¡Descarga Membrana celular: funciones y composición de proteínas y más Apuntes en PDF de Biología Celular solo en Docsity!
TEMA 1 ORGANITZACIÓ CEL·LULAR: EUCARIOTES I PROCARIOTES
Cèl·lules procariotes (primeres en originar-se evolutivament parlant) La diversitat bioquímica els permet viure i adaptar-se a la major part dels medis terrestres. Inclou 2 tipus d’organismes: eubacteris i arqueobacteris Estructura: Paret cel·lular: molt comuna per tal de protegir l’interior cel·lular d’agents vírics, patògens... Consten de dues membranes: una externa més porosa (pròpia dels GRAM -) i la plasmàtica. No tenen orgànuls ni nucli per separar físicament el material genètic d’altres substàncies presents al citosol de la cèl·lula. Cèl·lules eucariotes Inclouen: protists, fongs, animal i vegetals Es componen d’un nombre important d’orgànuls i complexos, la majoria separats per membranes internes, que compartimenten i organitzen les diverses funcions que es duen a terme a la cèl·lula. Estructura general: Nucli Reticle endoplasmàtic Aparell de Golgi Mitocondris Cèl·lula animal i cèl·lula vegetal: orgànuls diferencials Orgànuls exclusius de la cèl·lula animal Orgànuls exclusius cèl·lula vegetal Lisosomes Paret cel·lular Microvil·lis Vacúols Cloroplasts
Principals diferències en l’organització cel·lulars entre procariotes i eucariotes.
Fosfolípids: més del 50% de lípids que formen la membrana. N’hi ha dos tipus: fosfoglicèrids (derivat del glicerol) i esfingolípids (derivat de l’esfingosina). Fosfoglicèrids: les cues son àcids grassos de 16-18 C. Una és recta (cua saturada) i l’altre torçada degut a un doble enllaç (cua insaturada). Les cues deriven del glicerol, que ja forma part del cap de la molècula. El cap també està format per un fosfat amb un grup alcohol unit (colina, serina, etanolamina o inositol). El nom de cada fosfoglicèrid indica l’alcohol que forma part del cap polar. SINTETIZATS AL RE Esfingolípids: deriven de l’esfingosina. No són gaire abundants (un únic tipus a la membrana: esfingomielina). Les dues cues son saturades. SINTETITZATS A L’APARELL DE GOLGI Glucolípids: molt menys abundants. Només presents a la monocapa externa i deriven dels esfingolípids: glucoesfingolípids. Hi ha dos tipus: cerebròsids (només tenen un residu de sucre) i gangliòsids (múltiples residus de sucre i un d’aquests residus és el NANA o àcid siàlic i té càrrega negativa). Propis dels microdominis lipídics. Esteroides: només en eucariotes. En cèl·lules animals és el colesterol i en vegetals els fitoesteroides. Donen rigidesa a la membrana i interaccionen amb els caps polars dels fosfoglicèrids. Microdominis lipídics: regions de la membrana riques en esfingolípids, glucolípids, colesterol i fosfoglicèrids amb cues saturades (són més llargs que els fosfoglicèrids). És una zona més gruixuda i menys fluïda de la membrana. Les quantitats elevades de colesterol fan aquesta zona sigui molt més rígida i compacte que la resta de la cèl·lula. També s’anomenen balses lipídiques perquè floten per tota la membrana com a conjunt a través de les altres estructures que formen la membrana. S’hi acumulen proteïnes transmembranals o integrals. La seva funció principal és formar compartiments
per diferents proteïnes i per tant dur a terme diverses funcions (p.e. senyalització i formació de vesícules). Proteïnes: Responsables de les funcions específiques de la membrana del tipus de cèl·lula qualsevol. Son un 50% en massa, però es calcula que hi ha una proteïna per cada 50 molècules de lipídiques. Un gran nombre són proteïnes amfipàtiques. La gran majoria porten oligosacàrids, és a dir, són glucoproteïnes. En diferenciem 3 tipus: integrals, perifèriques i ancorades a un lípid. Proteïnes integrals de membrana: son amfipàtiques i els dominis hidrofòbics es troben a l’interior de la bicapa. Es tracta de les proteïnes més abundants en la membrana. SINTETITZADES AL RE. Proteïnes transmembranals d’un sol pas: un sol domini ancorat a la membrana. Proteïnes transmembranals multipas: més d’un domini transmembranal. Proteïnes integrals monotròpiques: són aquelles proteïnes inserides en només una de les bicapes (la interna principalment). Proteïnes