2 P
I A'.e.SJA
2 4 6 8 10 12 2 4 6 8 2 4 6 8 10 12 14 16
V~~~~~~~~~~~~~~~~~P
(^2 4 6 8 10 12) 14 16 18 20 22 24 26 28 30 32 34 SESSION NUMBER O_ CONDITIONING 0-0 EXTINCTION Fig. 1. Proportion of CS presentations with at least one key-contact response during autoshaping for each bird in the first group. Solid lines indicate acquisition, or reacquisition sessions (P3 and P4). Dotted lines indicate ex- tinction sessions.
Autoshaping with (^) Stimulus-Overlap Group Figure 2 shows the proportion of^ CSs in which there was^ a key-contact response for each bird in the second group. Key-contact re- sponses were autoshaped in all four birds. Mean number of CSs to the first response was 43 (range: 8 to 75) and the first response oc- curred during a key illumination for each bird. Responding steadily increased to a (^) high level for three birds, but showed a more grad- ual increase for (^) P12. Key-contact responses were partitioned into three categories for this group of birds: those occurring during the first 6 sec of key illumi- nation (the CS), those occurring during injec- tion of water while the key remained illumi- nated, and those occurring during the ISI while the key was dark. (^) Responses per minute for (^) each of the birds during Session 4 in
each of these three categories were, respec- tively: P9(38.0, 20.4, 1.1), P10(83.6, (^) 72.6, 14.1), P11(216.7, 282.9, 0.8), and P12(89.6, 123.5, 0.9). By Session 4, with birds in both the auto- shaping and the stimulus-overlap groups hav- ing each experienced the same number of CS and US presentations, (^) rates of response in the CS showed^ no marked difference between groups. In contrast, responding during the ISI was much less in the stimulus-overlap group. Only PlO showed substantial responding dur- ing the ISI in the second group. Figure 3 pre- sents examples of cumulative records for one bird in each (^) group and allows (^) comparison of the (^) patterns of (^) response generated by the two autoshaping procedures. The pattern of re- sponding found with the first autoshaping procedure was irregular. Bursts of responding occurred throughout the ISI as well as the CS.
ASSOCIATIVE RELATION OF AUTOSHAPING
. *,,,F , .J, .... .a. F
Fig. 3. Cumulative records of two pigeons' autoshaped contact responses to the lighted key. The lower event marker indicates^ CS^ presentations.^ The^ upper^ event^ marker^ located^ on^ the cumulative^ record itself^ indicates^ US presentation. The record^ on^ the^ left^ shows^ a^ portion^ of^ Session 4^ for^ P3^ in^ the^ first^ autoshaping^ group.^ The overall pattern of responding is irregular, with^ many responses occurring during the^ ISI^ as^ well^ as^ during^ the CS and US. The record on the right shows a portion of Session 4 for Pll in the stimulus-overlap group. The pat- tern of responding was very regular, with virtually all responses occurring during the^ CS^ and US.
during illumination^ of^ the key^ and^ persisting
afterward during the dark^ key at^ the^ begin-
ning of the ISI. Figure 4 shows the number of bursts during the dark key for each bird^ in^ the first and second groups. Birds in the first auto-
shaping group showed high levels of respond-
ing, which tended to increase over sessions. Birds in the stimulus-overlap group showed low (^) levels of responding, which tended to de-
crease over sessions. By Session 4, there^ was no
overlap between groups in^ levels of^ respond-
ing. All four pigeons in the first group far ex-
ceeded the highest levels of responding in the
second group. Furthermore, PlO and P12 did
not develop substantial responding during the
dark (^) key when switched from the stimulus- overlap to the first^ autoshaping procedure.
Responding during the dark key found in^ the
first group was apparently specific to the first
autoshaping procedure during initial acquisi-
tion of key-contact responding. Thus, the high
levels of ISI responding in the first group may
be attributable^ to an^ increased^ tendency^ to re-
spond to^ the^ dark^ key, produced by consuming
reinforcers during contact with^ the dark^ key.
