¡Descarga 4 apoptosis y más Guías, Proyectos, Investigaciones en PDF de Bioquímica solo en Docsity!
O RI G I N A L P A P E R
Does melatonin influence the apoptosis in rat uterus of animals
exposed to continuous light?
Cecı´lia S. Ferreira 1,2^ •^ Ka´tia C. Carvalho 2 •^ Carla C. Maganhin 2 •^ Ana P. R. Paiotti 3 • Celina T. F. Oshima 3 •^ Manuel J. Simo˜es 4 •^ Edmund C. Baracat 2 • Jose´ M. Soares Jr. 2
Published online: 5 November 2015 Ó Springer Science+Business Media New York 2015
Abstract Melatonin has been described as a protective agent against cell death and oxidative stress in different tissues, including in the reproductive system. However, the information on the action of this hormone in rat uterine apoptosis is low. Our objective was to evaluate the effects of melatonin on mechanisms of cell death in uterus of rats exposed to continuous light stress. Twenty adult Wistar rats were divided into two groups: GContr (vehicle control) and GExp which were treated with melatonin (0.4 mg/mL), both were exposed to continuous light for 90 days. The uterus was removed and processed for quantitative real time PCR (qRT-PCR), using PCR-array plates of the apoptosis pathway; for immunohistochemistry and TUNEL. The results of qRT-PCR of GEXP group showed up-regulation of 13 and 7, pro-apoptotic and anti-apoptotic genes, respectively, compared to GContr group. No dif- ference in pro-apoptotic proteins (Bax, Fas and Faslg) expression was observed by immunohistochemistry, although the number of TUNEL-positive cells was lower in the group treated with melatonin compared to the group not
treated with this hormone. Our data suggest that melatonin influences the mechanism and decreases the apoptosis in uterus of rats exposed to continuous light.
Keywords Melatonin � Female rats � Continuous light � Apoptosis
Introduction
Melatonin is an indoleamine synthesized and secreted in a rhythm pattern by the pineal gland of vertebrates, it has high serum levels during the nocturnal phase of the circa- dian cycle [1–4]. This neurohormone is an important endogenous mediator of photoperiodic information and is involved in the control of circadian and seasonal processes in many species [5, 6]. In fact, melatonin is a highly pleiotropic molecule, present in many tissues and systems (nervous, cardiovascular, gastrointestinal, reproductive, etc.). It produces anti-inflammatory, antioxidant and immunomodulating activities and further, modulates pro- liferation, differentiation and apoptosis in many tissues with the interaction of hormones and growth factors [7– 11]. In the reproductive system the presence of specific receptors for melatonin in ovaries, mammary glands, testes and uterus was demonstrated [12–15]. In rodents, mela- tonin could be considered to directly influence ovarian function; increasing the secretion of estrogen and proges- terone by the granulosa cells and disrupt the progression of the estrous cycle [16, 17]. Research also demonstrated that melatonin associated with progesterone might also inhibit ovarian aromatase expression and increase uterine recep- tors in female mice [18]. Other studies show that pinealectomy and continuous light exposure (a method of
& Cecı´lia S. Ferreira [email protected]
(^1) Departamento de Ginecologia, Universidade Federal de Sa˜o
Paulo, Avenida Doutor Arnaldo, 455. Sala 2113. Cerqueira Ce´sar, CEP: 01246-923 Sa˜o Paulo, Brazil (^2) Laboratory of Structural and Molecular Gynecology (LIM-
58), Disciplina de Ginecologia - Departmento de Ginecologia e Obstetrı´cia, Faculdade de Medicina da Universidade de Sa˜o Paulo, Sa˜o Paulo, Brazil (^3) Laboratory of Pathology Molecular, Departamento de
Patologia, Universidade Federal de Sa˜o Paulo, Sa˜o Paulo, Brazil (^4) Departamento de Morfologia e Gene´tica, Universidade
Federal de Sa˜o Paulo, Sa˜o Paulo, Brazil
DOI 10.1007/s10495-015-1195-
decreasing the endogenous melatonin) [19] on rodents can decrease the levels of gonadotropins (LH and FSH), induce precocious puberty, ovarian atrophy, hyperprolactinemia and inhibition of uterine implantation [20–23], this indi- cates that the hormone has an important role to play in the maintenance of the homeostasis in the female reproductive tract. Studies have shown the action of melatonin in the cell death of different tissues and systems, including the reproductive system. It seems that this endoleamine pre- sents a modulating role in the apoptosis process and it can present as both pro-apoptotic and anti-apoptotic actions [24, 25]. Anti-apoptotic action of melatonin was demon- strated on the thymus, kidney, brain and liver and this action is attributed by some authors, to the antioxidant properties of indoleamine [26] and by the elimination of hydroxyl anion (–OH), peroxyl (ROO–) and superoxide [27–32]. The modulation of immunological elements is also another mechanism by which melatonin may inhibit apoptosis by inducing the release of cytokines, such as interleukin-4 [33]. Melatonin may also interact with nuclear receptors and change the gene expression of inducers or inhibitors of apoptosis [34, 35]. According to a well-established model of decreasing the endogenous melatonin (exposition to continuous light) and the action of this hormone on cell death in different tissues, our study aimed to evaluate the effects of melatonin deprivation on the uterus. It is known that this endoleamine causes a decrease in the levels of estradiol, an important protector of cell death, although, some studies also show that melatonin has a protective action on mammalian ovaries [36, 37].