perifèriques: són hidrofíliques i estan ancorades a la membrana gràcies a interaccions electrostàtiques no covalents, normalment unides a proteïnes integrals de la membrana. Són més fàcils d’aïllar de la membrana, canviant la composició iònica del medi. Segons on es produeixi la síntesi d’aquestes proteïnes, es poden trobar a la zona del citosol (fetes al citosol mateix) o a la zona extracel·lular (fetes al RE). Proteïnes ancorades per lípids: hidrofíliques i unides covalentment a un lípid o glucolípid inserit a la bicapa. Les proteïnes que es troben al costat citosòlic estan sintetitzades al citosol i ancorades a un lípid (normalment a un àcids gras o grup prenil). Les proteïnes del costat extracel·lular estan sintetitzades al RE i ancorades per un glucolípid: GPI = glucosilfosfatidilinositol. Grup pernil GPI Oligosacàrids units a proteïnes: per formar glucoproteïnes. Dos tipus: N-units (aminoàcid Asn, que conté un grup amino, es la molècula d’unió entre el sucre i la proteïna) i O-units (l’aminoàcid d’unió es la Ser o Thr, que conté un O). En una sola proteïna podem trobar diversos oligosacàrids amb diferents formes d’unió. Plegament de les proteïnes: Hèlix alfa Làmina beta (poc comuna en cèl·lules animals, proteïnes barril)
TEMA 3 TRANSPORT DE MOLÈCULES A TRAVÉS DE LA MEMBRANA
La membrana és una estructura impermeable que permet protegir l’interior cèl·lula. Per tal que la cèl·lula pugui funcionar bé necessita certs ions, petites molècules o partícules. El transport d’aquestes molècules es fa a través de proteïnes de transport i altres mecanismes a la membrana. Difusió simple: Es produeix a favor de gradient electroquímic (la membrana té un cert potencial proper a -70mV). La membrana és permeable a aquestes petites molècules que són principalment gasos i molècules petites sense càrrega. Aquest mecanisme permet no consumir energia metabòlica. Depenent de les característiques fisicoquímiques de les molècules, la velocitat de difusió serà diferent. És proporcional linealment a la concentració que és necessària a l’interior cel·lular. Osmosi: Es produeix quan hi ha diferents concentracions en un costat i l’altre de solut, el qual no pot travessar la bicapa. L’aigua, que és el dissolvent essencial, és el que es mou a través de la membrana plasmàtica. Es diferencien tres tipus de solucions en què es pot trobar la cèl·lula: Isotòniques: concentracions en equilibri.
Hipertòniques: cal diluir la concentració de l’entorn per tant surt l’aigua de la cèl·lula. La cèl·lula entra en estat de plasmòlisi, és a dir, es desinfla. Hipotòniques: cal diluir la concentració de la cèl·lula respecte l’entorn. La cèl·lula en aquest cas s’infla i es troba en turgència. Aquest estat pot derivar en una mort cel·lular per xoc osmòtic si el volum d’aigua que ha d’entrar és superior al volum de la cèl·lula. Transport passiu: Transport de molècules a favor del gradient i la seva concentració però no pot passar directament a través de la membrana i és per això que s’anomena difusió facilitada, perquè hi participen proteïnes. Les proteïnes que actuen en aquests casos són: les permeases de transport passiu (perquè van a favor del gradient), les proteïnes canal... Proteïnes de canal: no pateixen canvi conformacional, és a dir, poden estar obertes o tancades però no canvien en el moment en què les molècules entren a la proteïna. N’hi ha diferents tipus: unions gap (entre cèl·lula i cèl·lula), porines, aquaporines, canals iònics. La velocitat de transport és molt més elevada que la de les permeases. Aquests canals no estan sempre obert perquè les concentracions no es podrien mantenir per tant es regulen i s’obren a partir de diversos estímuls. Segons la seva regulació s’anomenen: regulats per voltatge (segons la càrrega que produeixen les concentracions d’ions a un costat i l’altre), regulats per lligand (quan una petita molècula s’uneix al canal i l’obre), regulats mecànicament (és poc freqüent p.e. estimulació mecànica per les vibracions del so en les cèl·lules del conducte auditiu). o Canals de Na+: seleccionen l’ió en funció de la seva mida quan estan units (el Na+^ es un ió mes petit que el potassi) 24 dominis transmembranals, tots units. o Canals de K+: seleccionen degut al filtre selectiu. Hi ha àtoms d’oxigen (4) que toquen i interaccionen el K+. S’allibera de la molècula d’aigua i passa just pel canal (el sodi no arriba a interactuar amb els oxígens i segueix unit a l’aigua). 