¡Descarga apuntes botanica y más Apuntes en PDF de Botánica y Agronomía solo en Docsity!
ros pteridófitos terrestres. No está clara la conexión de los
antocerotópsidos, que se desarrollan muy tempranamente
y de forma paralela a las hepáticas y los briópsidos. Aún
se discute si, entre los musgos, estos son los más próximos
a las plantas terrestres superiores verdes. En sus formas
primitivas, tanto musgos como plantas vasculares habían
tomado las estructuras conductoras de agua y asimilados
de sus antepasados comunes. Lo corroboran las notables
coincidencias en cuanto a estructura y función de los hi-
droides y leptoides de los musgos con las traqueidas (trá-
queas) y células cribosas de los pteridófitos y, finalmente,
la presencia de arquegonios y estomas en ambos grupos.
Los briófitos han seguido un desarrollo en el cual su ga-
metófito (por lo tanto el musgo verde), al igual probable-
mente que el esporófito, se han visto sometidos a diversas
simplificaciones y, al mismo tiempo, a progresiones (re-
ducción progresiva). Así, dentro de los briópsidos se han
considerado primitivas formas erectas, acrocárpicas y al-
tas, con un sistema conductor bien desarrollado y seme-
jante a una protostela (politricanas) frente a las formas ba-
jas, ramificadas y pleurocárpicas sin sistema conductor ni
nervio central. Asimismo, la presencia de estomas afun-
cionales apunta hacia una regresión. En cambio, han ex-
perimentado un desarrollo continuo la red celular de las
hojas (de parenquimática a prosenquimática), el sistema
de ramificación (de acrocárpico a pleurocárpico) y la for-
mación de la cápsula (perístoma).
Se han encontrado briófitos fósiles esporádicamente desde el Devó-
nico superior; no han aportado mucho al conocimiento de su origen
filogenético. Sporogonites puede ser un elemento de conexión dis-
cutible entre los briófitos y los pteridófitos relacionados (psilofitóp-
sidos ). Las hepáticas talosas y foliosas, al igual que los primeros
briópsidos (Muscites) aparecen en el Carbonífero inglés. Esto apo-
ya la gran antigüedad de los briófitos. Se han descrito musgos, atri-
buibles a los esfágnidos y bríidos, del Pérmico de Rusia (Pechora,
Kuznetsk). Los briópsidos del Carbonífero inferior y del Pérmico
poseían nervios centrales en las hojas (también las protosfagnales),
mientras que se conocen formas sin ellos sólo a partir del Triásico y
de finales del Jurásico. La mayoría de los musgos fósiles, incluso
con una proporción creciente de formas pleurocárpicas, proceden
del Terciario y se pueden clasificar en los géneros actuales.
Tercera subdivisión:
Pteridophytina, helechos
Los pteridófitos se han adaptado primariamente a la vida
terrestre, como los musgos; sin embargo, sus órganos y te-
jidos están mucho más diferenciados funcionalmente.
Cuadro 11-9: Traqueófitos
En el tallo, en las hojas y en las raíces de pteridófitos y esper-
matófitos existen verdaderos haces conductores (no sólo cor-
dones conductores; v. más atrás), que sirven para transportar
sustancias (agua con sustancias minerales, asimilados). Se di-
ferenciaron de los musgos o briófitos, que carecen de tales ha-
ces conductores, con el nombre de traqueófitos o plantas vas-
culares. En los musgos con conducción interna de agua ya hay
cordones conductores, que se asemejan al tipo de estructura
primitiva de los haces conductores, pero cada cordón conduc-
tor está menos diferenciado del tejido circundante, y los ele-
Subdivisión: Pteridophytina, helechos 1 717
En la alternancia de generaciones de los pteridófitos re-
cientes domina el esporófito (fig. 11-128 H). Constituye
una planta verde independiente y se divide en tallo, hojas
y raíces en los licodiópsidos, equisetópsidos y pterópsi-
dos. Los vástagos de los psilofitóspidos, ya extinguidos,
solían estar formados todavía por brotes o ramas bifurca-
das, afilas y semejantes (telomas, fig. 11-130 A, B). Care-
cían de verdaderas raíces, como les ocurre a los psilotóp-
sidos actuales.
El gametófito haploide en los pteridófitos se denomina
protalo (fig. 11-128 A); suele vivir unas pocas semanas
nada más y alcanza a lo sumo algunos centímetros de diá-
metro, siendo frecuentemente, por su aspecto, semejante
a una sencilla hepática talosa; en los casos típicos -las
excepciones a la regla son muy diversas-, se compone de
un talo verde, sencillo, fijo en el suelo por medio de ri-
zoides unicelulares y tubulosos que nacen en su cara in-
ferior. En dicho protalo se originan en gran número ante-
ridios y arquegonios, y la fecundación sólo es posible,
como en los briófitos, en el agua, es decir, con los prota-
los mojados.
Después de la fecundación, nace del zigoto la generación
diploide, el esporófito (fig. 11-128 H: 3, 4), que presen-
ta en los pteridófitos una configuración completamente
distinta y extraordinariamente más diferenciada que en
los briófitos. Sólo se parece al de estos en el principio de
su desarrollo. El protalo desaparece pronto en la mayo-
ría de las especies (si no tiene lugar la fecundación, pue-
de vivir años) y el embrión del esporófito da lugar a un
organismo independiente, de ordinario perenne, con raí-
ces, tallo y hojas: la planta del pteridófito en sentido es-
tricto (figs. 11-128 H: 4, 11-148 A, 11-150, 11-152 A).
Así pues, el esporófito de los pteridófitos es un verdade-
ro cormo.
Teoría telomática: los órganos típicos de los cormófitos
se han originado probablemente a partir de ramas bifurca-
das o dicotómicas afilas (telomas: fig. 11-130 A, B) a tra-
vés de unos procesos fundamentales (procesos elementa-
les de la teoría telomática), es decir, por culminación,
planación, concrescencia, reducción e incurvación (fig.
Por culminación (fig. 11-129 A) se establece a partir de
telomas equivalentes, que todavía componían el vástago
de la mayoría de los psilofitópsidos, una diferenciación y
una división del trabajo entre ejes principales portadores
y ejes laterales. El eje principal, culminante, mantiene un
impulso de crecimiento mayor que la rama hermana, so-
brepasada por él; dichas ramas se convierten entonces en
mentos conductores del agua no presentan lignina en sus pare-
des. Por lo tanto, los elementos conductores de agua de los
musgos carecen de paredes rígidas del tipo de las traqueidas y
tráqueas de las plantas vasculares. La disposición del haz con-
ductor en el tallo de los traqueófitos se denomina estela. Du-
rante el transcurso de su adaptación a la vida terrestre, la este-
la se fue estructurando más o menos de acuerdo con su función
para el transporte de sustancias y su rigidez (protostela, acti-
nostela, plectostela, polistela, sifonostela, eustela, atactostela,
teoría estélica, cuadro 4-3).
718 11 Sistemática y filogenia
wa
E
31·
4
o @
Ü 2
1
wa
e
B
as
D
unos órganos apendiculares situados lateralmente (B) que
pueden ir asumiendo la función de asimilar. En la plana-
ción, los ejes de los brotes laterales se orientan en un mis-
mo plano (C). Por concrescencia congénita, estos telo-
mas que estaban en un mismo plano se convierten en
órganos apendiculares planos y foliiformes (D, G). Así
pudieron haberse desarrollado las grandes hojas (macro y
megafilos) con frecuencia divididas y provistas además