Materials and methods
Animals
Twenty virgin adult female Wistar rats (Rattus norvegicus albinus), with regular estrous cycle, weighing between 200 and 300 g, from the Development of Experimental Model in Medicine and Biology Center (CEDEME) of the Federal University of Sa˜o Paulo (UNIFESP) were used in the research. The animals were housed in plastic cages with food and water ad libitum, under the controlled tempera- ture of (22 ± 2 °C) and randomly divided into two groups: GContr and GExp, both groups had ten animals and were kept under constant artificial light (400 lux) for 90 days [19, 21]. The Laboratory Animal Care Committee of Federal University of Sa˜o Paulo approved this study (CEP 103/09) and all the animals used were handled in accor- dance with the Guiding Principles for The Care and Use of Animals.
Experimental groups
GExp group animals were treated with melatonin (Sigma Chemical Company, St. Louis, MO, USA) dissolved in 1 ml ethanol and diluted in the drinking water (0.4 mg/mL) for 90 consecutive days from 6:00 p.m. to 8:00 a.m. While GContr group animals received only water with the same concentration of ethanol (vehicle) for the same period of time [21]. After 90 days under treatment, vaginal smears were collected from the animals, those animals who were in the proestrous phase, from the GExp group, were euth- anized by decapitation, their blood was collected and their uterus’ were removed. The remaining animals, from the same group, continued under the same treatment and were euthanized once they reached the stage of the estrous cycle. All GContr group animals were euthanized after 90 days under treatment, as they were in persistent estrous. The uterus’ of the animals were removed and properly preserved according to the method which it had been subjected to. Right horn was conserved in liquid nitrogen and then frozen at - 80 °C, for gene expression analysis, and left horn fixed in 10 % buffered formaldehyde, for immunohistochemical, TUNEL techniques and hematox- ilin–eosin staining.
Melatonin measurement
The serum was obtained from the blood by centrifugation and melatonin was measured by ELISA method using IBL Kit (IBL International GmbH, Hamburg, Germany). All samples were measured in triplicate and the concentration of melatonin was given in pg/mL.
Extraction and purification of total RNA
Total RNA was extracted using QIAZOL Lysis Reagent (QIAGEN, Venlo, Netherlands) according to manufac- turer’s instructions and then treated with RNase-Free DNase Set (QIAGEN, USA). The total RNA obtained from each sample was quantified by spectrophotometric (ND NanoDrop Ò—Thermo Fisher Scientific Inc. Co.) and its integrity profile assessed by electrophoresis on 1 % agarose gel.