24 dominis transmembranals en 4 subunitats diferents. o Canals iònics del K+^ en les cèl·lules animals (canals de fuga de potassi): cap a l’exterior sempre, ja que la concentració dins de la cèl·lula és major degut a les bombes de Na+/K+. Permeases: són proteïnes altament específiques (transporten només un o dos tipus de molècules). Es localitzen també a la membrana d’orgànuls. Permeten transportar a alta velocitat moltes molècules. Segueixen la cinètica enzimàtica, és a dir, el creixement de la concentració pot augmentar, però la velocitat es mantindrà constant. La seva estructura principal és de 12 dominis transmembranals en hèlix alfa i una sèrie de dominis hidrofílics que queden exposats al citosol o a la matriu, els quals tenen les seqüències específiques d’aminoàcids per poder determinar quines molècules pot transportar la proteïna. Les molècules només van al lloc d’unió d’acord amb el gradient de concentració en cada moment. Aquest transport genera el potencial de membrana: valors negatius a la banda citosòlica i valors positius a la banda extracel·lular. Potencial d’acció (i la sinapsis neuronal): a les cèl·lules nervioses. Cada cèl·lula té una primera fase de potencial d’acció: despolarització (de valors negatius a valors positius). Tot seguit es produeix de repolarització en què es tornen als valors inicials. Aquests canvis són generats per l’acció de diversos canals iònics. Durant la despolarització entren ions sodi massivament. Durant la segona fase l’entrada de sodi s’atura i augmenta el flux de sortida d’ions potassi. En una neurona, comença quan rep un neurotransmissor que s’uneix a diversos canals d’acció de
Exemples de bombes: o Bomba de Na+/K+: és de tipus P, vital per al funcionament de les cèl·lules i es dediquen quasi el 1/3 d’ATPs en aquestes proteïnes. Per cada ATP que hidrolitza, treu 3 Na+^ en contra de gradient i entra 2 K+. Durant la fosforilació i desfosforilació la bomba canvia de conformació. En l’estat E1, els llocs d’unió del sodi són d’alta afinitat i s’uneixen les molècules a la vegada de l’ATP. Es produeix la fosforilació i canvia la conformació, fent que els 2 llocs d’unió del K+^ siguin d’alta afinitat ja que s’ha obert cap a l’exterior de la cèl·lula i entri en estat E2. S’allibera el sodi i es desfosforila de tal forma que es torna a canviar la conformació de la bomba, es torna a l’estat E1. Aquestes bombes col·laboren en un 10% en la creació de potencial de membrana: crea descompensació de càrregues, s’encarrega de mantenir el gradient de potassi dins de la cèl·lula. Estabilitzen la forma i el volum cel·lular, contrarestant el fet que les macromolècules atrauen ions i crea medi hipotònic extracel·lular. Per últim, també ajuda a controlar el gradient electroquímic generat a la membrana cel·lular. o Bomba de Ca2+: són bombes de tipus P, i es troben a la membrana plasmàtica i al RE. Expulsen el calci del citosol. Una vegada a entrat calci mitjançant canals iònics, el mecanisme més comú són aquestes bombes. El funcionament és molt similar al de les bombes anteriors: té dos estats E1 i E2, hi ha fosforilació i defosforilació. o Bomba d’H+: es troben en totes les cèl·lules eucariotes (en endosomes, lisosomes i en vacúols a les cèl·lules vegetals). Serveix per mantenir el pH neutre del citosol i acidifiquen l’interior dels orgànuls, emmagatzemant-ne els protons. En cèl·lules tipus P, a la membrana plasmàtica de bacteris, fongs i plantes, fan la mateixa funció que els canals de fuga de potassi. Les cèl·lules secretores d’àcid dels animals en tenen a la part apical de la cèl·lula. o Proteïna MDR1: es una bomba ABC que es troba al fetge, l’intestí i ronyons. S’encarrega d’eliminar toxines. Enlloc de ions, expulsa petites molècules. també consisteix en canvi de conformació degut a la unió d’ATP per alliberar la molècula. No es fosforila la bomba, sinó que el fòsfor queda lliure al citoplasma.