Omission-Training Group
Figure 5 shows the proportion of CSs in
which there was a key-contact response for
each pigeon in the omission-training proce-
dure. Key-contact responses were autoshaped
in all three birds. Mean number of trials^ to the first response was 71 (range: 69 to 73). The steady increase in responding found with the first two autoshaping groups was not evi-
denced by this group. Responding gradually
increased to^ moderate^ or low levels,^ and^ then
declined to very low levels in all birds. After
prolonged exposure to the negative response-
reinforcer contingency, the three birds seldom
operated the key. Rather, these pigeons exhib-
ited responses directed away from the illumi- nated key to adjacent areas of the stimulus
panel, Plexiglas door, or floor. P15 adopted a
response that did not operate the key, by mak- ing contact responses to the 2-mm wide edge of the key. Beginning with Session 14, the key was recessed for this bird and Figure 5 shows the immediate increase in recorded respond-
ing. Subsequently,^ responding^ declined^ to low
GUY WOODRUFF and D. R. WILLIAMS
P
P
I
P
P
0
-U-U-
P
P
__
SESSION NUMBER
* APPARATUS
FAILURE
* AUTOSHAPING
PROCEDURE
Plo
P
STIMULUS OVERLAP
PROCEDURE
Fig. 4. Number of bursts of responding on the key during the ISI for birds in the first and second autoshap- ing procedures plotted across sessions. On the left are the data for the birds in the first (^) group. On the (^) right are the data for the stimulus-overlap group. Solid bars indicate sessions under the first (^) autoshaping procedure. Hatched bars on the right indicate sessions under the (^) stimulus-overlap procedure. Notice that PlO and (^) P12 were switched to the first (^) procedure after Session (^) 5, shown at the far (^) right of the (^) figure.
levels as responses were directed away from the key, as was the case for P13 and P14. In contrast to (^) previous reports of omission training with grain (Williams and (^) Williams, 1969), key-contact responses autoshaped with the present water reinforcement technique showed a marked sensitivity to the negative response-reinforcer contingency.
Associative-Control Group
Key-contact responding was not generated by the^ associative-control procedure. One^ of the birds (P5) struck the key during an^ ISI once. By the end of 10 sessions, two birds^ con- sumed reinforcers while rooting on the floor of the chamber, and the other two birds re- mained relatively inactive and merely swal- lowed each reinforcer.
Topography of Water-Associated Responses Observation of all 15 birds showed that swallowing and "mumbling" (Moore, 1971) responses were always elicited by water in the mandibles. Peristaltic movements of the throat (swallowing) were (^) accompanied by rapid, rhythmic (four to seven per second) opening and closing movements of the beak (mum- bling). This pattern is the pigeon's species-spe- cific consummatory response for water (Craig, 1912; Lorenz, 1937). In all (^11) birds in the first three groups, responses (^) released by the illuminated key in- cluded mumbling and swallowing, but con- sisted of a broader set of water-associated responses. The complete species-specific drink- ing pattern in pigeons includes "bowing",
CO c-
cnm 100
z 2 cn w cc (^0)
ob 200-
w
z
100a
Doi 0
__ i 0 -
.
m
a
GUY WOODRUFF and D. R. WILLIAMS
A
B
C
D
Csu
Cs I I
Cs us
CS
LL C,S (^) I
Is CS US
~~~~~I.
60 GRAMS-
r \ ~~~~~~L * a_z_
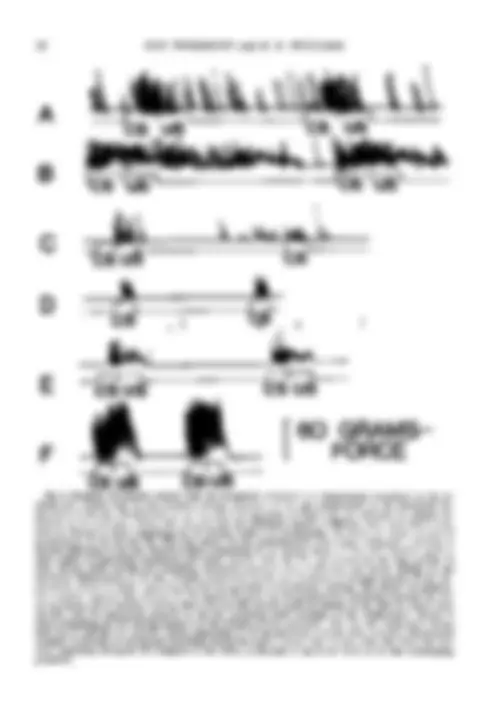
Csus Csus Fig. 6. Examples of response records from the dynograph connected to a displacement transducer on the re- sponse key. Contact with the key produced vertical (^) excursions of the pen proportional to the horizontal dis- placement of^ the key. Illuminations of the key (CS) and injections of water (US) are indicated by upward de- flections of the event pen. Notice that the time base for individual records is different: each event lasted 6 sec, however. Record A shows responding for P1 during Session 4 of autoshaping. The event pen shows two paired presentations of the CS and US. This bird struck the key predominantly with a peck topography, resulting in discrete deflections of the pen. Record B shows responding for P4 during Session 4. This bird showed a (^) pattern more typical of responding autoshaped with water. Contact with the key was maintained for long (^) periods of time, during which rooting and mumbling movements produced sharp (^) deflections and slower changes in the horizontal (^) displacement of the key. Pressure (^) maintained on the key produced an envelope pattern of key dis- placement. Record C shows^ responding for P13 during Session 3 of omission training. The second CS presenta- tion contains responses that omitted the US. Record D shows two CS presentations containing responses for P during Session 18 of omission training. Both P13 and P14 showed rapid movements of the beak in contact with the key; only P13 maintained pressure on the key, producing small envelopes of key displacement. Record E shows responding for P12 during Session 4 of the (^) stimulus-overlap procedure. The key was (^) illuminated during both the CS and the US. Record F shows (^) responding for P9 (^) during Session 3 of the same (^) procedure. Responding consisted (^) exclusively of (^) rooting and (^) mumbling during maintained contact with the key. Note that there was very little (^) responding during the ISI compared to that (^) shown in Records A and B for birds in the first autoshaping procedure.