de ramificación dicotómica. También pueden soldarse
entre sí telomas tridimensionales, con lo cual surge un
grueso eje parenquimático que ya no está atravesado por
un solo cordón conductor central (protostela, fig. 11-
C), sino que engloba dos o más haces conductores (fig.
11-129 H-K). Así se ve favorecida la resistencia del eje de
manera considerable. Por reducción (F, G) se puede ima-
ginar el origen de las hojas pequeñas más o menos acicu-
liformes y con un solo nervio (microfilos), aunque se dis-
cute sobre si, en vez de ser una excrescencia del eje, se
s r1t')
''fG
H
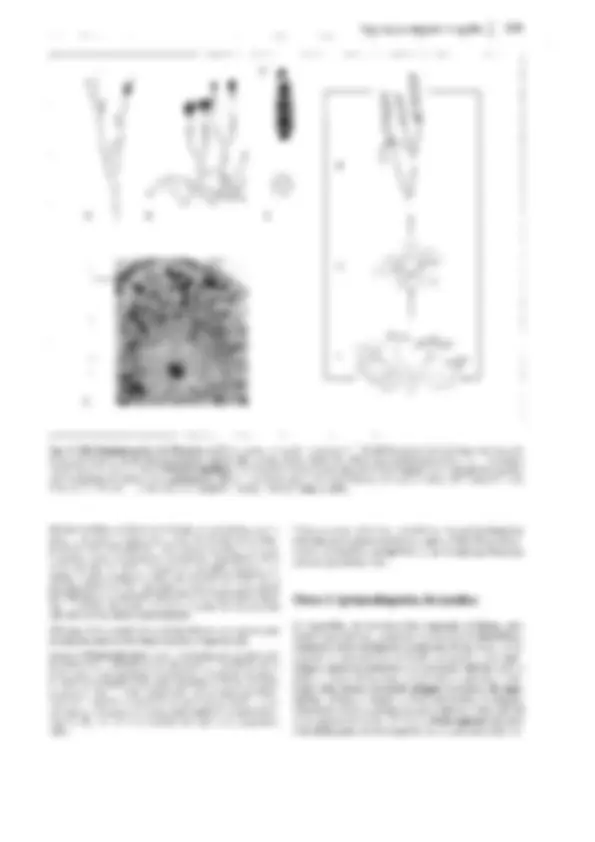
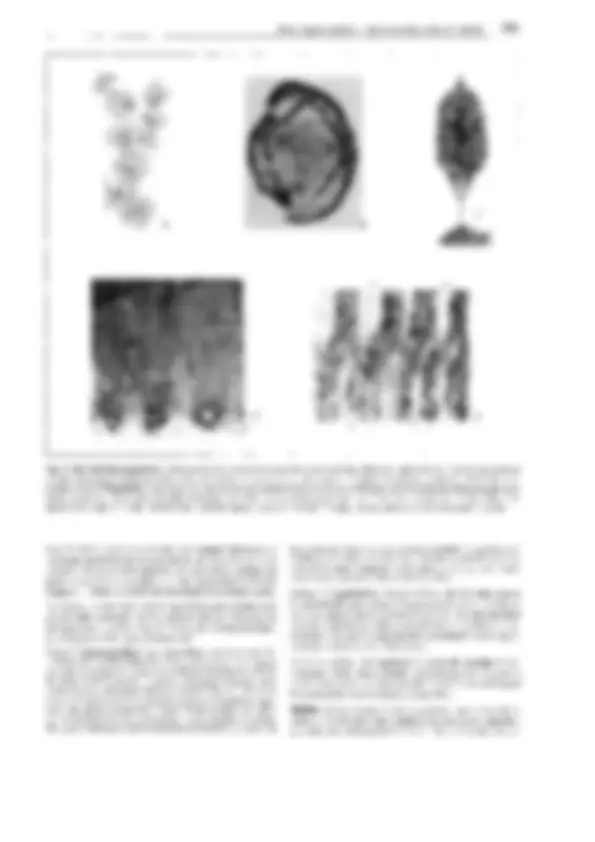
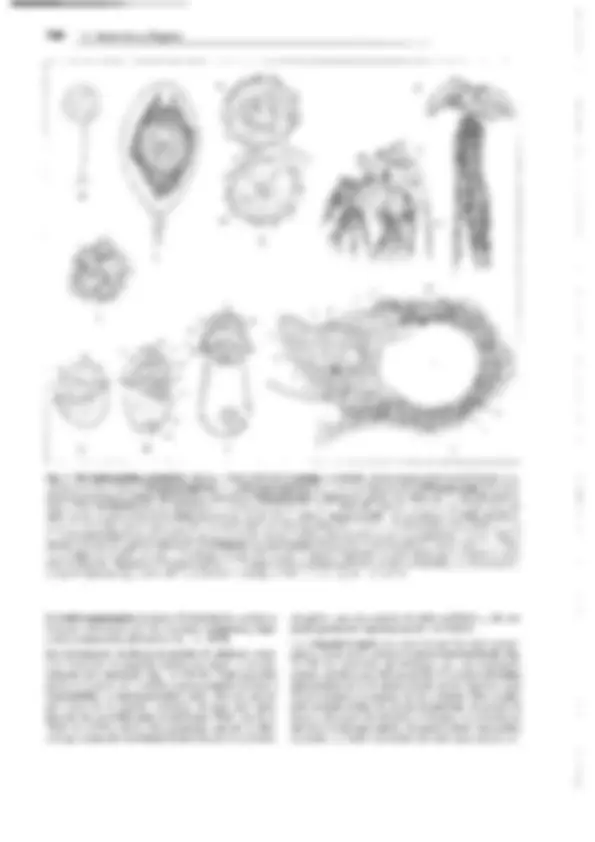
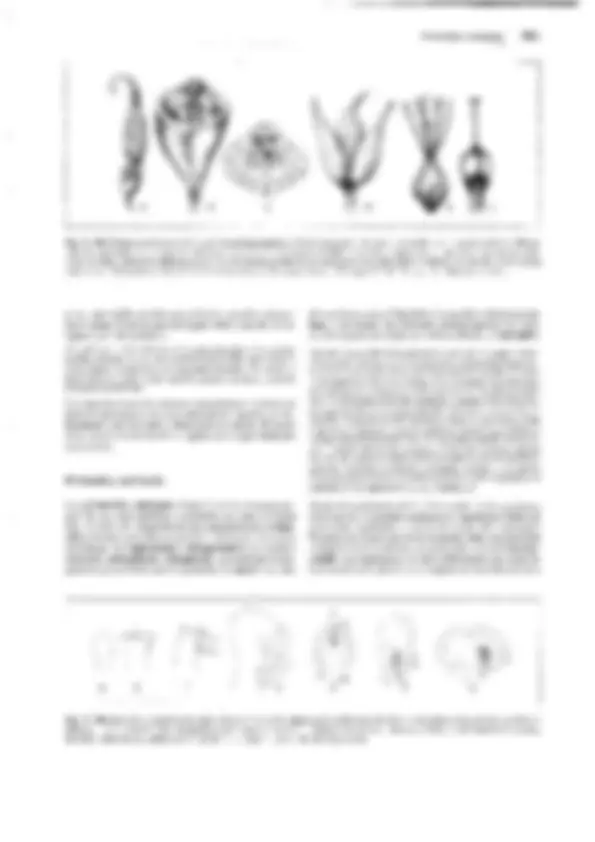
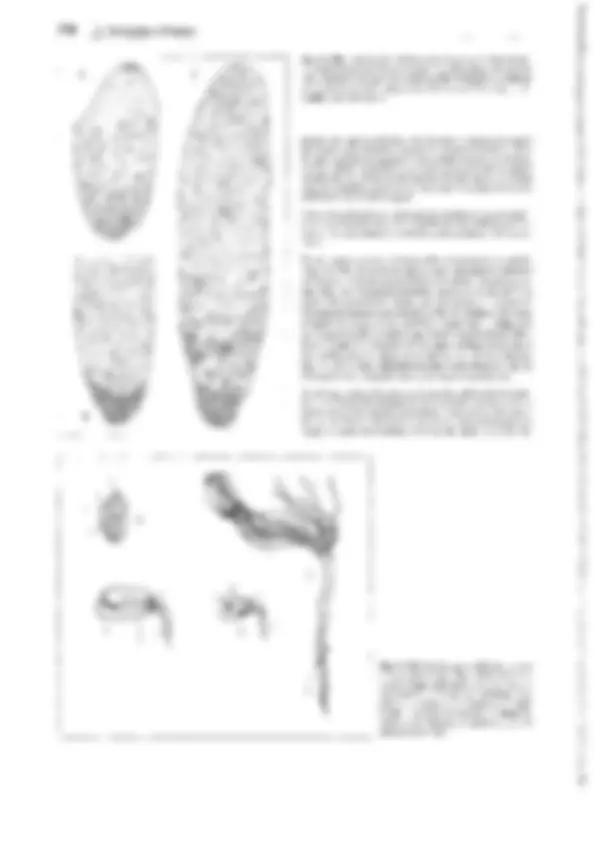
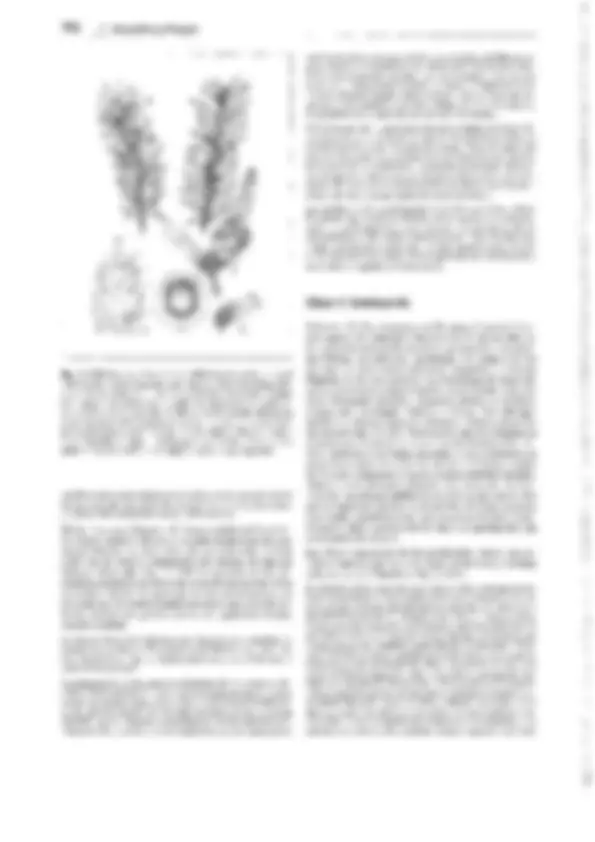
Fig. 11-128: Pteridophytina, Pteridopsida.
A, 8 Dryopteris filix mas. A Protalo (cara inferior)
con arquegonios ar, anteridios an y rizoides r. 8 Protalo con un esporófito joven (5x). C-G De- sarrollo del esporangio en los helechos
(C-E Asplenium, 300x; F, G Polypodium, 200x). C
Primeras divisiones del primordio originado a partir de una célula epidérmica. D División del primordio en un estrato parietal periférico w y una célula central as (arquesporio), que ya ha
producido una célula de tapete t. E El arquespo-
rio se ha dividido en células de tapete y tejido esporógeno sp. F Las células parietales w en- gruesan y forman el anillo; las del tapete desa- parecen, y las células madres de las esporas sm
forman las tétradas de las esporas. G Esporangio
maduro con esporas. H Esquema del desa- rrollo de un helecho; líneas rojas: fase ha- ploide, líneas negras: fase diploide. 1 espora, 2 protalo con gametangios 9 y cJ, 3 protalo con esporófito joven, 4 esporófito (muy reducido), con soros esporangiales, 5 esporangio no madu- ro (muy aumentado), de un soro, 6 esporangio maduro, con tétradas de esporas, 7 esporas.
A-B según H. Schenck; C-E: según R. Sadebeck;
F-H: según R. Harder. - an anteridio, ar arque- gonio, as arquesporio, b primera hoja, r rizoides, s esporas, sg tejido esporógeno, sm células ma- dres de las esporas, t célula del tapete, w raíz, wa capa parietal periférica, células de la pared
(anillo), 1-5 tabiques formados sucesivamente 1,
G gametófito, Sesporófito, R! división reductiva.
trata de un órgano sui generis, que no puede derivarse de
telomas. El proceso de la incurvación puede observarse,
p. ej., en los ejes esporangíferos de los equisetos (L, M;
fig. 11-141 F).
Desarrollo del embrión. En los pteridófitos actuales, tras
la primera división de la ovocélula fecundada se diferen-
cia ordinariamente, además de un haustorio (pie), un ápi-
ce radical, un ápice caulinar y un ápice foliar, que, al cre-
cer, primero todavía con el embrión unido al protalo (figs.
11-128 B, 11-155), se transforman después, respectiva-
mente, en la primera raíz, el tallo y la primera hoja (coti-
ledón). La posesión de raíces es característica para lama-
yoría de los pteridófitos. El extremo del eje de la plántula
opuesto al «polo caulinar» recibe el nombre de «polo
radical», pero sólo en los espermatófitos se desarrolla a
partir de él la raíz primaria (fig. 3-1 ), mientras que, en los
pteridófitos, la primera raíz aparece como formación en-
720 i 11 Sistemática y filogenia
dondean, se separan unas de otras y representan a las cé-
lulas madres de las esporas (generalmente 16); éstas, me-
diante meiosis, dan lugar, cada una, a cuatro meiósporas
haploides a menudo dispuestas en tetraedro.
En la periferia del tejido esporígeno se encuentran a me-
nudo células dispuestas en varios estratos sobre la pared
del esporangio; estas células proporcionan alimento a las
esporas y forman, conjuntamente, el tapete (E: t). Las cé-
lulas de un tapete de secreción secretan su contenido a
través de sus paredes. En el tapete plasmodial se disuel-
ven las paredes celulares y se liberan los protoplastos,
que se unen formando un periplasmodio. Entonces, éste
se sitúa entre las esporas jóvenes, que se disocian de las
tétradas, las nutre, participa en la formación de las mem-
branas de dichas esporas (perisporio) y acaba por consu-
mirse (F, G).
Esporas: la pared celular se divide en un endosporio, in-
terno, y un exosporio, resistente y externo, sobre el cual se
deposita el perisporio, de aspecto distinto. Las esporas ca-
recen casi siempre de clorofila y son de color pardusco a
amarillo.
En la mayor parte de los pteridófitos (los más primiti-
vos), todas las esporas tienen la misma constitución y,
cuando germinan, dan origen a un protalo en el cual se
forman juntos anteridios y arquegonios. Pero en algunos
casos, los protalos pueden ser dioicos, y la separación
de los sexos conduce entonces, en algunos grupos de
pteridófitos, al desarrollo de dos formas de meiósporas:
megásporas (= macrósporas), ricas en sustancia de re-
serva, nacidas de megasporangios (= macrosporangios)
y que al germinar sólo producen protalos femeninos, re-
lativamente grandes, y micrósporas, formadas en mi-
crosporangios, las cuales originan protalos masculinos,
menores (fig. 11-137). Con arreglo a lo que acabamos
de exponer tendremos que establecer una distinción en-
tre estirpes con esporas iguales (isospóreas) y diferen-
tes (heterospóreas), que no puede hacerse extensiva,
sin embargo, a la división del conjunto, puesto que se
produce del mismo modo en clases sistemáticamente
distintas.
Clase 1: Psilophytopsida
Los psilofitópsidos (extinguidos) son el grupo originario
de los pteridófitos. Su cuerpo vegetativo se compone de
telomas, que, en las familias primitivas, son lisos, mien-
tras que están cubiertos de emergencias en las de posición
más elevada. Los esporangios se presentan en posi-
ción apical o lateral en las ramas principales o laterales.
Todos los géneros son isospóreos. Todavía carecen de ver-
daderas raíces y están provistos de rizomas y rizoides. En
el tallo sólo se forma una sencilla protostela o actinostela.
Domina la isosporia.
Los psilofitópsidos son las plantas terrestres más antiguas
provistas de haces conductores y de estomas. Aparecieron
en el tránsito del Silúrico al Devónico (es decir, hace unos
400 millones de años), alcanzaron rápidamente una gran
riqueza de formas y se extinguieron ya al comienzo del
Devónico superior. Sus representantes morfológicamente
más primitivos pertenecían al
Orden 1: Rhyniales, que tenían un cuerpo vegetativo formado por
telomas dicótomos desnudos con esporangios todavía terminales.
Al género Caaksania, que se desarrolló entre el Silúrico superior
y el Devónico inferior, pertenece la planta terrestre más antigua
encontrada hasta ahora. El teloma, de 10 cm de altura y de ramifi-
cación dicótoma, estaba atravesado por una protostela y tenía es-
porangios que eran más largos que anchos.
Rhynia (incl. Aglaaphytan, antes atribuido a Rhynia), el género
que ha dado nombre las rhyniáceas, con dos especies halladas del
Devónico medio de Escocia (fig. 11-130 A, B), medía 1-2 m de al-
tura. Al menos en una especie (fig. 11-130 B), el esporófito se ele-
vaba sobre un^ «rizoma» subterráneo de crecimiento horizontal,^ ca-
rente de raíces y provisto de rizoides sin tabiques transversales.