Transport actiu secundari: les proteïnes que formen part d’aquest tipus, son permeases que necessiten una segona molècula (ions) per obtenir energia i fer circular a contra de gradient les molècules principals. Segons quins sigui el sentit de la molècula i el cofactor, s’anomenen antiport o simport. Els ions que es transporten a favor de gradient són: Na+^ en cèl·lules animals i de H+^ en bacteris, fongs i vegetals, i la membrana d’orgànuls. o Simport Na+/glucosa: a l’epiteli intestinal, permet digerir la glucosa i transportar-la fins al torrent sanguini. La permeasa que fa aquest transport (es localitza al domini apical) obté l’energia de transport de la glucosa gràcies a l’entrada del sodi. 2 Na+^ = 2 glucoses. Per tal que surti cap als capil·lars sanguinis, una permeasa de transport passiu portarà la molècula de glucosa a l’exterior, però a favor de gradient (es localitza al la zona basolateral). o Antiport Na+/Ca2+: col·laboren a mantenir la baixa concentració de calci. o Antiport Na/H+: junt amb les bombes de protons ajuden a mantenir la baixa concentració de H+ dins del citosol. (AQUESTES BOMBES DEPENEN DE LA BOMBA NA+/K+. INDIRECTAMENT, NECESSITEN ATP PER FUNCIONAR. EL PRIMARI CONSUMEIX DIRECTAMENT. SENSE EL FUNCIONAMENT DE LES BOMBES DE TRANSPORT PRIMARI, LES PERMEASES NO PODEN PARTICIPAR.)
on les cadenes formen xarxes laminars i a vegades esterificades, envoltant òrgans, etc. o Elastina: proteïna hidrofòbica molt rica en glicina i lisina. La tropoelastina (precursor de l’elastina) es secreta com a proteïna soluble per formar les fibres elàstiques. Normalment està envoltada per fibril·les de col·lagen. Proteïnes adhesives: tenen molts dominis d’unió i les seves funcions principals son organitzar el contingut de la matriu extracel·lular, unir les cèl·lules a la matriu i contribuir a la migració i la forma de les cèl·lules. N’hi ha dues de molt abundants: o Fibronectina: es una glucoproteïna formada per un dímer de dues cadenes similars de 2500 aminoàcids (unió per ponts disulfur). Tenen certs dominis que compleixen algunes funcions d’acord amb la molècula o cèl·lula a la qual estiguin units. S’uneix al col·lagen i s’adhereix a la cèl·lula. Tot i que la fibronectina derivi d’un sol gen, tenen diferents tipus de descodificació. o Laminina: consisteix en un conjunt de tres cadenes polipeptídiques diferents (unió per punts disulfur) que formen una estructura en forma de creu, un braç llarg i tres de curts. Poden formar xarxes i són el component principal de la làmina basal que s’adhereix a les cèl·lules amb integrines. Comunicació cèl·lula-matriu extracel·lular Tal com s’ha dit, la fibronectina i les integrines (proteïna d’unió entre la fibronectina i la cèl·lula) uneixen la matriu amb el citoesquelet. En canvi de conformació d’aquestes proteïnes fa que s’enviïn senyals químics que permeten que els components de la matriu puguin actuar respecte a la situació de les cèl·lules. La resposta son senyals mecànics, o fins i tot la regulació de l’expressió gènica cel·lular. Integrines: es tracta de dues subunitats (alfa i beta) transmembranoses glucoprotèiques. Mentre que la part extracel·lular s’uneix a la fibronectina, la part intracel·lular ho fa amb els filaments d’actina. La cèl·lula també pot influir en els canvis de la matriu mitjançant processos de síntesi, degradació, canvis d’orientació, etc... Malalties relacionades
La paret cel·lular té la funció principal de protegir la cèl·lula i donar-li una estructura i forma adequada. En les cèl·lules vegetals, consta d’un conjunt de fibres de cel·lulosa unides a una matriu que conté proteïnes i polisacàrids. Composició: Cel·lulosa: és un 40% de la massa de la paret. És un polisacàrid format per glucoses enllaçades per enllaços beta(14) formant microfibril·les i fibril·les. Aquesta capa es forma per complexos enzimàtics de la pròpia cèl·lula i la orientació en que es formi indica també la direcció en que creixerà la cèl·lula. Hemicel·lulosa: representa el 20% del pes. Son cadenes lineals de glucosa amb ramificacions de pocs monosacàrids. La seva funció és unir les fibres de cel·lulosa i altres components extracel·lulars. Pectina: és un polisacàrid ramificat que té carrega negativa. Donen resistència a les forces compressives que rep la cèl·lula. és el 30% del pes de la paret. Extensina: (10% del pes) glucoproteïna que aporta resistència i es troba en més quantitat a teixits de suport estructural de la planta. Lignina: polímer d’alcohols aromàtics insolubles que donen rigidesa a la paret de teixits llenyosos. Parts de la paret cel·lular: Paret primària: acumulació de 3 o 4 capes aleatòries de microfibril·les de cel·lulosa. Està en creixement constant i són capaces d’estendre’s. Paret secundària: està tocant a la membrana plasmàtica. Consta de 3 parts (S1, S2, S3). La primera està formada per cel·lulosa, pectina i hemicel·lulosa i les altres dues de cel·lulosa, hemicel·lulosa i lignina. Només es troba en cèl·lules madures i diferenciades. És una capa on les fibres estan molt ordenades, fet que augmenta la rigidesa de la cèl·lula. Làmina mitjana: capa que uneix les parets primàries de dues cèl·lules. Formada per pectina i hemicel·lulosa.
