¡Descarga La Membrana Plasmática: Definición, Estructura y Función - Prof. Ochando y más Apuntes en PDF de Citología solo en Docsity!
La membrana plasmática
TABLA DE CONTENIDOS
- 1 Definición
- 2 Estructura
- 3 Función de la membrana plasmática
- 4 Transporte específico de moléculas hacia el interior o hacia el exterior celular
- 5 Diferentes tipos de proteínas transportadoras de membrana
- 6 Ensamblaje de una membrana ⚬ 6.1 Estructura química de un fosfolípido ⚬ 6.2 Ensamblajes de fosfolípidos anfipáticos en soluciones acuosas ▪ 6.2.1 Micela ▪ 6.2.2 Bicapa ▪ 6.2.3 Liposoma ▪ 6.2.4 Bicapa lipídica
- 7 Asimetría en la membrana
- 8 Funciones de la membrana plasmática ⚬ 8.1 Transporte ▪ 8.1.1 Proteínas transportadoras de membrana ⚬ 8.2 Señalización molecular ⚬ 8.3 Reconocimiento célula-célula
- 9 Referencias
DEFINICIÓN
Todas las células están rodeadas y separadas de su ambiente exterior por una membrana con una permeabilidad selectiva llamada membrana plasmática, que define los límites de la célula, su perímetro celular, actuando como una barrera que separa su contenido interno (el citoplasma y núcleo) del medio externo.
El modelo actual que describe la organización estructural de la membrana plasmática fue propuesto en1972 por los científicos Garth Nicholson y S. Jonathan Singer 1. Este modelo describe la membrana plasmática como un mosaico fluido que contiene diversas proteínas embebidas en una matriz de fosfolípidos.
Los fosfolípidos en la membrana forman una bicapa lipídica con las cabezas polares dirigidas hacia el exterior y las colas hidrofóbicas hacia el interior de la bicapa. Esta tiene un grosor aproximado de 75 Å (ángstrom) o 7.5 nanómetros (nm), por ello no es posible visualizarla al microscopio óptico pero sí con el microscopio electrónico, ya que éste ofrece imágenes de la membrana plasmática en la que se pueden observar dos líneas oscuras laterales y una central más clara, imagen que recuerda a las vías del tren.
ESTRUCTURA
El modelo de mosaico fluido es el que describe la estructura de capas de La membrana plasmática| membrana plasmática.
La membrana plasmática es un mosaico de diferentes tipos de proteínas (generalmente glicoproteínas) embebidas en una bicapa de fosfolípidos. El conjunto se mueve en el plano de la membrana como si fuera un fluido, de ahí el nombre que recibe este modelo de estructura: mosaico fluido.
Así pues, todas las membranas biológicas son entidades dinámicas, estructuras fluidas, pues la mayoría de suslípidos y proteínas son capaces de moverse en el plano de la membrana, además de sufrir un continuo recambio de componentes. Al igual que un mosaico, la membrana plasmática es una estructura compleja construida de diferentes elementos, proteínas, fosfolípidos y esteroides. Cuatro fosfolípidos, tres fosfoglicéridos principales fosfatidilcolina, fosfatidiletalonamina , y fosfatidilserina y la esfingomielina constituyen más del 50 % de los fosfolípidos de la mayoría de las biomembranas.
Los esteroides, como el colesterol , tienen un importante papel en la regulación de las propiedades físico-químicas de la membrana biológicas regulando su resistencia y fluidez. La cantidad relativa de estos componentes varían de membrana en membrana, y los tipos de lípidos en la membrana también pueden variar.
Las proteínas integrales (llamadas también proteínas transmembrana ) cruzan completamente la bicapa lipídica. Las proteínas periféricas se asocian con la membrana principalmente a través de interacciones no covalentes específicas con las proteínas integrales o lípidos de membrana. Las cadenas de oligosacáridos se encuentran unidos covalentemente a muchas proteínas extracelulares y al dominio exoplasmático de muchas proteínas transmembrana.