Constaba de tallos cilíndricos, erectos, con ramificación dicótoma
y sin hojas.
Los tallos poseían cutícula y estomas de estructura relativamente
sencilla (v. 3.2.2.1) y eran evidentemente órganos asimiladores.
Así pues, Rhynia era una planta terrestre y formaba poblaciones
parecidas a juncales. El haz conductor se componía de hidroides,
como Aglaaphytan (Rhynia) majar y Harneaphytan, o ya entra-
queidas, con engrosamientos muy sencillos (anulares y helicoida-
les; en Rhynia gwynne-vaughanii). Formaba una protostela (fig.
A del cuadro 4-3, fig. 11-130 C), en parte ya con metaxilema; en
el tejido externo del haz, es decir, en el floema, no había aún cé-
lulas cribosas típicas con campos cribosos. No existía tampoco el
engrosamiento secundario. Los esporangios, cilíndricos o clavi-
formes y relativamente grandes, se hallaban situados en los ex-
tremos o los flancos de los tallos, tenían las paredes constituidas
por varios estratos celulares y se abrían por una fisura longitudi-
nal; su contenido consistía en abundantes tétradas de isósporas
(fig. 11-130 D, E).
En Rhynia gwynne-vaughanü aparecieron las ramificaciones late-
rales, producidas por culminación; igualmente, los esporangios
eran superados a veces (fig. 11-130 A).
En Harneaphytan, de aspecto parecido al del género Rhynia, la es-
tructura de los esporangios oblongos, reunidos en grupos de 2-4,
recuerda a la del esporogonio de Sphagnum; el tejido esporígeno
se aboveda en forma de campana sobre una columela formada por
células alargadas; los esporangios se abrían por un poro apical.
Debido a la carencia de traqueidas, ni Aglaaphytan (Rhynia) ma-
jar ni Harneaphytan se consideran ya plantas vasculares.
Las especies del Devónico inferior, extendidas por todo el mun-
do, del
Orden 2: Zosterophyllales (-ceae). constaban también de ramas
dicótomas desnudas, pero sus esporangios, laterales y con una
abertura transversal preformada, solían estar unidos formando es-
pigas (fig. 11-130 H). Se considera el grupo antecesor de los equi-
setópsidos y se ordena frecuentemente con ellas por este motivo.
En algunos géneros (p. ej., Zasterophyllum) se pudo ver que el ga-
metófito representaba una pequeña planta con ramificación estre-
llada (fig. 11-130 F), que formaba sobre los esporangióforos ar-
queados ascendentes sombrillas con arquegonios centrales y
anteridios periféricos. El joven esporófito originado tras la fecun-
dación se separaba del gametangióforo y se desarrollaba de mane-
ra independiente (fig. 11-130 G, H). Por lo tanto, no es correcto
considerar gametófitos a los «rizomas» de las rhyniáceas, pues di-
chos gametófitos permanecen unidos toda la vida al esporófito
como ocurre en los briófitos.
Así pues, estos pteridófitos primitivos tenían una alternancia de
generaciones anisomorfa y la fuerza de las generaciones era
aproximadamente igual. A partir de entonces, la vía filogenética
tanto hacia los briófitos (predominio del gametófito) como hacia
los pteridófitos derivados (predominio del esporófito) quedó
abierta por igual.
Como en muchas algas (p. ej., Halicystis-Derbesia, clorófitos),
también aquí las diferentes generaciones recibieron al principio
Clase: lycopodiopsida, licopodios 721
D
H
B E
G
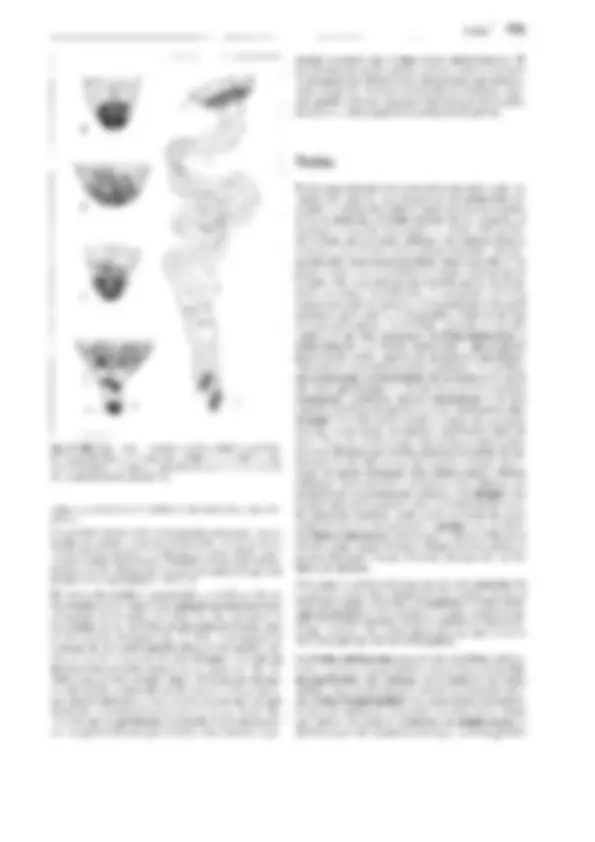
F
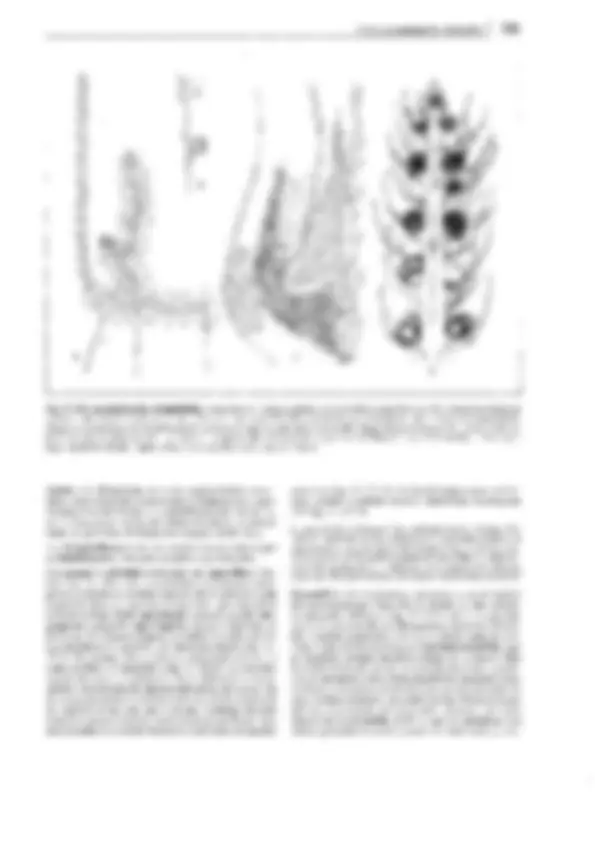
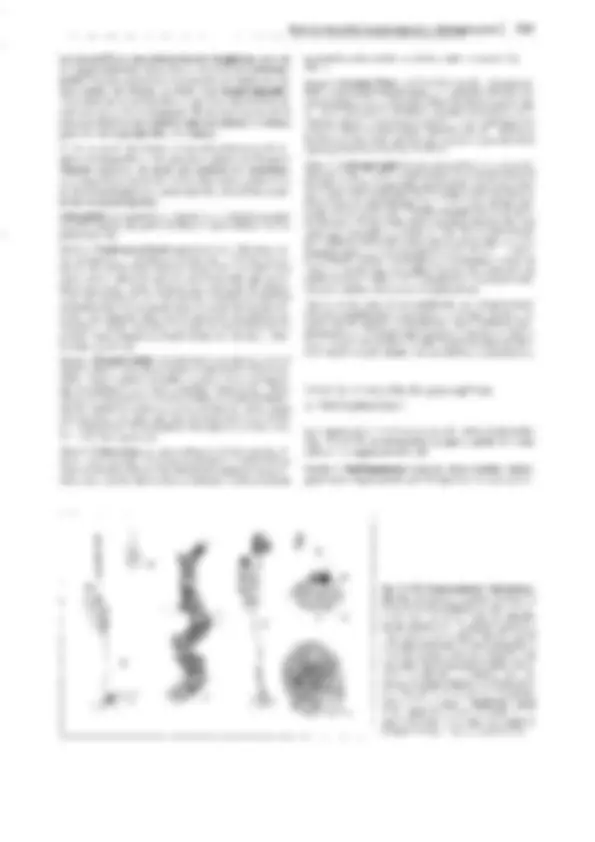
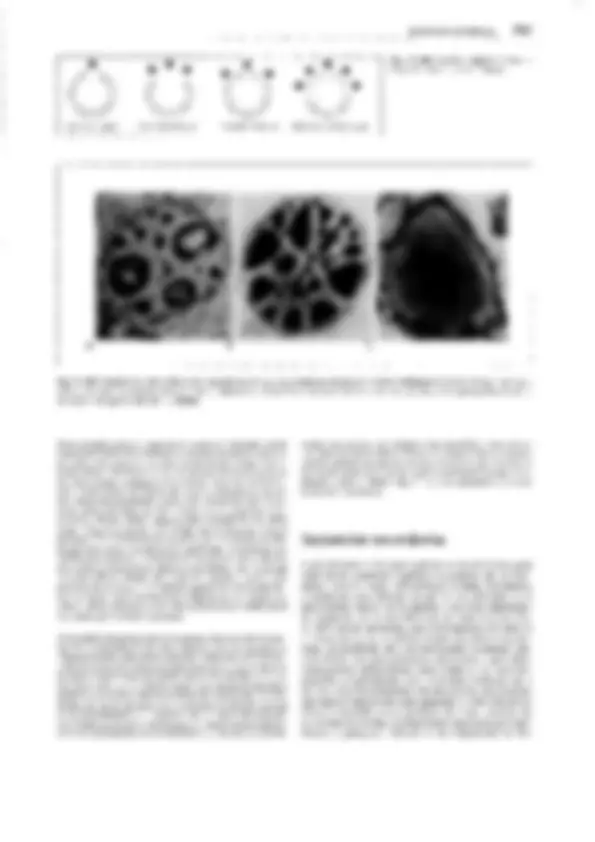
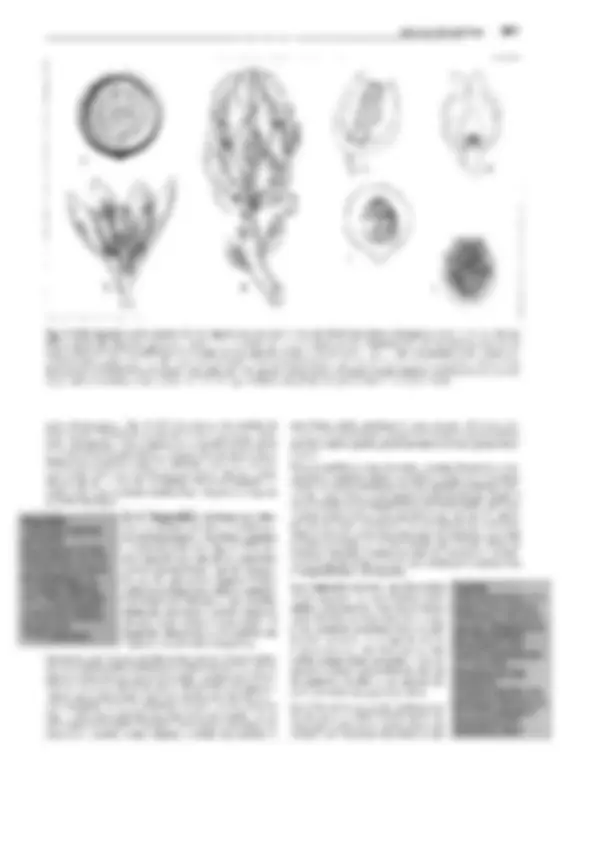
Fig. 11-130: Psilophytopsida. A-E Rhyniales, A Rhynia gwynne-vaughanii, reconstrucción (1/4x). B Aglaophyton (Rhynia) majar, reconstrucción
(1/Sx); C-E Rhynia, C sección transversal pulimentada del tallo, con la protostela visible (50x). D Sección pulimentada longitudinal de un esporangio
(2x). E Tétrada de esporas (100x). F-H Zosterophyllales. Zosterophy!lum rhenanum, reconstrucción (1/2x). F Gametófito (= Sciadophyton). G, H Es-
porófito, G joven, en conexión con el gametangióforo, H adulto con esporangios. -A-B: según Edwards; C-E: según R. Kidston y W.H. Lang; F, G: según R. Krausel y H. Weyland. a corteza externa, e epidermis, i corteza interna, p floema, x xilema.
distintos nombres genéricos antes de que se reconociera su perte-
nencia a un único y mismo ciclo vital. Así Taeniocrada corres-
ponde al gametófito estéril de la generación esporofítica de Zoste-
rophyllum, como Sciadophyton corresponde al gametófito fértil
del mismo (fig. 11-130 F). lyonophyton, con tallos rastreros y re-
dondos, cordón conductor central que conduce a los hidroides, y
gametangióforos erectos, ensanchados como platos, representaba
probablemente la generación gametofítica de Aglaophyton major
(fig. 11-130 B). Aquí podía reconocerse incluso la estructura del
anteridio con las células espermatógenas.