I
ASSOCIATIVE RELATION OF A UTOSHAPING
sponse (^) topography found with (^) the present pro- cedures. Topography was similar to that illus- trated in Jenkins and Moore's (1973) Figure 4
(p. 177). Nine of the 11 birds showed only
rooting and mumbling topographies; two birds (P1 in the first group and Pll in the
second group) struck the key forcefully with a
ballistic-peck topography. Only the rapid
mumbling motions that occasionally appeared
on the pen tracings distinguished these birds'
topography from that of food pecks (Jenkins
and Moore, 1973). The four birds in the associative-control group also occasionally showed rooting and bowing responses, but in each case (^) they were directed to the floor of the chamber. These
responses were^ observed^ infrequently in^ the
first few sessions, and did not occur consist-
ently during or immediately after water injec-
tions. They were not elicited by water in the
mandibles, but rather appeared to develop in
the present situation over several sessions.
DISCUSSION
A major finding of the present experiment
is that water introduced directly into the man-
dibles of pigeons was an^ effective reinforcer
for autoshaping contact responses to^ a^ lighted
key correlated with water. Three general char-
acteristics of autoshaping were evident in the
present data. (1) Responding emerged after a
typical number of pairings of the lighted key
and water (mean = 49 pairings; range = 8 to
76 pairings) for the 11 birds in the two auto-
shaping groups and the omission training
group. (2) Once responding emerged, substan-
tial rates of responding quickly developed for
birds in the two autoshaping groups. (3) Re-
sponding did^ not^ develop in^ the control group
with uncorrelated presentations of the lighted
key and water. Two aspects of the present results differed
markedly from previous reports of autoshap-
ing and omission training. First, in the omis-
sion-training group, the negative response-
reinforcer contingency suppressed autoshaped
contact responses to the key. Bowing and^ root-
ing responses were directed toward the floor
and parts of the stimulus panel near the key
as responding directed toward the lighted key
diminished. This finding is at odds with some
studies of omission training with grain
reinforcement, which showed substantial re-
sponding to the key in spite of the negative contingency (Schwartz, 1972; Williams and Williams, 1969). However, Barrera (1974) found that a substantial amount of responding may be directed away from the key during omission training with grain reinforcement. (^) In our laboratory, we often (^) find that bowing and rooting responses (^) in the water magazine tend to replace autoshaped contact responses to the key during extended exposure to omission training with water. A second difference was the unusually high level of ISI responding generated by the first autoshaping procedure. Previous studies of autoshaping (Brown and Jenkins, 1968; Gamzu and (^) Williams, 1971; Jenkins and (^) Moore, 1973) have typically found very little responding (^) during the ISI, although there are instances of (^) substantial responding during the ISI in the literature (Gamzu and Schwartz, 1973; Perkins, Beaver, Hancock, Hemmendinger, Hemmendinger, and Ricci, 1975). In the present experiment, this effect is attributable to the occurrence of responding on the dark key during reinforcement in the first procedure, and not to the water reinforce- ment technique, per se. The (^) second procedure (stimulus (^) overlap) eliminated the occurrence of (^) responding to the dark key during rein- forcement, and effectively eliminated ISI re- sponding. Thus, neither the suppression of re- sponding by the negative response-reinforcer contingency, nor the generation of significant levels of ISI responding, alter the conclusion that water introduced directly into the mandi- bles is an effective reinforcer for autoshaping. A further finding of (^) importance concerns
the particular responses controlled by the
lighted key and reinforcer in the present pro- cedures. The oral stimulus of water in the mandibles reliably elicited mumbling and
swallowing consummatory responses. The
lighted key paired with water controlled a
more complex set of conditioned behaviors comprising the pigeon's (^) complete species-spe- cific (^) drinking pattern. Approach to the lighted key, (^) bowing in a downward direction, rooting on^ the surface of the key, mumbling
motions of the mandibles, and swallowing mo-
tions of the throat were observed during the CS. Contact responses autoshaped to the lighted key consisted of a subset of the com- plete pattern: rooting and mumbling motions.