Synthesis of complementary strand DNA (cDNA) and quantitative real time PCR (qPCR)
One microgram of total RNA purified from each sample was transcribed into cDNA by reverse transcription reac- tion, using the First Strand kit RT2 (QIAGEN), according to manufacturer’s instructions. The cDNA was subjected to reactions in qPCR, using PCR-array plates which present a large panel of the apoptosis pathway gene expression
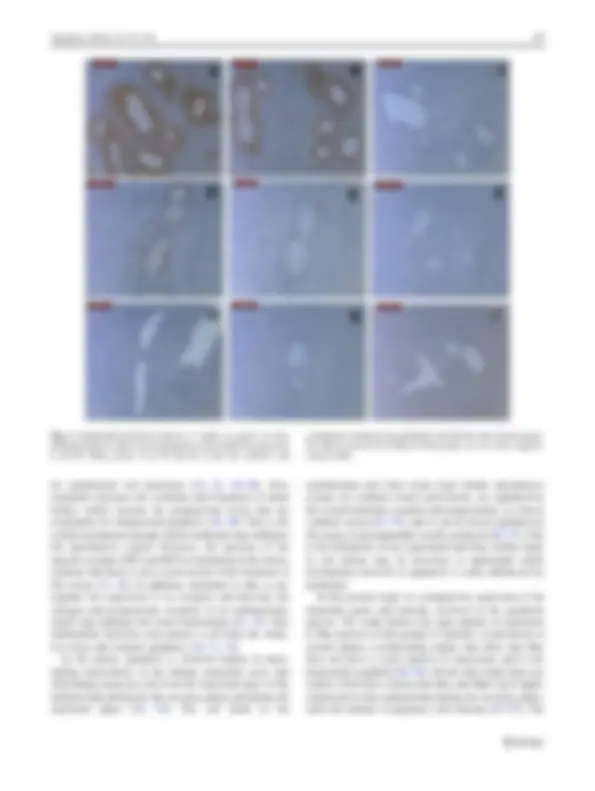
Immunohistochemical analysis
In contrast, immunohistochemical tests revealed no sig- nificant statistical difference in the expression of pro- apoptotic proteins studied (Table 2). Both groups showed positive and strong cytoplasmic labeling for Bax in the endometrial epithelium, glandular and stromal cells. Fas was present in the epithelial and glandular cells, with weak
labeling cytoplasmic. No labeling for Faslg was observed in all the animals (Fig. 2).
TUNEL
The morphological analyses showed that in both groups, the epithelium of endometrium presented as a simple cylindrical, with an elliptical core, located at basal pole, with one or more nucleoli. A significant statistical difference in the number of TUNEL-positive cells between the two groups (Table 2) was observed, with GEXp group presenting fewer cells with fragmented DNA, indicating with other findings that, in female rats submitted to continuous light, melatonin influences the pattern of cell death (Fig. 3).
Discussion
Studies have shown that melatonin clearly influence the reproductive function in both seasonal breeding and non- seasonal breeding animals. Female rats pinealectomized, or exposed to continuous light, for example, usually develop persistent estrous, a similar condition to the polycystic ovary syndrome, where there is high blood levels of estradiol and testosterone and low blood levels of proges- terone [21, 22]. These hormonal changes may produce proliferation on the endometrial tissue which could increase the risk of hyperplasia and neoplasia [44, 45]. In addition, melatonin treatment has also shown that it reverses this condition in rat pinealectomized or exposed to continuous light [21]. One possible explanation for this effect may be due to the apoptosis of uterine cells. In fact, our results corroborate this idea because melatonin decreases the cell death on the uterus’ of rats. It was demonstrated that in rodents and humans, mela- tonin has an antigonadotrophic effect, decreasing the secretion of LH and FSH and influencing the testosterone synthesis and testicular maturation in males. In females, melatonin influences ovarian function through a decrease in the levels of estradiol, which is an important hormone
Fig. 1 Serum melatonin levels of GContr (17.1 ± 8.43 pg/mL) and GExp (172.6 pg/mL ± 93.3), *p = 0.
Table 1 Pro-apoptotic genes of intrinsic and extrinsic pathway and anti-apoptotic genes up-regulated on GExp group
Gene Fold
Pro-apoptotic genes Intrinsic pathway Apaf-1 7. Bad 3. Bak1 6. Bcl10 3. Casp3 2. Casp6 4. Casp9 3. Tp63 4. Casp12 2. Extrinsic pathway CD40 2. Faslg 4. Anti-apoptotic genes Api5 7. Aven 8. Bag1 4. Bcl2 2. Bcl2a1d 6. Birc5 3. Faim 2.