Más que el orden anterior se desarrollaron las especies que
existieron entre el Devónico medio y superior del
Orden 3: Trimerophylales (-ceae ). Formadas por ejes principa-
les extendidos (simpódicos) con ejes laterales aquí dicótomos o
trifurcados y con esporangios terminales y dispuestos en grupo.
El orden se interpreta como grupo genealógico de las «progim-
nospermas» (fig. 11-288). Dawsonites, al contrario que Trime-
rophyton, tenía los esporangios dirigidos hacia fuera y sobre
ejes curvos. Psilophyton ornatum presentaba en sus ejes excres-
cencias (fig. 11-131 A) y condujo, por ello, a los licopodióp-
sidos.
Como plantas terrestres primitivas, los psilofitópsidos
constituyen el grupo inicial del cual derivan filogenética-
mente los restantes pteridófitos y tal vez incluso hasta las
mismas gimnospermas.
Clase 2: Lycopodiopsida, licopodios
El esporófito, de ramificación a menudo dicótoma, pre-
senta hojas indivisas, pequeñas o estrechas (= microfilos ),
dispuestas helicoidalmente la mayoría de las veces. A ex-
cepción de algunas formas fósiles extinguidas, los espo-
rangios aparecen solitarios y en posición adaxial sobre o
junto a la base de las hojas (esporofilos ), que muy a me-
nudo están unidas formando grupos terminales de espo-
rofilos («flores»). Según la teoría telomática, la disposi-
ción foliar de los esporangios puede haberse originado tal
como aparece en la fig. 11-133. La heterosporia está muy
extendida junto con la isosporia. Los espermatozoides ra-
El mesofilo de L. clavatum es sencillo; sólo en contadas especies
se puede distinguir ya una diferenciación en parénquima en empa-
lizada y parénquima esponjoso. La epidermis foliar no presenta
cloroplastos. Como pasa siempre en los casos de dicotomía, la ra-
mificación de los ejes no está en relación con las hojas.
El haz conductor es una plectostela muy dividida que
puede derivarse de una actinostela (v. fig. A del cua-
dro 4-3) con células cribosas en el floema que presentan
campos cribosos en las paredes longitudinales, pero toda-
vía no poseen auténticas placas cribosas. En la sección
transversal del tallo, esta plectostela aparece rodeada ha-
cia fuera por una vaina de células no lignificadas, cuya
capa más externa contiene almidón; sigue una endodermis
de uno a dos estratos, con lignina en las delgadas paredes
celulares; la endodermis, en este caso, como en todos los
pteridófitos, es la capa más interna de la corteza. La cor-
teza externa consta de células esclerenquimáticas muy lig-
nificadas (fig. 11-134 L).
Una parte de las ramas es negativamente gravitrópica. A
menudo, por encima de una región más pobre en hojas, los
esporofilos se disponen en densas agrupaciones espicifor-
mes (flores) (fig. 11-134 G). El ápice caulinar se consume
en su formación, de modo que el grupo de esporofilos
constituye el extremo del tallo. Los esporofilos (H) tienen
la forma de escamas amplias y llevan en la base de su haz
o cara superior un esporangio grande, deprimido y arriño-
. .
A
((~ (l)j'~_
B
e
D
Clase: lycopodiopsida, licopodios 1 723
nado, en el que se hallan numerosas meiósporas, todas del
mismo tamaño (isósporas) (J, K). Del borde de los espo-
rofilos penden lóbulos membranosos que, como «indu-
sio», protegen el esporangio inferior vecino.
La pared del esporangio se compone de varios estratos ce-
lulares externos; a ellos se une hacia dentro un tapete de
secreción. El esporangio se abre en dos valvas por medio
de una cisura longitudinal que parte del ápice, a lo largo de
una línea que puede percibirse ya en la estructura anató-
mica de las células. Las esporas permanecen unidas, for-
man tétradas, hasta su madurez; su exosporio, formado
por varias capas, muestra líneas reticuladas de engrosa-
miento (fig. 11-134 J, K). En la naturaleza sólo germinan
después de pasados 6 o 7 años y producen, en un princi-
pio, a expensas de sus propias reservas, un embrión de
cinco células (fig. 11-135 A), y éste, después de un perío-
do de reposo, prosigue su desarrollo únicamente cuando
penetran en sus células basales filamentos fúngicos, para
formar una especie de micorrizas (B: p).
Los protalos (figs. 11-134 E, F; 11-135) viven subterrá-
neos y constituyen tuberculitos blanquecinos, saprófitos;
son cuerpos de forma variada según las especies, con ló-
bulos prominentes, hasta de unos 2 cm de diámetro y pro-
vistos de largos rizoides tubulares, encargados de captar el
agua. Los hongos micorrizógenos (fig. 11-135 B, C), que
H
K
L
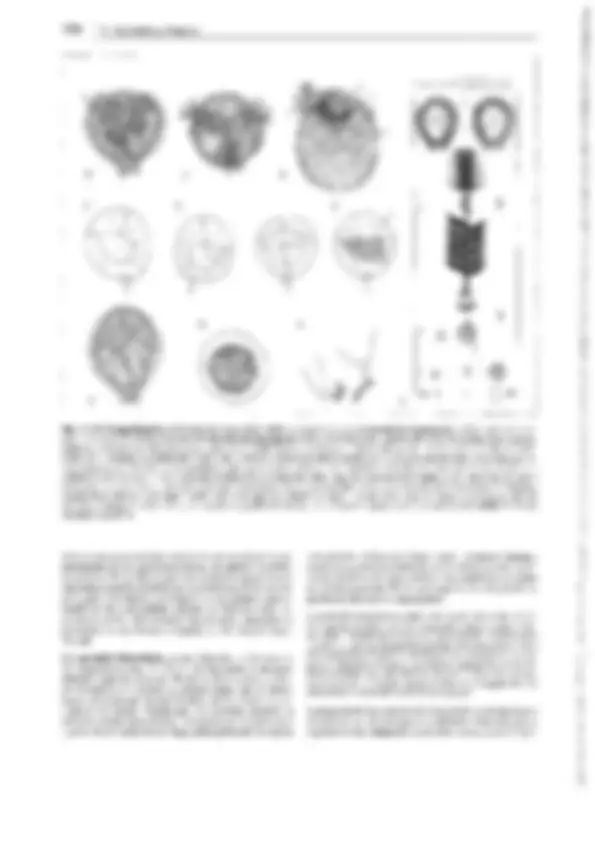
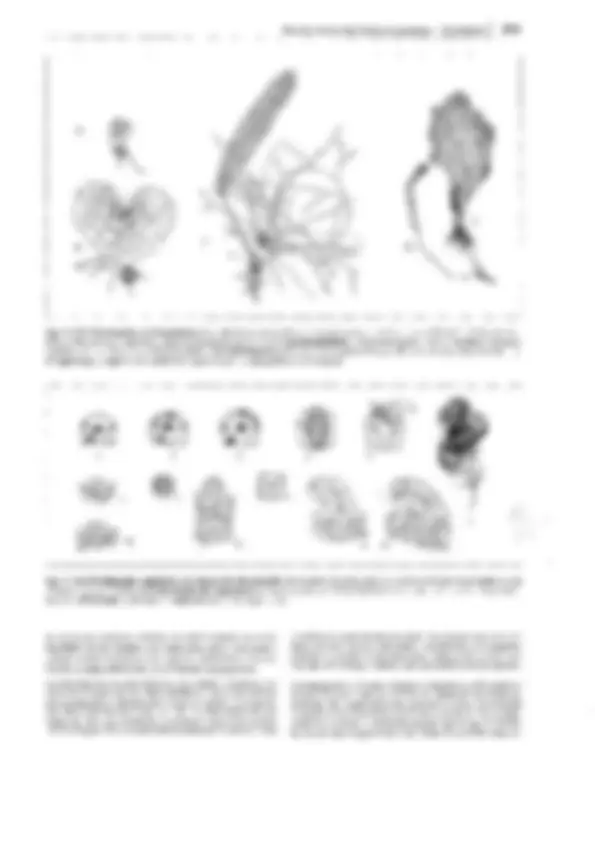
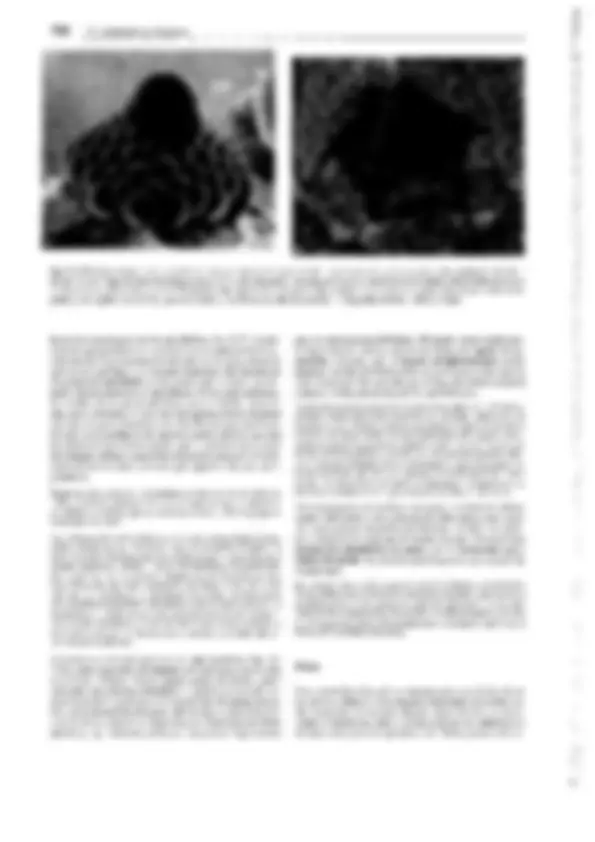
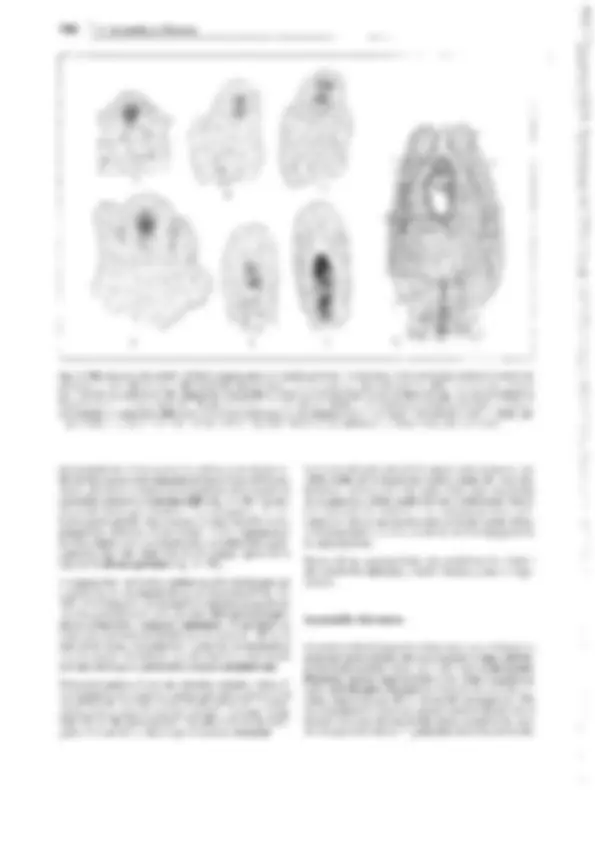
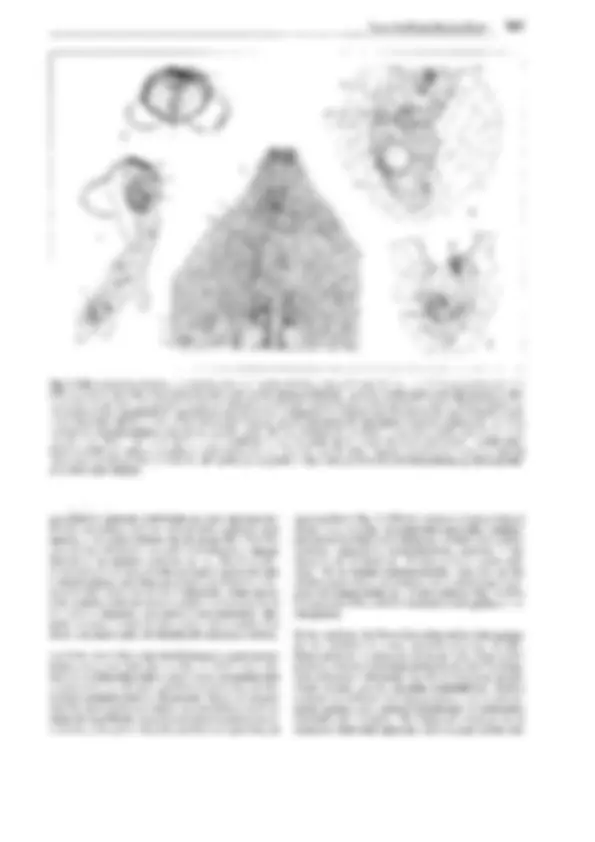
Fig. 11-134: lycopodiopsida, lycopodiales, Lycopodium c!avatum. A Anteridio, todavía cerrado, sección longitudinal (75x). B Espermatozoides
(400x). C Arquegonio joven, todavía cerrado, D abierto y maduro para ser fecundado (75x). E Protalo viejo (2x). F Protalo con planta joven (3/4x). G
Planta con grupo (estróbilo) de esporofilos (1/3x). H Esporofilo con esporangio abierto (8x). J, K Esporas en dos posiciones (400x). l Sección transver- sal a través del tallo (1 Oüx). -A-F: según H. Bruchmann; G-H: según H. Schenck. b base foliar con cavidad h, e epidermis, en endodermis, h cavidad, p floema, r corteza, s vaina amilácea, x xilema.