Permeabilidad de una bicapa de fosfolípidos a varias tipos de moléculas
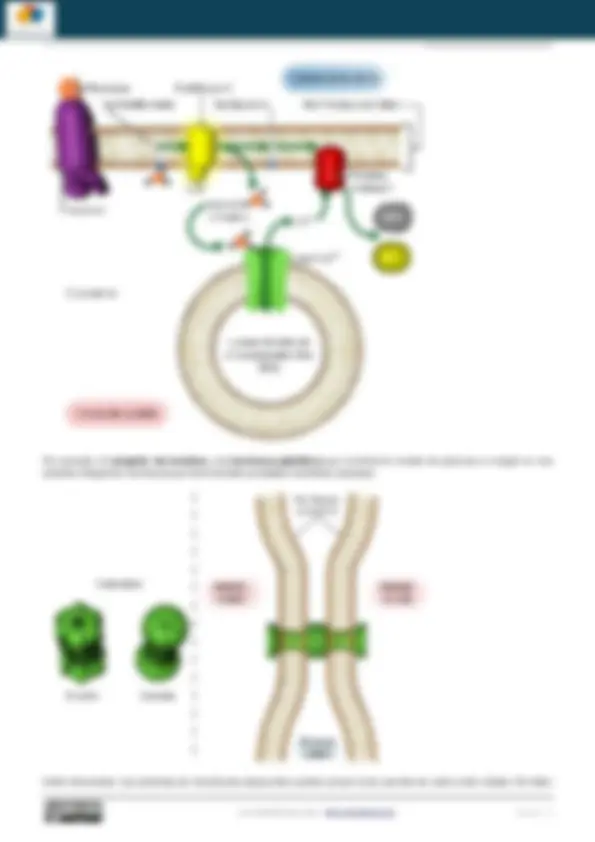
Las proteínas embebidas en la membrana realizan varios tipos de funciones, algunas de las cuales están relacionadas con el mantenimiento de la hemostasia celular (e.g. transporte, enzimas) y otras que son básicas para integrar una célula en un tejido (receptores, funciones de anclaje, de conexión y reconocimiento intercelular).
Diferentes tipos de función adscritas a distintos tipos de proteínas de membrana transmembranales: transporte, de
anclaje y conexión, receptores y enzimas. Una misma proteína transmembrana puede más de una función por ejemplo ser un receptor y tener actividad enzimática.
TRANSPORTE ESPECÍFICO DE MOLÉCULAS HACIA EL
INTERIOR O HACIA EL EXTERIOR CELULAR
Esta función de las proteínas de membrana es de vital importancia para la toma de nutriente por la célula, la salida de productos de desecho de la célula; así como el mantenimiento de diferentes tipos de gradientes electroquímicos (e.g. potencial de membrana) y de concentración de diferentes moléculas a través de la membrana necesarios para mantener la vida celular.
Tipos de transporte: pasivo y activo.
Diferentes tipos de movimiento de las moléculas a través de las membranas biológicas. Difusión pasiva y la difusión facilitada no requieren el consumo de energía, ya que se realiza a favor de gradiente de concentración o electroquímico; cuando las sustancias están cargadas la dirección y magnitud del flujo de iones a través de una membrana depende tanto de la diferencia de concentración y de la diferencia eléctrica a través de ella, estas dos fuerzas son por ello colectivamente conocidas como gradiente electroquímico. Algunas sustancias entran directamente en la célula a través de difusión pasiva pero muchas sustancias de interés para la célula atraviesan la membrana mediante difusión facilitada. El transporte activo se realiza con consumo de energía (acoplando a la hidrólisis ATP) al realizarse enana dirección energéticamente desfavorable contra un gradiente electroquímico o de concentración.
DIFERENTES TIPOS DE PROTEÍNAS TRANSPORTADORAS
DE MEMBRANA
Se suelen distinguir dos clases de proteínas que intervienen en la difusión facilitada: Las proteínas canales o de canal facilitan la difusión formando poros (e.g porinas) abiertos en la membrana que permiten la libre difusión de cualquier molécula de tamaño y carga apropiados. Las proteínas transportadoras se unen específicamente en un lado de la
Estructura de la fosfatidilcolina unos de los fosfolípidos (fosfoglicerido) más comunes en las membranas biológicas. Estructura química y representación de modelos de bolas de su estructura atómica, que da una idea aproximada de la forma de la molécula.
Ensamblajes de fosfolípidos anfipáticos en soluciones acuosas
MICELA
Unidades con forma de cuña (cabeza más grande que la cadena). En lasmicelas, las cadenas hidrofóbicas de los ácidos grasos están secuestradas en el núcleo de la esfera. No hay agua en el interior hidrofóbico.