Values expressed as relative fold change of expression compared to GContr group
Table 2 Data of Bax, Fas and Faslg of immunoreactivity and TUNEL positive-cells per field GContr GExp p value Mean ± SD Mean ± SD
Bax 4.857 ± 1.464 4.857 ± 2.295 0. Fas 3.250 ± 1.389 3.375 ± 1.923 1. Faslg 1.000 ± 0.926 0.889 ± 1.167 0. TUNEL 5.314 ± 2.863 3.129 ± 0.830 0.046* The asterisk indicates the value with statistical significance (p \ 0.05)
for endometrial cell protection [22, 23, 46–48]. Also, melatonin increases the ovulation and formation of luteal bodies, which increase the progesterone levels that are responsible for endometrial apoptosis [49, 50]. This is the central mechanism through which melatonin may influence the reproductive system. However, the presence of the specific receptors MT1 and MT2 of melatonin in the uterus, confirms that there is also a local action of this hormone in this tissue [12, 16]. In addition, melatonin is able to up- regulate the expression of its receptors and decrease the estrogen and progesterone receptors in rat endometrium, which may influence the tissue homeostasis [51, 52]. This indoleamine therefore, may protect a cell from the oxida- tive stress and controls apoptosis [10, 31, 35]. In the uterus, apoptosis is involved mainly in main- taining homeostasis in the human menstrual cycle and eliminating senescent cells from the functional layer of the endometrium during the late secretory phase and during the menstrual phase [53, 54]. The cell death in the
endometrium and other tissue from female reproductive system, for example ovaries and breasts, are regulated by the sexual hormones estradiol and progesterone, as well as cytokine action [54, 55], and it can be down-regulated by the action of prostaglandins locally produced [56, 57]. This is the limitations of our experiment and thus, further study in cell culture may be necessary to understand which mechanisms involved in apoptosis is really influenced by melatonin. In this present study we evaluated the expression of the important genes and proteins involved in the apoptosis process. We could observe the same pattern of expression of Bax protein in both groups of animals, in proestrous or estrous phases, corroborating studies that show that Bax does not have a cyclic pattern of expression, and is not hormonally regulated [58, 59]. On the other hand, there are studies which have shown that Bax and Bak reach higher expressions in the endometrium during the secretory phase, when the number of apoptotic cells increase [53–57]. The
Fig. 2 Immunohistochemical analysis of adults rat uterus for Bax labeling (a, b), Fas (d, e) and Faslg (g, h). a, d and g GContr group. b, e and h: GExp group. In a, b and d, e note the positive and
cytoplasmic staining in the glandular endometrial cells in both groups. No label is present for Falslg in both groups. In c, f, and i, negative control slides
Compliance with ethical standards
Conflict of interest The authors declare no conflict of interest.
References
- Reiter RJ (1993) The melatonin rhythm: both a clock and a calendar. Experientia 49:654–
- Ganguly S, Coon SL, Klein DC (2002) Control of melatonin synthesis in the mammalian pineal gland: the critical role of serotonin acetylation. Cell Tissue Res 309:127–
- Macchi MM, Bruce JN (2004) Human pineal physiology and functional significance of melatonin. Front Neuroendocrinol 25:177–
- Zawilska JB, Skene DJ, Arendt J (2009) Physiology and phar- macology of melatonin in relation to biological rhythms. Phar- macol Rep 61:383–
- Arendt J, Skene DJ (2005) Melatonin as a chronobiotic. Sleep Med Rev 9:25–