Notably, both^ the^ complete set of conditioned
responses, as well as the (^) subset consisting of
ASSOCIATIVE RELATION OF AUTOSHAPING 13
organized responses by stimuli that^ precede reinforcement, rather than by the reinforcer itself, is made clear by experiments such as Wasserman's (1973) and the present one. "Learned release" suggests that the origin of autoshaped "preparatory" responses lies in the selection and instigation of components of the organism's species-specific^ behavior reper- toire by associative learning factors.
REFERENCES
Barrera, F. J. Centrifugal selection^ of^ signal-directed pecking. Journal of the^ Experimental Analysis of Behavior, 1974, 22, 341-355. Brown, P. L. and Jenkins, H. M. Auto-shaping of the pigeon's keypeck. Journal of the Experimental Anal- ysis of Behavior, 1968, 11, 1-8. Craig, W.^ Observation^ on^ doves^ learning^ to^ drink. Journal of Animal Behavior, 1912, 3, 273-279. Gamzu, E. and Schwartz, B.^ The maintenance of^ key pecking by stimulus-contingent and^ response-inde- pendent food presentation. Journal of the^ Experi- mental Analysis of Behavior, 1973, 19,^ 65-72. Gamzu, E. and Williams, D. R. Classical conditioning of a complex skeletal response. Science, 1971, 171, 923-925. Gamzu, E. and^ Williams,^ D.^ R.^ Associative^ factors underlying the pigeon's key^ pecking^ in^ autoshaping procedures. Journal of the Experimental Analysis^ of Behavior, 1973, 19, 225-232. Griffin, R. W. and Rashotte, M. E. A^ note^ on^ the^ neg- ative automaintenance procedure. Bulletin of the Psychonomic Society, 1973, 2, 402-404. Hearst, E. and Jenkins, H. M. Sign tracking: the stimulus-reinforcer relation and directed action. Psychonomic Society Monograph, 1974. Hilgard, E.^ R.^ The^ nature^ of^ the^ conditioned^ re- sponse: I.^ The^ case^ for and^ against^ stimulus^ substi- tution. Psychological Review, 1936,^ 43, 366-385. Jenkins, H. M. and Moore, B.^ R.^ The form of^ the auto-shaped response with^ food^ or^ water^ rein- forcer. Journal of the Experimental Analysis of Be- havior, 1973, 20, 163-181. Lorenz, K. Uber den Begriff der Instinkthandlung. Folia Biotheoretica, 1937, 2, 18-50.
Martin, I. and Levey, A. B. The genesis of the classi- cal conditioned response, Oxford: Pergamon Press,
Moore, l. R. On directed respondents. Unpublished doctoral dissertation, Stanford University, 1971. Moore, B. R. The role of directed Pavlovian reactions in simple instruumental learning in the pigeon. In R. A. Hinde and J. Stevenson-Hinde (Eds.), Con- straints on learning. New York: Academic Press,
- Pp. 159-186. Perkins, C. C. Jr., Beaver, W. O., Hancock, R. A. Jr., Hemmendinger, P. C., Hemmendinger, D., and Ricci, J. Some variables affecting rate of key peck- ing durinig response-independent procedures (auto- shaping). Journal of the Experimental Analysis of Behavior, 1975, 24, 59-72. Rescorla, R. A. and Solomon, R. L. Two-process learning theory: relationships between^ Pavlovian conditioning and instrumental^ learning. Psychologi- cal Review, 1967, 74, 151-182. Schwartz, B. The role of positive conditioned rein- forcement in the maintenance of key-pecking which prevents delivery of a primary reinforcement. Psy- chonomic Science, 1972, 28, 277-278. Schwartz, B. and Gamzu, E. Pavlovian control of oper- ant behavior: an (^) analysis of (^) autoshaping and its implications for (^) operant conditioning. In (^) W. K. Honig and J. E. R. Staddon (Eds.), Handbook^ of operant behavior. New York:^ Prentice Hall, (in press). Staddon, J. E. R. and Simmelhag, V. L. The "super- stition" experiment: a re-examination of its implica- tions for the principles of adaptive behavior. Psy- chological Review, 1971, 78, 3-43. Wasserman, E. A. Pavlovian conditioning with heat reinforcement produces stimulus-directed pecking in chicks. Science, 1973, 181, 1875-1877. Williams, D. R. and Williams, H. Automaintenance in the pigeon: Sustained pecking despite contingent nonreinforcement. Journal of the Experimental Analysis of Behavior, 1969, 12, 511-520. Zener, K. The significance of behavior accompanying conditioned salivary secretion^ for^ theories of^ the conditioned response. American Journal of Psy- chology, 1937, 50, 384-403.
Received 3 April 1975. (Final Acceptance 11 March 1976.)