724 j 11 Sistemática y filogenia
viven en las capas celulares periféricas, desempeñan, sin
duda, un papel importante en su nutrición. En condiciones
naturales, la madurez sexual no se presenta hasta transcu-
rridos de 12 a 15 años, debiendo comprender la vida total
de los protalos unos 20 años. En cambio, en cultivo puro
artificial, en ausencia de bacterios, todo el desarrollo se
realiza en pocos meses. En algunas especies, los protalos
sobresalen del suelo por su parte superior y allí toman co-
lor verde. Los protalos son monoicos, y llevan general-
mente hacia su parte apical numerosos órganos sexuales
(figs. ll-134A-D, 11-135 C: ag). Los anteridios (a) están
un poco hundidos en el tejido y son pluricelulares; cada
una de sus células, exceptuadas las parietales, produce un
espermatozoide (fig. 11-134 B) oval, que tiene debajo de
p
2~-e
D
ba
SS e
E 4
Fig. 11-135: Lycopodiales, A, B Lycopodium, L. annotinum. Desarrollo
del protalo. A Espora en germinación, incolora, quinquecelular con mem-
brana sh, célula rizoidal r, célula basal b, célula apical s (580x). B Plántu-
la joven, con las células inferiores habitadas por un hongo endófito p; la
célula apical se ha dividido en tres células meristemáticas apicales (s 1 , s 2 ,
s) (470x). C-E Diphasium, D. complanatum. C Protalo maduro con ante-
ridios a, arquegonios ag y células fungíferas (en negro) (24x). D-E Desa-
rrollo del embrión. D Embrión con las primeras divisiones; la pared basal
1 separa el primordio del embrióforo e del del cuerpo embrionario; los ta-
biques transversales 2 y 3 (el último en el plano de corte), así como el ta-
bique transversal 4, proporcionan dos estratos tetracelulares, de los cua-
les el situado entre 1 y 4 produce el haustorio y el inferior, la parte del
vástago. E Fase intermedia (112x). Según H. Bruchmann. - a anteridio,
ag arquegonio, b célula basal, ba, primordio foliar, e embrióforo, p hon-
go, r célula rizoidal, s célula apical, célula del meristema apical, sh mem-
brana de la espora, ss ápice del tallo.
su ápice sólo dos flagelos. Los arquegonios (figs. 11-
C, D, 11-135 C: ag), igualmente hundidos, poseen de or-
dinario numerosas células del canal del cuello (hasta 20,
aunque pueden reducirse a una); las células supremas de
la pared son desviadas en la dehiscencia. De la ovocélula
fecundada surge, tras varias divisiones celulares, un em-
brión, cuyo suspensor (e) lo introduce hacia el tejido del
protalo. La fig. 11-135 E muestra el desarrollo de un haus-
torio, que absorbe el alimento del protalo, y de la primera
hoja, que persiste en forma de escama (ba). La primera
raíz tiene un aspecto caulógeno.
En Lycopodium, los esporofilos se agrupan en espigas que se ele-
van sobre cortas ramas laterales; en Huperzia, trofofilos y esporo-
filos se forman alternativamente siguiendo el ritmo anual sobre
vástagos erguidos y bifurcados. Diphasium posee espigas de espo-
rofilos como Lycopodium, pero sus vástagos son planos, dorsiven-
trales, con hojas escuamiformes.
Las especies de Lycopodites, del Devónico superior, se
parecían ya mucho a los representantes actuales de la fa-
milia. Así pues, la forma de los licopodiópsidos se ha
mantenido sin cambios a lo largo de más de 300 millones
de años.
Mientras que hasta ahora predominaba la isosporia, los
siguientes órdenes han progresado hacia la heterosporia.
En las axilas de las hojas se encuentra una pequeña ex-
crescencia lingüiforme: la lígula (fig. 11-136 C).
Orden 4: Selaginellales. Por su hábito, las especies herbá-
ceas de Selaginella se parecen un poco a los musgos. Sin
embargo, por sus características generativas y anatómicas
son verdaderos pteridófitos y, por lo tanto, también verda-
deros cormófitos. En Europa, este género sólo está repre-
sentado por unas cuantas especies, pero en los trópicos po-
see unas 700.
Hábito: Selaginella tiene en parte los tallos tendidos, en
parte erguidos, pero en todos los casos muy ramificados,
con dicotomía; unas veces forman céspedes y otras trepan
por la maleza con tallos de varios metros de longitud.
El tallo se halla cubierto de hojas escuamiformes dis-
puestas en cuatro series, helicoidales o con mayor fre-
cuencia decusadas, generalmente dorsiventrales, con dos
series de hojas pequeñas en su parte superior, y otras
dos series de hojas mayores, opuestas a las primeras, en
su porción inferior (figs. 4-67 B, 11-136 A; anisofilia,
v. p. 193). Las hojas tienen un solo nervio medio no rami-
ficado y raramente poseen, además de parénquima espon-
joso, también parénquima en empalizada; en algunas
especies, las células del mesofilo contienen un solo cloro-
plasto grande, cupuliforme. Las hojas de Selaginella pre-
sentan en la base de su cara superior una pequeña
escama membranosa que carece de clorofila y sale de la
epidermis: la lígula (fig. 11-136 C), que como órgano cap-
tador de agua facilita la rápida absorción de las precipita-
ciones por los brotes foliosos y, en algunas especies, em-
palma con el haz conductor por medio de traqueidas.
En las bifurcaciones del tallo se originan de manera exó-
gena, en muchas especies, unas ramas cilíndricas, alarga-
das, pero incoloras y afilas, los rizóforos (fig. 11-136 A:
w), que crecen hacia abajo, se dividen por dicotomía y en
cuyo extremo libre nacen endógenamente fascículos de
raíces. En condiciones apropiadas, los rizóforos pueden
dar lugar a hojas, como los tallos. La disposición de los
haces conductores oscila entre la protostela central, la
726
k
h
K
D E F
p p p^ p
G H
\k (^) L
ºººº 6 0000
~R'te&
ws@
4 s
~
s 3 G
(^2) ®d G (^) ~
o
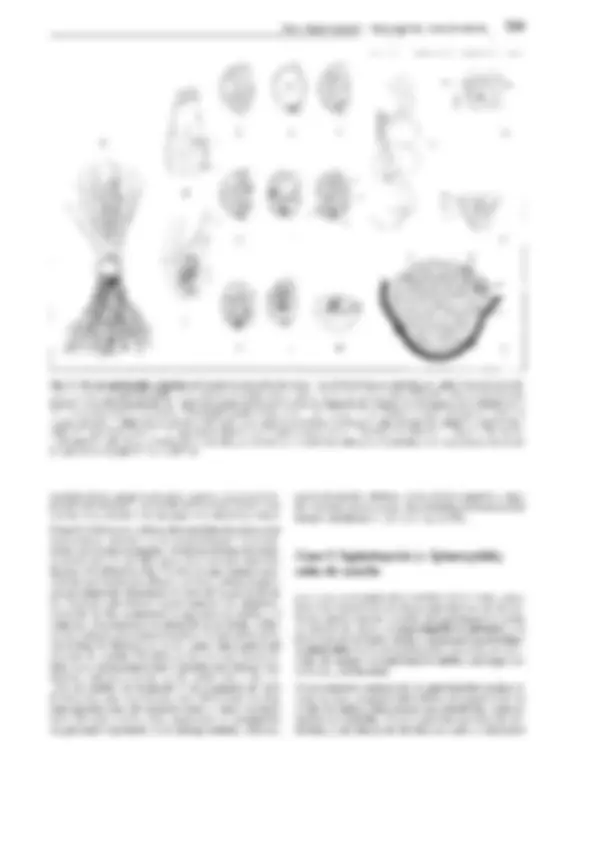
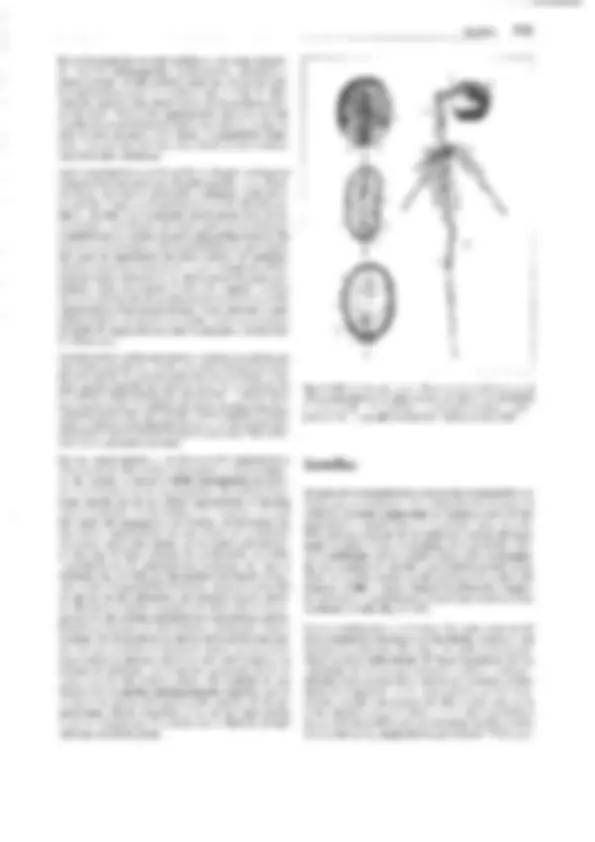
Fig. 11-137: Lycopodiopsida, A, B 5elaginella inaequalifo/ía. A Megasporangio con una sola tétrada de megásporas y células madres de las es-
poras atrofiadas (70x). B Microsporangio con tétradas de micrósporas. C-G 5. stolonífera (640x). Germinación de las micrósporas, fases sucesivas;
célula del protalo (p), que debe considerarse como rizoidal; C, D, F vista lateral, E dorsal. En G, la célula protálica no se ve y las parietales están des-
truidas. H 5. cuspídata, espermatozoides (780x). J-K 5. martensíí. J Megáspora abierta, protalo con 3 protuberancias rizoidales y varios arquegonios;
visto desde arriba ( 112x). K Sección longitudinal, 2 arquegonios con los embriones desarrollándose, embríóforo (e), haustorio (h), rizóforo (w), hojas
embrionarias (k) con lígula (150x). L Esquema del desarrollo de 5elagínella; líneas rojas: fase haploide, líneas negras o todo negro: fase diploide; 1
meiósporas; 2 lo mismo tras la formación del protalo; 3 megáspora y protalo con esporófito germinado. 4 esporófito (S); 5 esporangios; 6 meiósporas
después de ser soltadas. -A-8: según J. Sachs, modif.; C-H: según W.C. Belajeff; J, K: según H. Bruchmann; L: según R. Harder. - a células parietales del
anteridio, e embrióforo, h haustorio, k hojas germinales, p célula del protalo, s células espermatógenas, t tapete, w rizóforo, G gametófito, R! división
reductiva, S esporófito.
tituyen una capa mucilaginosa en la que se alojan las es-
pennátidas (F, G); queda persistente, en cambio, la celuli-
ta protálica (F: p ). Hasta aquí todo el protalo masculino se
encuentra todavía envuelto por la membrana de la micrós-
pora, pero, finalmente, se rompe ésta y los gámetas proce-
dentes de las espennátidas quedan en libertad como es-
permatozoides débilmente encorvados, mazudos y
provistos en su extremo delgado de dos largos flage-
los (H).