- Revel FG, Masson-Pevet M, Pevet P, Mikkelsen JD, Simonneaux V (2009) Melatonin controls seasonal breeding by a network of hypothalamic targets. Neuroendocrinology 90:1–
- Carrillo-Vico A, Lardone PJ, Naji L et al (2005) Beneficial pleiotropic actions of melatonin in an experimental model of septic shock in mice: regulation of pro-/anti-inflammatory cyto- kine network, protection against oxidative damage and anti- apoptotic effects. J Pineal Res 39:400–
- Claustrat B, Brun J, Chazot G (2005) The basic physiology and pathophysiology of melatonin. Sleep Med Rev 9:11–
- Hardeland R, Pandi-Perumal SR, Cardinali DP (2006) Melatonin. Int J Biochem Cell Biol 38:313–
- Radogna F, Paternoster L, Albertini MC et al (2006) Melatonin as an apoptosis antagonist. Ann NY Acad Sci 1090:226–
- Sarlak G, Jenwitheesuk A, Chetsawang B, Govitrapong P (2013) Effects of melatonin on nervous system aging: neurogenesis and neurodegeneration. J Pharmacol Sci 123:9–
- Vanecek J (1998) Cellular mechanisms of melatonin action. Physiol Rev 78:687–
- Clemens JW, Jarzynka MJ, Witt-Enderby PA (2001) Down-reg- ulation of mt1 melatonin receptors in rat ovary following estro- gen exposure. Life Sci 69:27–
- Ekmekcioglu C (2006) Melatonin receptors in humans: biological role and clinical relevance. Biomed Pharmacother 60:97–
- Tamura H, Takasaki A, Taketani T et al (2014) Melatonin and female reproduction. J Obstet Gynaecol Res 40:1–
- Soares JM Jr, Masana MI, Ersahin C, Dubocovich ML (2003) Functional melatonin receptors in rat ovaries at various stages of the estrous cycle. J Pharmacol Exp Ther 306:694–
- Itoh MT, Hosaka T, Takahashi N, Ishizuka B (2006) Expression of luteinizing hormone/chorionic gonadotropin receptor in the rat pineal gland. J Pineal Res 41:35–
- Bondi CD, Alonso-Gonzalez C, Clafshenkel WP et al (2014) The effect of estradiol, progesterone, and melatonin on estrous cycling and ovarian aromatase expression in intact female mice. Eur J Obstet Gynecol Reprod Biol 174:80–
- Wideman CH, Murphy HM (2009) Constant light induces alter- ations in melatonin levels, food intake, feed efficiency, visceral adiposity, and circadian rhythms in rats. Nutr Neurosci 12:233–
- Acuna-Castroviejo D, Fernandez B, Castillo JL, del Aguila CM (1993) Similarity between the effects of suprachiasmatic nuclei lesions and of pinealectomy on gonadotropin release in ovariec- tomized, sulpiride-treated and melatonin-replaced rats. Experi- entia 49:797–
- Prata Lima MF, Baracat EC, Simoes MJ (2004) Effects of melatonin on the ovarian response to pinealectomy or continuous light in female rats: similarity with polycystic ovary syndrome. Braz J Med Biol Res 37:987–
- Dair EL, Simoes RS, Simoes MJ et al (2008) Effects of melatonin on the endometrial morphology and embryo implantation in rats. Fertil Steril 89:1299–
- Dardes RC, Baracat EC, Simo˜es MJ (2000) Modulation of estrous cycle and LH, FSH and melatonin levels by pinealectomy and sham-pinealectomy in female rats. Prog Neuropsychopharmacol Biol Psychiatry 24:441–
- Rodriguez C, Martin V, Herrera F et al (2013) Mechanisms involved in the pro-apoptotic effect of melatonin in cancer cells. Int J Mol Sci 14:6597–
- Bizzarri M, Proietti S, Cucina A, Reiter RJ (2013) Molecular mechanisms of the pro-apoptotic actions of melatonin in cancer: a review. Expert Opin Ther Targets 17:1483–
- Sainz RM, Mayo JC, Rodriguez C, Tan DX, Lopez-Burillo S, Reiter RJ (2003) Melatonin and cell death: differential actions on apoptosis in normal and cancer cells. Cell Mol Life Sci 60:1407–
- Pieri C, Marra M, Moroni F, Recchioni R, Marcheselli F (1994) Melatonin: a peroxyl radical scavenger more effective than vitamin E. Life Sci 55:Pl271–Pl