BICAPA
Unidades con forma de cilindro (cabeza de igual tamaño que la cadena). En una bicapa abierta todas las cadenas hidrofóbicas están protegidas del agua, excepto las situadas en los bordes. Representación abreviada típica de la estructura anfipática de los fosfolípidos, con una cabeza polar hidrofílica y una cola apolar hidrofóbica
LIPOSOMA
Estructuras esféricas huecas con una pared en forma de bicapa lipídica y una cavidad interior acuosa
BICAPA LIPÍDICA
Las proteínas integrales de membrana (PIM) tienen diferentes modos de anclarse a la membrana plasmática : a través de uno o varios segmentos - helicoidales hidrofóbicos con distinta orientación topológica [amino N- Carboxilo terminal] ( a,b,c ). Existen también PIM que se anclas por medio de láminas (e.g. porinas , proteínas con estructura en forma de barril construido de 8 a 22 láminas que forman un poro, presentes en la membrana exterior de las bacterias Gram negativas y en la membrana exterior de las mitocondrias). Por otra parte, existen proteínas que se encuentran ancladas exclusivamente a una de las hojas de la bicapa (monocapa) por un larga cadena lipídica hidrofóbica de diferente composición de ácido graso (e.g. ácido mirístico, ácido palmítico) o de grupos prenilo ( d ). Otras proteínas se anclan exclusivamente a la monocapa exterior de la memebrana plasmática través de un anclaje de glicosilfosfatidilinositol (un glicolípido abreviadamente GPI), llamadas por ello proteínas GPI ( e ).
El colesterol se inserta dentro de la bicapa de fosfolípidos con sus grupos polares hidrófilo (-OH) próximos a las cabezas de los fosfolípidos.
FUNCIONES DE LA MEMBRANA PLASMÁTICA
La membrana plasmática, tiene un doble papel fisiológico en la célula , por una parte aísla y por lo tanto diferencia el medio interno celular del ambiente exterior pero a su vez media la interacción entre la célula y su entorno exterior al permitir intercambio selectivo de materia y energía e información (diferentes tipos de señales físicas y químicas) entre ambos, intercambio que es necesario para mantener una adecuada homeostasis del medio interno, clave en el mantenimiento de la vida celular. Esta doble función de la membrana plasmática es posible por una parte gracias a la naturaleza aislante que en medio acuoso proporciona la bicapa lipídica hidrofóbica y por otra en las funciones de transporte que desempeñan las proteínas embebidas en la membrana. Es la actividad específica transportadora de dichas proteínas la que determina permeabilidad selectiva de las biomembranas y de ese modo desempeñan un papel crucial en la función de la membrana.
La membrana plasmática es una barrera selectivamente permeable que permite el paso de unas sustancias pero no de otras. Determina pues que sustancias entran o salen de la célula. El interior hidrofóbico de la bicapa de fosfolípidos es una de las razones por las que la membrana es selectivamente permeable. Así, la bicapa lipídica tiene un papel fisicoquímico dual pues sirve por una parte como un solvente de las proteínas de la membrana y por otra actúa como una barrera a la permeabilidad.
Mientras que las moléculas hidrofóbicas, que son solubles en lípidos (e.g etanol) pueden pasar fácilmente la membrana, moléculas pequeñas como el oxígeno (O2), dióxido de carbono (CO2), Nitrógeno (N2) pueden difundir entre los fosfolípidos de membrana, pero moléculas hidrofílicas pequeñas como agua, nutrientes como la glucosa e iónes [Na+, K+, protones (H+), etc?] no pueden pasar directamente a través de los fosfolípidos de la membrana plasmática. Estos compuestos deben pasar a través de proteínas de transporte específico situadas en la membrana.
Las proteínas embebidas en la membrana realizan varios tipos de funciones algunas de las cuales están relacionadas con el mantenimiento de la hemostasia celular (e.g. transporte, enzimas) y otras que son básicas para integrar una célula en un tejido (receptores, funciones de anclaje, de conexión y reconocimiento intercelular)
Transporte
Transporte específico de moléculas hacia el interior o hacia el exterior celular. Esta función de las proteínas de membrana es de vital importancia para la toma de nutriente por la célula, la salida de productos de desecho de la célula; así como para el mantenimiento de diferentes tipos de gradientes electroquímicos (e.g. potencial de membrana) y de concentración de diferentes moléculas a través de la membrana necesarios para mantener la vida celular.