Los protalos femeninos, no tan reducidos, se forman en
las megásporas (fig. 11-137 J). Su desarrollo es un poco
distinto según las especies. El núcleo de la espora se divi-
de libremente en numerosos núcleos hijos, que se distri-
buyen en el plasma parietal del ápice de la espora y se en-
vuelven en sendas membranas; se continúa después la
división celular hacia la base. Así acaba por llenarse poco
a poco desde arriba hacia abajo principalmente la espora
con grandes células protálicas; pero, al mismo tiempo,
empieza a producirse también, en el mismo sentido, la di-
visión sucesiva de estas células y se constituye un tejido
de células pequeñas. En la parte superior de este protalo se
producen unos pocos arquegonios.
La pared de la megáspora se abre sobre sus tres aristas (fig. 11- J); el protalo incoloro y de células pequeñas emerge un poco y for- ma, sobre 3 protuberancias hísticas, algunos rizoides que sirven para absorber el agua. A continuación se produce la fecundación de uno o más arquegonios. El zigoto se divide por su primera pared en un sus- pensor (embrióforo, K) que se curva hacia el arquegonio y en un ver- dadero embrión, que, para liberarse del protalo, tiene que curvarse hacia fuera (K: e); después queda incluido en el megaprotalo que nuevamente se encuentra dentro de la megáspora.
La mayoría de las especies de Selaginella viven tapizando
el suelo en las selvas tropicales húmedas. Sólo unas pocas
especies se han adaptado a ambientes secos, como S. !epi-
727
B D
A
Fig. 11-138: Lycopodiopsida, Lepidodendrales. A, B Sigillaria, A reconstrucción (1/80x), B Pulvínulos foliares (2, 5x). C-E Lepidodendron. C re-
construcción (1/200x). D Pulvínulos foliares (tamaño natural). E Cono de esporofilos (tamaño natural). -A-C, E: según K. Magdefrau; D según Stur.
dophylla, de América central, cuyos vástagos dispuestos
en roseta se arrollan en tiempo seco (falsa «rosa de Jeri-
có»). Las especies de Selaginellites, herbáceas semejantes
a Selaginella, son del Carbonífero ya eran heterospó-
reas. Hace 300 millones de años el de las
actuales especies de Selaginella.
Orden 5: Lepidodendrales (Lepidófitos). Las lepidoden-
drales arbóreas alcanzaban hasta 40 m de altura y 5 m de
grueso (fig. 11-138). Tuvieron su máximo desarrollo du-
rante el Carbonífero (fig. 11-163) y han tenido una parti-
cipación importante en la formación de la hulla. Sus ho-
jas lineares, del tipo de los microfilos (pueden alcanzar,
sin embargo, 1 m), ordenadas helicoidalmente, tenían los
estomas dispuestos en dos surcos longitudinales situados
en el envés. Después de su caída dejaban en la superficie
del tronco cicatrices características y pulvínulos foliares
(fig. 11-138 B, D). La fijación de estos árboles se verifi-
caba por medio de rizóforos (A, C), ramificados repetida-
mente por dicotomía y dotados de engrosamiento secun-
dario, que corrían casi superficiales (suelo pantanoso); en
ellos se originaban de modo exógeno gran número de raí-
ces, relativamente débiles, de estructura particular (los lla-
mados apéndices), que luego se desprendían y dejaban ci-
catrices en gran cantidad, por lo que los rizóforos fueron
llamados estigmarios.
Las hojas eran recorridas por un haz conductor simple, raras veces bifurcado, y no tenían todavía tejido en empalizada. La señales que se reconocen en las cicatrices foliares en número de uno (fig. 11- B) o dos pares (D) indican el punto de salida de cordones de tejido
lagunar que servían para la aireación y transcurrían paralelos a los
rastros foliares en la corteza primaria. Los tallos poseían sifonoste- la (fig. 11-139 A); su floema, de membrana fina, estaba todavía poco diferenciado. Un anillo de cámbium no muy activo formaba nuevos tejidos que daban lugar a un engrosamiento secundario; las traquei- das escalariformes del leño secundario (con los engrosamientos constituidos de modo algo especial) presentaban un diámetro muy constante y el conjunto de ellas y de los radios medulares unistratos, que a veces ya se hallaban presentes, hace pensar en el leño de las coníferas actuales (aunque sin poros areolados y, como en casi todas las plantas que vivían en el hemisferio Norte durante el Carbonífe- ro, sin anillos anuales). Ciertamente, todo el leño secundario debía de tener escasa importancia para la estabilidad del árbol y la con- ducción de agua en el mismo. Los troncos contaban ya con un me- ristema equivalente al cámbium suberógeno, que secretaba, espe- cialmente hacia dentro, numerosos estratos celulares, de tal manera que se constituía una corteza extraordinariamente gruesa en relación con el diámetro del leño (en Lepidodendron, hasta el 99 % del cor- te transversal, fig. 11-139 A); dicha corteza estaba principalmente constituida por tejido de sostén, pero además debía tomar parte en el aprovechamiento del agua por medio de las lígulas, que todavía se conservaban durante algún tiempo después de la caída de la hoja.
Los troncos de las Sigillariaceae, las sigilariáceas arbóreas (fig. 11-138 A), estaban cubiertos de series longitudinales de pulvínu-
Clase: Equisetopsida (= Sphenopsida), colas de caballo 729
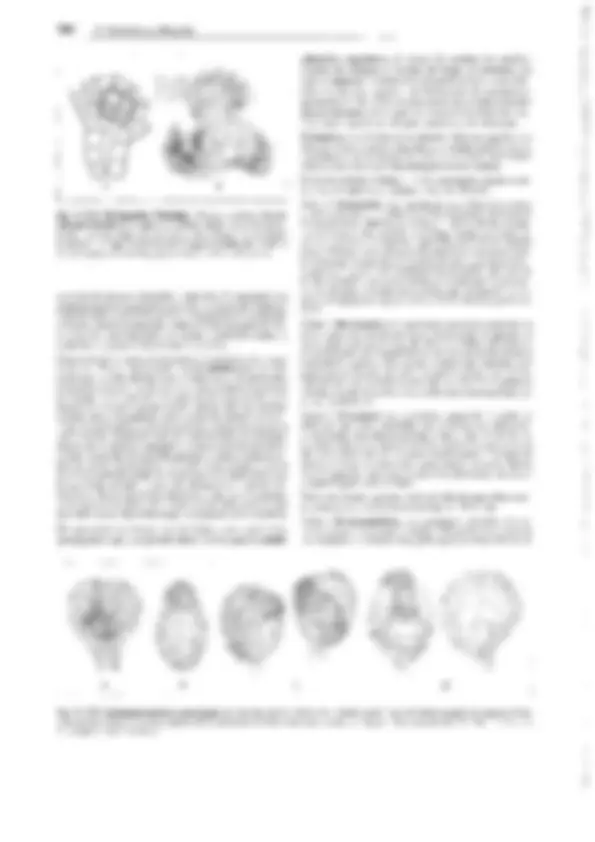
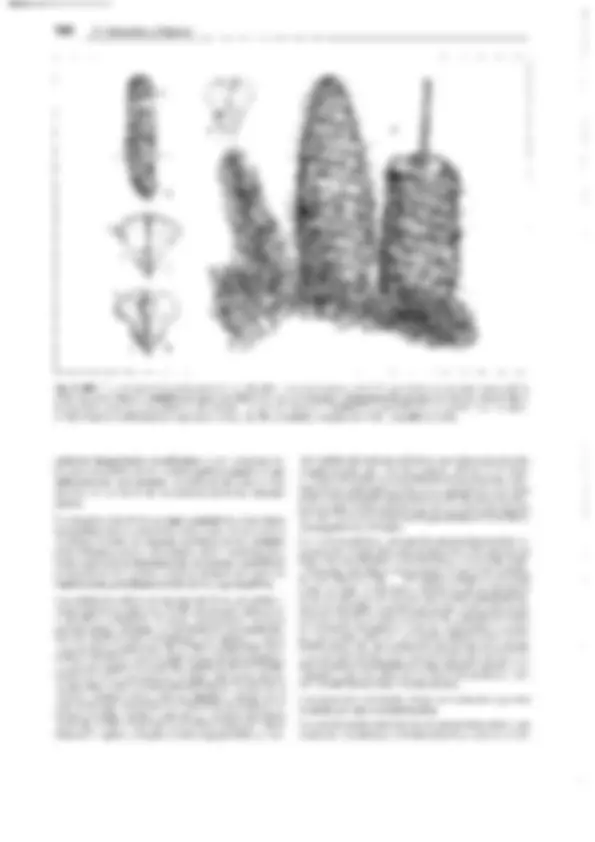
o E F^ Q
A N
B
h~~·^ hk^ ~
z G)
G H^ p
e
K L M
Fig. 11-140: Lycopodiopsida, lsoetales. A-C !saetes lacustris. A Planta entera (1/2x). B Porción basal de una hoja, con lígula y fóvea (2x). C La mis-
ma en sección longitudinal (4x). D-M l. setacea. Desarrollo del microprotalo y formación de los espermatozoides (500x). N /. ma/inverniana. Esperma-
tozoide ( 11 OOx). 0-Q Megaprotalo. O Protalo 9 de Stylites andicola dentro de la membrana rota de la espora, con arquegonios (ar), el derecho con la
célula del conducto ventral y la ovocélula (60x). P, Q Desarrollo del arquegonio a partir de una célula superficial en !saetes echinaspara, h células de
la pared del cuello, hk célula del conducto del cuello (250x). -A-C: según R. von Wettstein; D-M: según Liebig; N: según W.C. Belajeff; O: según W. Rauh
y Falk; P, Q: según D.H. Campbell. - ar arquegonio, b célula del canal ventral, e exina y, dentro, intina, f fóvea, h células de la pared del cuello, hk célu-
la del canal del cuello, li lígula, m micrósporas, o ovocélula, p célula protálica, s células espermatógenas, t trabéculas, w células parietales, z célula cen- tral, proporciona la célula del canal ventral (b).
(lepidodendrales), aunque relativamente grandes, y provistas de ho-
jas relativamente largas, y se continúa en Pleuromeia, Nathorstiana
y en los Stylites actuales, con una progresiva reducción del tronco.