- Barlow-Walden LR, Reiter RJ, Abe M et al (1995) Melatonin stimulates brain glutathione peroxidase activity. Neurochem Int 26:497–
- Nava M, Romero F, Quiroz Y, Parra G, Bonet L, Rodriguez- Iturbe B (2000) Melatonin attenuates acute renal failure and oxidative stress induced by mercuric chloride in rats. Am J Physiol Renal Physiol 279:F910–
- Hoijman E, Rocha Viegas L, Keller Sarmiento MI, Rosenstein RE, Pecci A (2004) Involvement of Bax protein in the prevention of glucocorticoid-induced thymocytes apoptosis by melatonin. Endocrinology 145:418–
- Baydas G, Koz ST, Tuzcu M, Etem E, Nedzvetsky VS (2007) Melatonin inhibits oxidative stress and apoptosis in fetal brains of hyperhomocysteinemic rat dams. J Pineal Res 43:225–
- Molpeceres V, Mauriz JL, Garcia-Mediavilla MV, Gonzalez P, Barrio JP, Gonzalez-Gallego J (2007) Melatonin is able to reduce the apoptotic liver changes induced by aging via inhibition of the intrinsic pathway of apoptosis. J Gerontol A Biol Sci Med Sci 62:687–
- Maestroni GJ (1993) The immunoneuroendocrine role of mela- tonin. J Pineal Res 14:1–
- Mediavilla MD, Cos S, Sanchez-Barcelo EJ (1999) Melatonin increases p53 and p21WAF1 expression in MCF-7 human breast cancer cells in vitro. Life Sci 65:415–
- Pedreanez A, Rincon J, Romero M, Viera N, Mosquera J (2004) Melatonin decreases apoptosis and expression of apoptosis-as- sociated proteins in acute puromycin aminonucleoside nephrosis. Nephrol Dial Transpl 19:1098–
- Voznesenskaya T, Makogon N, Bryzgina T, Sukhina V, Grushka N, Alexeyeva I (2007) Melatonin protects against experimental immune ovarian failure in mice. Reprod Biol 7:207–
- Wang SJ, Liu WJ, Wu CJ et al (2012) Melatonin suppresses apoptosis and stimulates progesterone production by bovine granulosa cells via its receptors (MT1 and MT2). Theriogenology 78:1517–
- Barrezueta LF, Oshima CT, Lima FO et al (2010) The intrinsic apoptotic signaling pathway in gastric adenocarcinomas of Brazilian patients: immunoexpression of the Bcl-2 family (Bcl-2, Bcl-x, Bak, Bax, Bad) determined by tissue microarray analysis. Mol Med Rep 3:261–
- Paiotti AP, Ribeiro DA, Silva RM et al (2012) Effect of COX- inhibitor lumiracoxib and the TNF-alpha antagonist etanercept on TNBS-induced colitis in Wistar rats. J Mol Histol 43:307–
- Gavrieli Y, Sherman Y, Ben-Sasson SA (1992) Identification of programmed cell death in situ via specific labeling of nuclear DNA fragmentation. J Cell Biol 119:493–
- Beigi Boroujeni M, Salehnia M, Khalatbary AR, Pourbeiranvand S, Beigi Boroujeni N, Ebrahimi S (2010) Effect of ovarian stimulation on the endometrial apoptosis at implantation period. Iran Biomed J 14:171–
- Bubenik GA, Brown GM (1997) Pinealectomy reduces melatonin levels in the serum but not in the gastrointestinal tract of rats. Biol Signals 6:40–
- Maganhin CC, Simoes RS, Fuchs LF et al (2014) Melatonin influences on steroidogenic gene expression in the ovary of pinealectomized rats. Fertil Steril 102:291–
- Siiteri PK (1978) Steroid hormones and endometrial cancer. Cancer Res 38:4360–
- Konishi I, Koshiyama M, Mandai M et al (1997) Increased expression of LH/hCG receptors in endometrial hyperplasia and carcinoma in anovulatory women. Gynecol Oncol 65:273–
- Grosse J, Maywood ES, Ebling FJ, Hastings MH (1993) Testic- ular regression in pinealectomized Syrian hamsters following infusions of melatonin delivered on non-circadian schedules. Biol Reprod 49:666–
- Li C, Zhou X (2015) Melatonin and male reproduction. Clin Chim Acta 446:175–
- Aleandri V, Spina V, Morini A (1996) The pineal gland and reproduction. Hum Reprod Update 2:225–
