¡Descarga El Citoplasma: Función, Componentes y Procesos de Plegamiento de Proteínas - Prof. Ferrer y más Apuntes en PDF de Biología Celular solo en Docsity!
4.2. Citoplasma
- El citoplasma és el territori entre el nucli i la membrana. Inclou als orgànuls cel·lulars i al citoesquelet. A més conté partícules i inclusions.
- La fracció soluble del citoplasma és el citosol o hialoplasma.
- En el citosol hi ha aigua, ions, nutrients, en concentracions molt precises. I proteïnes (estructurals, reguladores etc).
- S’hi realitzen reaccions metabòliques diverses (glicòlisi, síntesi de sucres, aminoàcids, ac. grassos).
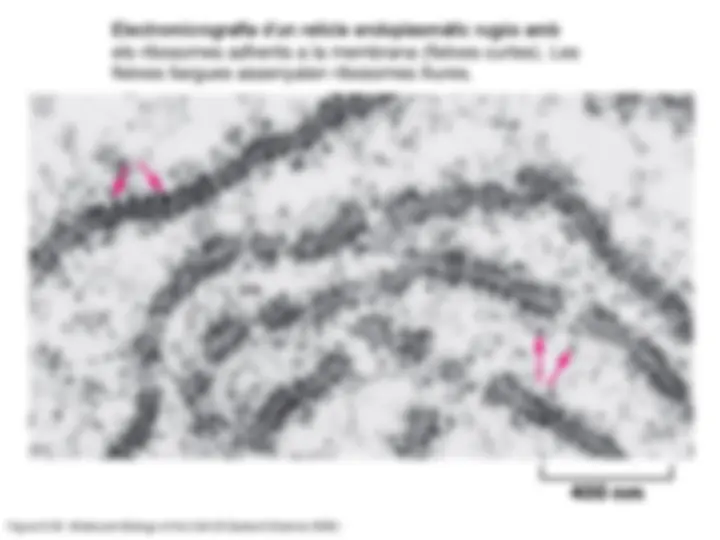
- Les partícules més abundants del citoplasma són els ribosomes , encarregats de la síntesi de les proteïnes.
- La majoria de proteïnes sintetitzades al citosol hi realitzen tota la seva funció en aquest compartiment. Hom diu que són proteïnes residents en el citosol. Hi són exemples les proteïnes del citosquelet o els enzims que catalitzen reaccions que tenen lloc en el citosol. Per assolir llur funcionalitat, aquestes proteïnes han de ser plegades. Aquest plegament el realitzen diferents proteïnes de plegament o xaperones residents també en el citosol.
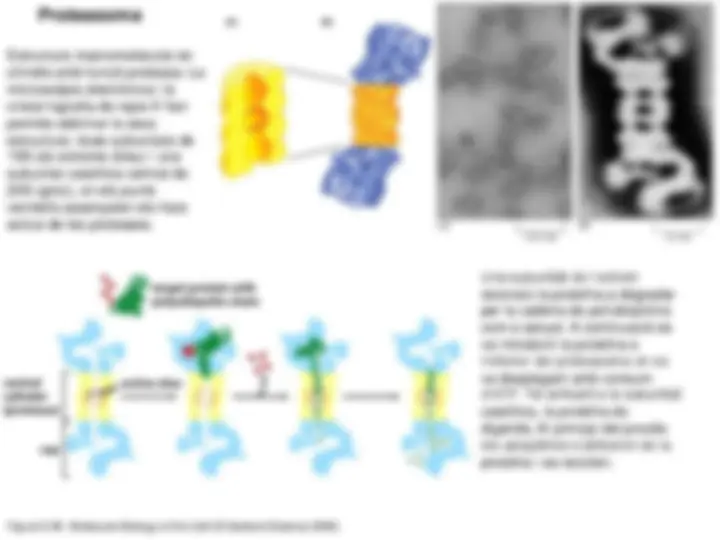
- En el citosol també té lloc una degradació. Concretament les proteïnes velles o defectuoses es degraden en unes partícules citoplasmàtiques anomenades proteasomes.
- En algunes cèl·lules animals, com ara els hepatòcits o les fibres musculars, el citoplasma conte inclusions de glicogen, com a magatzem de carbohidrats.
- En el citoplasma dels adipòcits, el citoplasma inclou gotes de lípids.
Proteòmica subcel·lular
Table 6-4 Molecular Biology of the Cell (© Garland Science 2008)
Figure 6-82 Molecular Biology of the Cell (© Garland Science 2008)
Plegament: Per la
confecció d’una proteïna
funcional, el polipèptid
naixent del ribosoma s’ha
de plegar correctament en
la seva conformació
tridimensional. Això
s’assoleix per medi
d’interaccions no covalents
entre els diferents dominis
de la molècula i per medi de
cofactors de petita massa
molecular. Algunes
proteïnes necessiten, a
més, modificacions
postraductives de tipus
covalent. Moltes vegades
això té lloc a mesura que el
polipèptid abandona el
ribosoma.