Después de lo expuesto hasta ahora pueden relacionarse los
representantes actuales de los licopodiópsidos frecuente-
mente con formas extinguidas. Se desenvolvieron de mane-
ra mucho más considerable que en la actualidad, sobre todo
durante el Carbonífero (fig. 11-163), en que estaban repre-
sentados por numerosos géneros de formas arborescentes y
excepcionalmente alcanzaron el nivel de organización de
los vegetales seminíferos (lepidospermas). Las imperfec-
ciones de las vías conductoras de agua de estas plantas, así
como las concernientes a la absorción de la misma, debie-
ron de conducir a la desaparición de las formas arbóreas ha-
cia el final del Paleozoico, ya sea como consecuencia del
aumento de la aridez climática, ya debido a que fueran eli-
minadas al expansionarse tipos vegetales con sistemas con-
ductores más perfectos (p. ej., las cordaítidas; v. fig. 11-
163). En cambio, los licopodios y las selaginelas de porte
herbáceo han persistido durante unos 300 millones de años,
hasta nuestros días, sin variación notable («tipos persisten-
tes»). De todos modos, estos organismos no desempeñan
ningún papel importante en los paisajes actuales, mientras
que los licopodios arbóreos, junto con los calamites y algu-
nos helechos arborescentes, determinaban la fisonomía del
bosque carbonífero (v. 11.3.2.2, fig 11-295).
Clase 3: Equisetopsida (= Sphenopsida),
colas de caballo
Esta clase se distingue de la anterior (II) en varios carac-
teres: los esporofilos son claramente distintos de los tro-
fofilos. En las especies actuales, los esporangios se sitúan,
en número de varios, en esporangióforos peltados y no
en la axila de las hojas. Dichos esporangios poseen tape-
te plasmodial (en los licopodiópsidos, un tapete de secre-
ción). El vástago está articulado en nudos, con hojas ver-
ticiladas, y entrenudos.
Como caracteres comunes de los equisetópsidos pueden ci-
tarse: sus hojas pequeñas (microfilos), en comparación con
el tallo, las cuales, a diferencia de otros pteridófitos, están or-
denadas en verticilos. El tallo suele tener ramificación ver-
ticilada y está claramente dividido en nudos y entrenudos
730
L
D
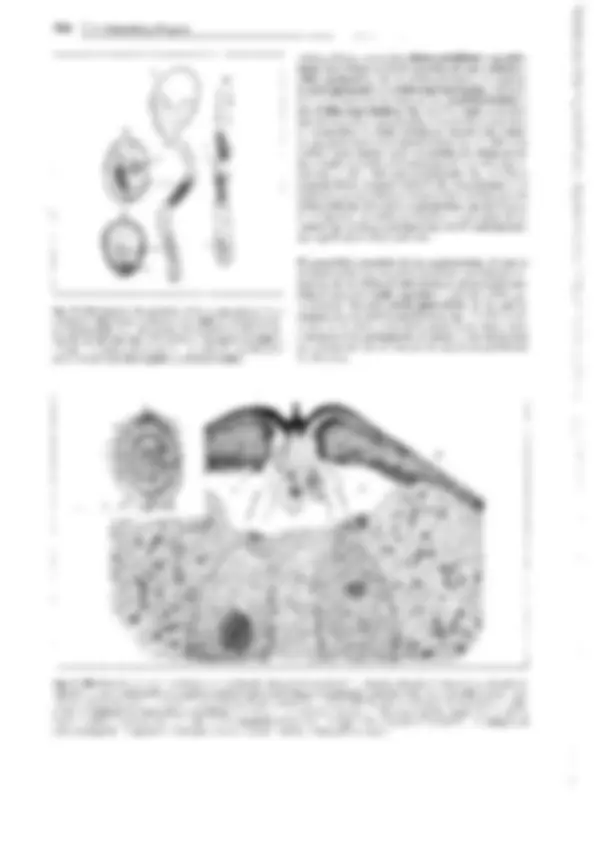
Fig. 11-141: Equisetopsida, Equisetaceae, Equisetum. A Protalo 9 con arquegonios (a), visto por la cara inferior (17x). B Espermatozoide (1250x).
C Embrión; 1, 2 tabiques que lo dividen en cuadrantes; de la mitad situada encima del tabique basal 1 se origina el tallo (s) y el primer verticilo de ho-
jas (b); de la mitad inferior, la raíz (w) y el haustorio (h) (165x). O Protalo 9 con plántula (dibujada más oscura), visto de lado, con los primeros vertici-
los de hojas y la raíz. E-L Equísetum arvense. E Vástagos fértiles (f) que brotan de un rizoma tuberoso, con vástago vegetativo (v) todavía en fase de
yema (1/2x). F y G Esporofilo con esporangios, abiertos en G (6x). H Espora con las dos bandas helicoidales (hapterios) del perisporio (360x). J Espora
con las bandas helicoidales extendidas en estado seco; un poco menos aumentada que en H (100x). K Vástago vegetativo, estéril (1/2x). L Sección
transversal del tallo; en los haces conductores, en negro el xilema y conducto carinal el, con cordones esclerenquimáticos (ss) en los surcos y costillas
( 16x). - A, D: según K. von Goebel; B: según L.W. Sharp; C: según R. Sadebeck; E-K: según H. Schenck.- a arquegonio, b verticilo foliar, b1, b2 los pri- meros verticilos foliares, c citoplasma, cg tejido clorofilífero, ck conducto carinal, e endodermis, f vástago fértil, g flagelo, h haustorio, k núcleo, m cavi-
dad medular lisígena, so fila de estomas, ss cordones esclerenquimáticos, s tallo, v vástago vegetativo, vk conducto valecular, w raíz.
732 j 11 Sistemática y filogenia
ración generalmente anual; éstos permanecen sencillos o se
ramifican en verticilo, cuyas ramas pueden a su vez formar
nuevos verticilos de segundo, tercer orden, etc. (fig. 11-141 ).
Los ejes, estriados, se componen de entrenudos alargados.
En los nudos se insertan, separados entre sí por los entre-
nudos, verticilos de microfilos escuamiformes acuminados
(v. 4.2.2), provistos de un haz conductor delgado y solda-
dos entre sí inferiormente en una vaina que rodea el tallo
(fig. 11-141 E). Los entrenudos, que crecen de modo inter-
calar, están envueltos en su base por dichas vainas. En los
nudos se encuentra siempre un anillo cerrado de haces con-
ductores con xilema interno y floema externo (sifonoste-
la). En los entrenudos, el anillo se descompone encordo-
nes vasculares, incluidos en el parénquina (eustela; L).
En un nudo están siempre juntos, en la parte inferior, los primordios de las ramas laterales y encima los rastros foliares, que constan de protoxilema; los últimos sólo penetran en las hojas en el nudo in- mediatamente superior. Los haces conductores junto con sus ras- tros foliares se disponen en orden inverso en los entrenudos conse- cutivos (como en la fig. 11-142 E). Las ramas laterales salen al exterior siempre atravesando las vainas y alternando con las hojas.
Dado el pequeño tamaño de las hojas, que pronto pierden
la clorofila, los tallos verdes se encargan de la asimilación.
Los haces conductores colaterales (v. 3.2.4.3) son muy
pobres en xilema. Las partes más viejas de dicho xilema
desaparecen pronto y dejan conductos intercelulares, que
en la sección transversal aparecen como un círculo de ca-
vidades carinales (fig. 11-141 L). También en la medula,
muy desarrollada, se origina un gran espacio intercelular,
que conduce aire (conducto central) y, asimismo, en la
corteza se forma un círculo de conductos valeculares (si-
tuados debajo de los surcos superficiales del tallo). Hacia
dentro, la corteza suele limitar con una endodermis de
1-2 capas y con banda de Caspary.
Las membranas externas de la epidermis del tallo se hallan más o menos fuertemente impregnadas de sílice en los equisetos, que se emplearon en otro tiempo por este motivo para pulir vasijas metá- licas. En los surcos, entre las estrías, están los estomas, siempre or- denados de dos en dos en filas longitudinales. Muestran particula- ridades que no existen en los demás vegetales: las células oclusivas están completamente recubiertas por sus células anexas; al aumen- tar la turgencia, las células oclusivas se redondean y el movimien- to se transmite a las células anexas por medio de los engrosamien- tos de la pared limitante; esto hace que el estoma se abra.
Los esporangios se originan sobre esporangióforos de
configuración especial, agrupados en varios verticilos al-
ternantes en los extremos de los vástagos, donde constitu-
yen, debido a un gran acortamiento de los entrenudos,
agrupaciones de esporofilos («flores»), en forma de estró-
bilo o piña (fig. 11-141 E). Los esporangióforos («espo-
rofilos») adoptan la forma de velador, en cuyo envés se
disponen 5-10 esporangios a modo de saco (F, G); sus ha-
ces conductores son concéntricos.
El tejido esporígeno se halla rodeado, en los esporangios jóvenes, por una pared pluriestratificada; pero el estrato más profundo de esta pared (tapete plasmodial) forma, al disolverse la pared de las célu- las, un periplasmodio, que se insinúa entre las esporas redondeadas y se emplea en la formación de las paredes de éstas, de modo que, en la madurez, únicamente se conservan los dos estratos más exter- nos como pared definitiva del esporangio; las células de la epider- mis presentan engrosamientos helicoidales o anulares en sus mem- branas. La dehiscencia se verifica según una grieta longitudinal, que se produce en la cara interna del esporangio mediante un mecanis- mo de cohesión producido al vaciarse el agua que llena las células.
En los equisetos actuales, del esporangio abierto salen nu-
merosas esporas verdes, provistas de una membrana de es-
tructura particular. Sobre la membrana propiamente dicha
de la espora, compuesta de exosporio y endosporio, ha de-
positado el periplasmodio un perisporio pluriestratificado,
cuya capa exterior está constituida por dos bandas o cintas
estrechas (hapterios; fig. 11-141 H, J), arrolladas en hélice
en disposición paralela y con los extremos conformados a
modo de espátula. Al secarse las esporas, los hapterios pier-
den su arrollamiento y se extienden, pero quedan unidos
entre sí y con el exosporio por un punto en su mitad (J) y
vuelven de nuevo a plegarse al humedecerse el ambiente
(v. 8.4); sirven, por sus movimientos higroscópicos, no sólo
para diseminar las esporas, sino para reunirlas en grupos; en
correspondencia, los gametófitos muchas veces crecen jun-
tos, en grupos densos. En todos los casos, las esporas sólo
mantienen la capacidad de germinar durante algunos días.
Las esporas, todas de la misma forma y constitución, pro-
ducen, cuando germinan, protalos talosos verdes muy di-
vididos (fig. 11-141 A).
Los protalos son lobulados y crespos, bastante ramificados y dor- siventrales; pueden ser monoicos o dioicos. La determinación del sexo de los protalos, potencialmente bisexuales, se produce feno- típicamente, gracias a factores externos. En condiciones de caren-
cia se forman sobre todo gametófitos d. La madurez sexual se al-
canza a las 3-5 semanas; el desarrollo es tan rápido que así se termina pronto la fase del gametófito, muy sensible en relación con la economía hídrica y frente a la competencia de los musgos.
Los gametófitos d, a diferencia de los<;;?, están muy pigmentados
por carotenoides, particularidad que también se observa en mus-
gos (pared del anteridio) y hongos (gámetas cJ de Allomyces) y se
interpreta como protección contra las radiaciones mutágenas. Los anteridios están hundidos en el protalo y los arquegonios so- bresalen de su superficie. Los espermatozoides están arrollados en hélice y poseen numerosos flagelos (fig. 11-141 B ); se originan de 250 a 1000 por anteridio. En la división del zigoto, la primera tabicación (tabique basal, 1- en la fig. 11-141 C) da lugar a la formación de dos células que, a diferencia de Lycopodium (fig. 11-135 D), participan ambas, des- pués de nuevas divisiones (cuadrantes, octantes), en la formación del embrión; no se desarrolla ningún suspensor. Las primeras ho- jas se disponen rápidamente en el polo apical formando un verti- cilo y circundando en forma de anillo el extremo del brote, que continúa su crecimiento por medio de una célula apical trígona (fig. 3-2 A). El primordio de la primera raíz se halla situado late- ralmente respecto al eje longitudinal (fig. 11-141 C) y atraviesa el protalo hacia abajo (fig. 11-141 D).
La mayoría de las especies del género Equisetum, difun-
dido desde los países tropicales hasta las zonas frías, pre-
fieren los ambientes húmedos. Mientras las especies euro-
peas sólo llegan a 2 m de altura (E. telmateia), algunos
representantes tropicales del género (p. ej., E. giganteum,
de América del Sur), de porte trepador, pueden alcanzar
12 m de longitud.
En Equisetum arvense, la cola de caballo o equiseto menor
(fig. 11-141), así como en otras especies que reducen tam-
bién sus partes epigeas en invierno, las cortas ramas latera-
les del rizoma se convierten en tubérculos invernantes re-
dondeados que almacenan sustancias de reserva; sin
embargo, hay también especies sempervirentes (p. ej., E.
hyemale).