- Romeu LR, da Motta EL, Maganhin CC et al (2011) Effects of melatonin on histomorphology and on the expression of steroid receptors, VEGF, and PCNA in ovaries of pinealectomized female rats. Fertil Steril 95:1379–
- Tamura H, Nakamura Y, Korkmaz A et al (2009) Melatonin and the ovary: physiological and pathophysiological implications. Fertil Steril 92:328–
- Chuffa LG, Seiva FR, Favaro WJ et al (2011) Melatonin reduces LH, 17 beta-estradiol and induces differential regulation of sex steroid receptors in reproductive tissues during rat ovulation. Reprod Biol Endocrinol 9:
- Slominski RM, Reiter RJ, Schlabritz-Loutsevitch N, Ostrom RS, Slominski AT (2012) Melatonin membrane receptors in periph- eral tissues: distribution and functions. Mol Cell Endocrinol 351:152–
- Kokawa K, Shikone T, Nakano R (1996) Apoptosis in the human uterine endometrium during the menstrual cycle. J Clin Endo- crinol Metab 81:4144–
- Harada T, Kaponis A, Iwabe T et al (2004) Apoptosis in human endometrium and endometriosis. Hum Reprod Update 10:29–
- Vaskivuo TE, Stenback F, Karhumaa P, Risteli J, Dunkel L, Tapanainen JS (2000) Apoptosis and apoptosis-related proteins in human endometrium. Mol Cell Endocrinol 165:75–
- Slayden OD, Hirst JJ, Brenner RM (1993) Estrogen action in the reproductive tract of rhesus monkeys during antiprogestin treat- ment. Endocrinology 132:1845–
- Gosden R, Spears N (1997) Programmed cell death in the reproductive system. Br Med Bull 53:644–
- Teixeira C, Reed JC, Pratt MA (1995) Estrogen promotes chemotherapeutic drug resistance by a mechanism involving Bcl- 2 proto-oncogene expression in human breast cancer cells. Can- cer Res 55:3902–
- McLaren J, Prentice A, Charnock-Jones DS, Sharkey AM, Smith SK (1997) Immunolocalization of the apoptosis regulating pro- teins Bcl-2 and Bax in human endometrium and isolated peri- toneal fluid macrophages in endometriosis. Hum Reprod 12:146–
- Gompel A, Sabourin JC, Martin A et al (1994) Bcl-2 expression in normal endometrium during the menstrual cycle. Am J Pathol 144:1195–
- Castro A, Johnson MC, Anido M, Cortinez A, Gabler F, Vega M (2002) Role of nitric oxide and bcl-2 family genes in the regu- lation of human endometrial apoptosis. Fertil Steril 78:587–
- Otsuki Y, Misaki O, Sugimoto O, Ito Y, Tsujimoto Y, Akao Y (1994) Cyclic bcl-2 gene expression in human uterine endome- trium during menstrual cycle. Lancet 344:28–
- Yamashita H, Otsuki Y, Matsumoto K, Ueki K, Ueki M (1999) Fas ligand, Fas antigen and Bcl-2 expression in human endo- metrium during the menstrual cycle. Mol Hum Reprod 5:358–
- Selam B, Kayisli UA, Mulayim N, Arici A (2001) Regulation of Fas ligand expression by estradiol and progesterone in human endometrium. Biol Reprod 65:979–
- Song J, Rutherford T, Naftolin F, Brown S, Mor G (2002) Hor- monal regulation of apoptosis and the Fas and Fas ligand system in human endometrial cells. Mol Hum Reprod 8:447–
- Panzan MQ, Mattar R, Maganhin CC et al (2013) Evaluation of FAS and caspase-3 in the endometrial tissue of patients with idiopathic infertility and recurrent pregnancy loss. Eur J Obstet Gynecol Reprod Biol 167:47–
- Tewari M, Yu M, Ross B, Dean C, Giordano A, Rubin R (1997) AAC-11, a novel cDNA that inhibits apoptosis after growth factor withdrawal. Cancer Res 57:4063–
- Chau BN, Cheng EH, Kerr DA, Hardwick JM (2000) Aven, a novel inhibitor of caspase activation, binds Bcl-xL and Apaf-1. Mol Cell 6:31–
- Kutuk O, Temel SG, Tolunay S, Basaga H (2010) Aven blocks DNA damage-induced apoptosis by stabilising Bcl-xL. Eur J Cancer 46:2494–
- Czabotar PE, Lessene G, Strasser A, Adams JM (2014) Control of apoptosis by the BCL-2 protein family: implications for physi- ology and therapy. Nat Rev Mol Cell Biol 15:49–