Cicle de la Hsp 70
Hsp 70 és una família molt gran de xaperones que actuen en el plegament no solament de proteïnes naixents, si no de proteïnes sintetitzades complertament. En el cicle de la xaperona, aquesta reconeix a la proteïna incomplertament plegada i d’una forma ràpida s’hi uneix per un domini d’unió, SBD, de la xaperona amb la conformació d’unió a ATP (1). La hidròlisi del nucleòtid per una proteïna satèlit, la DnaJ/Hsp40, promou un canvi conformacional de la xaperona en el que la proteïna a plegar queda entrapada dins del domini SBD, ara en forma de bossa. La xaperona esta ara amb la conformació d’unió a ADP (2). El bescanvi de nucleòtids, ADP per ATP, per un altra proteïna satèlit (GrpE/BAG1), promou un canvi conformacional de la xaperona en la que el domini SBD s’obre (3) alliberant-se la proteïna, plegada totalment o no (4). En aquest darrer cas es torna a repetir el cicle.
Lodish, 6ª Ed. 2008
Seminari. Xaperona Hsp
Les xaperones Hsp70 s’uneixen a regions hidròfobes dels polipèptids que s’estan plegant. Aquesta unió es va estudiar per la xaperona DnaK, una de les dues principals xaperones Hsp70 de E. coli. Amb aquesta finalitat, es marcaren bacteris salvatges amb metionina 35 S durant un temps pols de 15 segons, i a continuació s’aïllaren proteïnes en fase de plegament, en absència d ‘ATP, per immunoprecipitació amb anticossos contra la DnaK. Es separaren electroforèticament per SDS-PAGE i una autoradiografia posterior va permetre visualitzar les proteïnes de l’immunoprecipitat en la figura (A) carril 1. DnaK te un pes molecular de 66 kD. El carril 2 correspon a un immunoprecipitat tot tractant els bacteris amb un potent detergent, com ara l’SDS. El carril 3 correspon a un immunoprecipitat procedent de bacteris d’E coli mutants que no expressen DnaK. I el carril 4 correspon a un immunoprecipitat de bacteris salvatges però que no s’ha fet cap marcatge radioactiu (temps pols = 0). La figura B correspon a un cultiu de bacteris, però suplementant-lo amb un excés de metionina no marcada, després del temps pols, i fent la mateixa immunoprecipitació en diferents temps cacera i en presència d’ATP.
1. Quin tipus d’experiments constitueixen els immunoprecipitats corresponents als carrils 2, 3 i 4 **de la figura A?
- Quins foren els resultats amb presència d’ATP?** **Doneu-hi una explicació
- Per què les proteïnes marcades desapareixen** **d’acord amb els temps cacera?
- Demostren aquests experiments que la DnaK** s’uneix a les proteïnes mentre aquestes encara estan sent sintetitzades en els ribosomes?
Ubicuitines i ubicuitinització
Molècula d’ubicuitina: 76
aminoàcids
Les proteïnes a degradar pels proteasomes han d’estar poliubiqüitinitzades. Per la primera molècula d’ubiqüitina, un enzim activador de la ubiqüitina, E1, forma un enllaç tioéster entre una cisteïna de E1 i una molècula d’ubiqüitina. Aquest enllaç és d’alta energia, consumint-se ATP en el procés (1). A continuació la molècula d’ubiqüitina es transfereix a un enzim conjugador de la ubiqüitina, E2, formant-se igualment un enllaç tioéster amb una cisteïna d’E2 (2). Per últim es transfereix la ubiqüitina a un tercer enzim, la ubiqüitina-lligasa E3, la qual a la vegada catalitza la formació d’un enllaç isopeptídic entre l’extrem carboxil de la ubiqüitina i una lisina del substrat diana (3). La segona molècula d’ubiqüitina forma un enllaç isopeptídic entre el seu extrem carboxil i la lisina 48 de la primera ubiqüitina, igualment catalitzat per la ubiqüitina-lligasa E3, lo que suposa que la segona ubiqüitina ha estat transferida segons els tres passos 1, 2 i 3. Per tant els tres passos es repeteixen tantes vegades com a molècules d’ubiqüitina s’hi afegeixen. La cadena de poliubiqüitina es reconeguda pel proteasoma, el qual, amb consum d’ATP, desplega la proteïna (4) i la digereix, resultant de tot plegat les molècules d’ubicqüitina i els oligopèptids (5).
GOLBERG, A.L. et al. 2001. Proteasomas. Investigación y Ciencia, 294 (Marzo):22-
Figure 6-93 Molecular Biology of the Cell (© Garland Science 2008)
Diferents graus d’ubicuitinització