Diferentes tipos de movimiento de las moléculas a través de las membranas biológicas. Difusión pasiva y la difusión facilitada no requieren el consumo de energía , ya que se realiza a favor de gradiente de concentración o electroquímico; cuando las sustancias están cargadas la dirección y magnitud del flujo de iones a través de una membrana depende tanto de la diferencia de concentración y de la diferencia eléctrica a través de ella, estas dos fuerzas son por ello colectivamente conocidas como gradiente electroquímico. Algunas sustancias entran directamente en la célula a través de difusión pasiva pero muchas sustancias de interés para la célula atraviesan la membrana mediante difusión facilitada. El transporte activo se realiza con consumo de energía (acoplando a la hidrólisis ATP) al realizarse en una dirección energéticamente desfavorable contra un gradiente electroquímico o de concentración.
PROTEÍNAS TRANSPORTADORAS DE MEMBRANA
Se suelen distinguir dos clases de proteínas que intervienen en la difusión facilitada: Las proteínas canales o de canal facilitan la difusión formando poros (e.g porinas) abiertos en la membrana que permiten la libre difusión de cualquier molécula de tamaño y carga apropiados. Las proteínas transportadoras se unen específicamente en un lado de la membrana a las moléculas que van a ser transportadas, sufren entonces un cambio conformacional que permite que la molécula pase a través de la membrana y sea finalmente liberada al otro lado. Se distinguen tres tipos de transportadores, Uniportes , transportan un solo tipo de molécula a favor de gradiente de concentración y proteínas de cotransporte ( Simportes y Antiportes ), que median el movimiento de una molécula en contra de su gradiente de concentración dirigido por el cotransporte (en la misma dirección o en sentidos opuestos respectivamente) de otra molécula o ión a favor de gradiente. Existen también canales iónicos (que transportan iones, moléculas cargadas) cuya apertura es regulada por la unión de un pequeño ligando (e.g. un neurotransmisor, acetilcolina) o con una apertura regulada por cambios en el voltaje eléctrico (canales iónicos regulados por voltaje para cationes Na+, K+, Ca2+, etc?).
Por otra parte, las bombas potenciadas por ATP o bombas ATPasas como también se las denomina, que permiten a la célula el transporte activo de ciertas moléculas en contra de gradiente de concentración o electroquímico. Se han descrito diferentes tipos de bombas ATPasas que median el transporte activo dependiente de la hidrólisis de ATP. La familia de transportadores de tipo ABC (acrónimo de ATP-binding Cassete en inglés ) es una de la más numerosa.
Por ejemplo, el receptor de insulina , una hormona péptidica que controla los niveles de glucosa en sangre es una proteína integral de membrana que tiene también actividad enzimática (quinasa).
Unión intercelular. Las proteínas de membranas adyacentes pueden actuar como puentes de unión entre células. Permiten
la comunicación intercelular. Las uniones comunicantes ( gap junctions en inglés) un ejemplo de estructuras para la comunicación intercelular construidas con proteínas integrales de membrana llamadas conexinas.
Reconocimiento célula-célula
Los oligosacáridos que se encuentra unidos covalentemente a glicoproteínas y glicolípidos de la membrana plasmática se dirigen hacia el espacio extracelular formando el glicocalix , una especie de manto de carbohidratos que recubre la superficie celular. El glicocalix sirve para proteger la superficie celular de diferentes tipos de agresiones mecánica y químicas, además de estar implicado en diferentes tipos de interacción célula-célula al servir como puntos de reconocimiento (marcadores) de la superficie celular que son identificados por receptores específicos encontrados en otras células (reconocimiento intercelular). Por ejemplo los grupos sanguíneos humanos A, B, AB y O reflejan variaciones en la estructura de los oligosacáridos unidos a proteínas o lípidos en la superficie de los eritrocitos o glóbulos rojos [[1]].
Los distintos grupos sanguíneos humanos son una consecuencia de la diferente estructura de los oligosacarídos unidos covalentemente a lípidos y proteínas de la membrana de los eritrocitos o glóbulos rojos. Los glóbulos blancos (leucocitos,e.g neutrófilos, macrófagos) son capaces de adherirse a las células endoteliales gracias a los restos de oligosacarios que expone en su superficie y que son reconocidos por receptores específicos (selectinas) de la superficie de las células endoteliales.
.- Proteínas receptoras, forman enlaces estructurales entre las proteínas del citoesqueleto celular y la matriz extracelular. De importancia fundamental en la construcción de tejidos y en el movimiento celular.