En ciertas especies de equisetos, una parte de los tallos
permanece estéril y se ramifica en abundancia, mientras
que otros llevan en sus extremos las «flores» y se ramifi-
Clase: Psilotopsida 733
A B e
Fig. 11-143: Equisetales. A Archaeocalamitaceae, Archaeoca/amites radia tus ( 1 /3x). B-D Calamitaceae. B Annu/aria stel/ata ( 1 /2x). C Calamostachys
binneyana, estróbilo de esporangios en sección longitudinal, con hojas estériles (4x). D Calamostachys casheana, sección tangencial pulimentada de
un esporangióforo, que lleva tres megasporangios y un microsporangio (22x). -A, B: según D. Stur; C: según M. Hirmer, modif.; D: según W.C. William-
son y D.H. Scott.
can más tarde y con menos abundancia en brotes laterales
estériles, o no se ramifican en absoluto (fig. 11-141 E, K).
Las dos familias siguientes están completamente extinguidas. Las especies de las Archaeocalamitaceae, que sólo vivieron en el Carbonífero inferior, tenían hojas dicótomas (fig. 11-143 A), dis- puestas en verticilos superpuestos que se correspondían con los haces conductores que atravesaban los nudos de modo rectilíneo.
Las Calamitaceae se distinguen de las equisetáceas por las siguien- tes características: en los tallos reproductores alternaban verticilos de esporangióforos peltados y verticilos de brácteas lanceoladas (fig. 11-143 C). Junto a las especies isospóreas había una heteros- pórea (D). Las esporas carecían de hapterios. El género Calamites (fig. 11-142 C), muy difundido durante el Carbonífero superior y el Pérmico, era uno de los principales componentes de los bosques que han originado la hulla, en los que convivía con los lepidodendros y las sigilarias. Algunas de sus especies tenían hasta 30 m de altura y, debido a la intensa formación de leño secundario, alcanzaban hasta 1 m de grosor (C, D), pero su tronco presentaba una gran cavidad central, como la de los equisetos (árboles tubulosos). Los tallos, en la mayoría de las especies, se ramificaban en verticilo, pero en al- gunas permanecían simples. Los haces conductores se bifurcaban (igual que en Equisetum) en el extremo superior del entrenudo; las ramas originadas por los haces contiguos se confundían a pares en el entrenudo inmediatamente superior, para formar un haz, mientras que un tercer cordón se apartaba hacia el exterior como rastro foliar (E). Había «conductos infranodales» de curso radial, que se forma- ban por destrucción de células de membrana fina y debían de servir para la aireación. Las hojas (fig. 11-143 B) eran sencillas, uninervias
y lanceoladas; en su ápice había un hidatodo, igual que en los dien- tes de la vaina foliar de los equisetos actuales. En relación con la al- ternancia de los haces conductores primarios, en los entrenudos su- cesivos, los verticilos de hojas alternaban también entre sí.
La clase de los equisetópsidos alcanzó su mayor desarrollo
en el Paleozoico; hoy se hallan todos extinguidos, salvo el
género Equisetum (v. fig. 11-163). Y aun éste sólo represen-
ta en la actualidad un resto de lo que fue en otro tiempo, en
que alcanzó un desarrollo más considerable; en el Mesozoi-
co existieron todavía Equisetites arbóreos con engrosamien-
to secundario. Así pues, nuestros equisetos actuales son sólo
reliquias que, de todos modos, no se pueden considerar di-
rectamente derivadas de los representantes heterospóreos
del Paleozoico, pues la heterosporia puede proceder de la
isosporia, pero no a la inversa. Los equisetos residentes de-
ben de proceder, por consiguiente, de antiguas formas aún
isospóreas. Algunas de las formas extinguidas (Calamites,
Sphenophyllum) eran ciertamente heterospóreas; los equise-
tópsidos han llegado, como llegaron las lepidospermas, has-
ta la formación de semillas en el género Calamocarpon.
Clase 4: Psilotopsida
Con sus brotes bifurcados y por la carencia de raíces, las
especies de Psilotum tienen un cierto parecido externo con
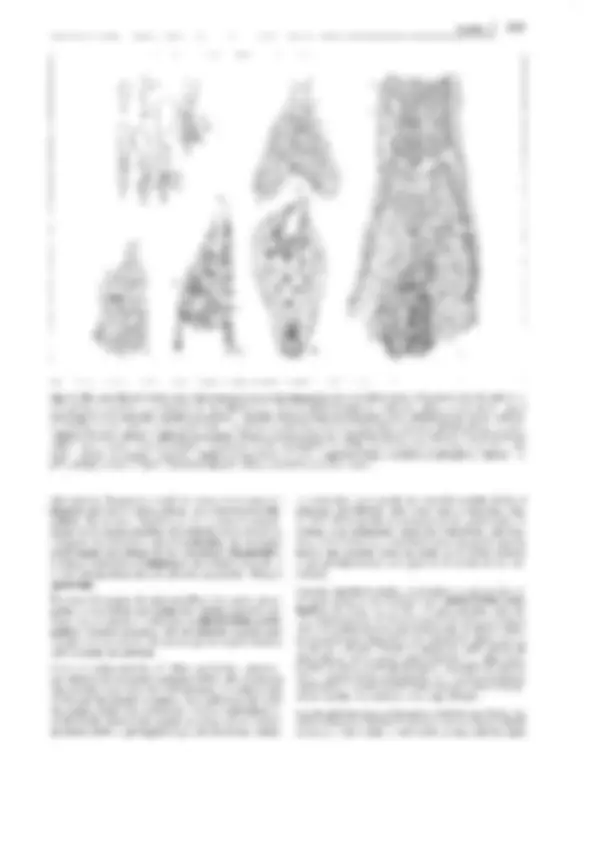
Nivel de desarrollo: Primofilices (= Protopteridiidae) 735
K
L
A B e^ D
## M ## E F G H Fig. 11-145: Pteridopsida. A-D Paso del teloma fértil al esporofilo y E-H de los esporangios al envés de la hoja. J Esporofilo de _Acrangiophyllum_ ### (vegetal semejante a un helecho y de posición sistemática desconocida), del Carbonífero superior (7x). K-M Nerviación de plantas semejantes a hele- ### chos. K Flabelada (Archaeopteris, Devónico superior). L Pinnada (Alethopteris, un representante de las pteridospermas del Carbonífero superior). M Re- ### ticulada (Linopteris, Carbonífero superior) (12x). -A-H: según W. Zimmermann; J: según S. Mamay; K-M: según A.C. Seward, W. Gothan. ## bién el nombre de «frondes». Los esporangios, origina- ## riamente terminales, en las formas derivadas se sitúan en ## el borde o el envés de la hoja. Tallos, raíces y hojas suelen ## crecer por medio de células apicales -como en la clase an- ## terior de los equisetópsidos- no con grupos de iniciales ## como en los licopodiópsidos. ## Los grandes, generalmente peciolados y provistos de nerva- ## dura abundante, están arrollados en su ápice cuando son jó- ## venes (excepto en las ofioglosales). El arrollamiento se ## produce porque el envés, abaxial, de los jóvenes primordios ## (de la hoja o del teloma) crece con más rapidez; posterior- ## mente se iguala. Así pues, las hojas, que suelen desarrollar- ## se acroplásticamente (v. 4.3.1.2), presentan, sobre todo en el ## envés, numerosos esporangios con frecuencia agrupados ## (soros). El tallo, en general, presenta ramificación nula o es- ## casa. Se puede imaginar que la formación del limbo foliar a ## partir de sistemas de telomas y el paso de los esporangios a ## la cara inferior de la hoja en virtud de un mayor desarrollo ## de la superior han tenido lugar tal como indica la fig. 11- ## A-H. En J se reproduce un fósil que representa una fase in- ## termedia del proceso. También las grandes frondes lacinia- ## das pueden haberse originado de manera semejante. Las ## Primofílices, extinguidas, y los grupos actuales de las Eus- ## porangiatae, Leptosporangiatae e Hydropterides presen- ## tan ciertamente relaciones de parentesco, pero no constitu- ## yen, al parecer, comunidades naturales de descendencia, ## sino que son expresión del desarrollo de distintos caracteres. ## Los antepasados de los pteridópsidos hay que buscarlos en- ## tre los psilofitópsidos. Las primofílices representan el nexo ## de unión entre los dos grupos; por un lado muestran una ## cierta afinidad con los psilófitos, pero, por otro, presentan, ## a la vez, caracteres de nivel muy superior. Es común a todas ## las primofílices la posesión de esporangios terminales ## (fig. 11-146 B), así como la particularidad de que los seg- ## mentos foliares todavía no se hallan situados en un mismo ## plano ( «frondes tridimensionales»). La transición que ## conduce de los psilófitos a las primofílices es tan gradual ## que resulta posible dudar respecto a si algunas formas (Pro- ## topteridium y Pseudosporochnus; fig. 11-146 A-E) deben ## situarse aún en los psilófitos o ya entre los helechos. La dis- ## tribución de las primofílices en el tiempo se corresponde ## también con su posición filogenética; aparecen en el Devó- ## nico medio y se extinguen en el Pérmico inferior. ## Dentro de las primofílices se realiza la transformación de ## las ramas bifurcadas dispuestas en fascículos con espo- ## rangios terminales (p. ej., Pseudosporochnus; fig. 11- ## 146 A, B) en hojitas claviformes y planas, con ramifi- ## cación dicótoma irregular y esporofilos con esporangios ## marginales formados de la misma manera (p. ej., Cla- ## doxylon; fig. 11-146 M). Los esporofilos estaban, en par- ## te, entremezclados con segmentos foliares y, en parte, uni- ## dos formando grupos, aunque sin formar todavía «flores». ## El aumento de concrescencias laterales de los telomas ## produjo hojas de gran superficie y de nerviación dicó- ## toma, que ya existían en el Devónico superior (fig. 11- ## 145; v. también entre las leptosporangiadas recientes ## Adiantum, fig. 4-60 A). ### A partir de la nerviación dicótoma se desarrolla progresivamente la ### nerviación reticulada en helechos y pteridospermas. En el Devó- ### nico superior sólo existía nerviación flabelada con ramificación di- ### cótoma de los nervios, en el Carbonífero inferior se presenta por pri- ### mera vez la nerviación pinnada y en el Carbonífero superior, la ### reticulada, que determina un aprovisionamiento más completo de la ### hoja en agua y sustancias nutritivas (fig. 11-145 K-M). ### Un requisito previo a la constitución de estas formas foliares la- ### minares es que los telomas se dispongan en un solo plano. En las ### formas primitivas se hallaban a veces todavía perpendiculares en- ### tre sí (como sucede ahora en las ofioglosales ); también podía no ### haberse producido aún la planación de los segmentos, de modo ### que las «hojas» podían ser aún cilíndricas. Los dos hechos se dan ### en Stauropteris (fig. 11-146 G, Carbonífero superior); sin embar- ### go, en tales telomas foliares cilíndricos podía presentarse ya pa- ### rénquima en empalizada. **736 11 Sistemática y filogenia** _¡_ _:~_ 50 ### A \ \ \. /; \~ ¡ J _if 1_ 1 lJ G ## D ### Fig. 11-146: Primofilices. A-8 Pseudosporochnales. A Pseudosporochnus, reconstrucción, B extremo de una ramificación (tam. nat.). C-F Protop- ### teridiales, C-E Protopteridium hostimense, Devónico, C fronde (1/4x), D pinna estéril y E fértil (3x); F Pertica quadrifaria, Devónico, rama con esporan- ### gios (s). G-H Coenopteridales, Stauropterís o/dhamía. Carbonífero, G sección de una fronde estéril, reconstrucción (tam. nat.), H ídem, esporangio con ### punto de dehiscencia (35x). J-M Cladoxylales, Cladoxylon scoparium, Devónico medio, J porción de una rama (2/3x), K folíolo (2x), L sección trans- ### versal a través de la plectostela (4x), M grupo de esporangios (2x). - A: según W. Zimmermann; B: según S. Leclercq y P.H. Banks; C-E y J-M: según ### R. Krausel y H. Weyland; F: según Kasper y H.N. Andrews; G: según M. Chaphekar, modif. por K. Magdefrau; H: según D.H. Scott. s esporangio.