Cada unió presenta una forma diferent: Bandes d’adhesió (cèl·lula-cèl·lula i actina): es formen bandes al voltant de la cèl·lula, prop de la zona apical i per sobre de les unions estretes. Les cadherines s’uneixen a l’actina pel domini citosòlic, gràcies a proteines ancorants com la beta-catenina, la alfa catenina i la vinculina. La p- 120catenina regula l’activitat de la cadherina. Per unió homofilica, el domini extracel·lular de la cadherina s’uneix a la cadherina de la cèl·lula del costat. Aquesta unió permet que la cèl·lula es contragui i canvi de conformació. Plaques d’adhesió o contactes focals (cèl·lula-matriu i actina): unió molt comuna dels fibroblasts gràcies a les integrines. El cap de la proteïna, que es troba a la matriu extracel·lular, d’uneix de forma directa amb la fibronectina. La cua, al citosol, s’uneix mitjançant unes proteïnes ancorants (tal·lina, filamina i vinculina) a l’actina (a través d’alfa actinina). Permeten un moviment molt dinàmic. Desmosomes (cèl·lula-cèl·lula i filaments intermedis): unions que donen resistència mecànica a la cèl·lula (teixits epitelials madurs). A la superfície de l’interior de la cèl·lula es forma una placa de proteïnes d’adhesió on els filaments intermedis (de queratina o desmina) s’hi enganxen. Les cadherines (desmogleina i desmocolina) uneixen a aquestes plaques (de catenines placoglobina o placofilina i desmoplaquines) i les mantenen gràcies a la intervenció del Ca2+. Hemidesmosomes (cèl·lula-matriu i filaments intermedis): unions de la zona basal de la cèl·lula amb la matriu extracel·lular. Mitjançant integrines, la laminina s’uneix als filaments intermedis de queratina. Proporcionen resistència mecànica als teixits. La integrina alfa6beta4 s’uneix a la queratina per proteïnes enllaçants com la plectina o la distonina. El complex final també inclou col·lagen, que està unit a la laminina.
Unions comunicants o de canal: Unions gap: unions de les cèl·lules animals que permeten el pas d’ions i petites molècules entre cèl·lula i cèl·lula. Responsables de la homogeneïtzació de processos electroquímics i metabòlics entre cèl·lules d’un mateix teixit. Es formen en regions on les cèl·lules estan situades exactament paral·leles i estan unides per complexos proteics que s’anomenen conexines. Aquestes proteïnes tenen 4 hèlix transmembranals que s’uneixen de 6 en 6 per formar conexons. Dos conexons alineats formen un canal aquos continu, el qual permet el pas d’ions inorgànics i molècules solubles. La obertura i tancament d’aquests canals es regula per la concentració de Ca2+ i el pH. S’obren quan la concentració de calci és alta o el pH és baix. Plasmodesmes: unions pròpies de les cèl·lules vegetals que actuen com a unions gap. Canals recoberts per membrana plasmàtica que permeten el pas de petites molècules o macromolècules. Fan que el citoplasma sigui continu. El canal consta d’una estructura central que ve del RE llis: un desmotúbul. Adhesió cel·lular: Són unions temporals, a vegades precursores de les unions cel·lulars. Les CAMs es classifiquen segons la dependència al Ca2+. Les cadherines, les selectines i les integrines ho són. En canvi les immunoglobines són independents a aquest ió. CAMs-Ca2+^ dependents o Cadherines: n’hi ha varis tipus segons el teixit específic que formin les cèl·lules i també segons si participen en unions o adhesions. Quan participen en unions, ho fan en agrupacions però en adhesions ho fan com a molècula individual. Es gràcies al Ca2+ que els dímers d’aquesta proteïna s’activen per interaccionar amb els dímers de la cèl·lula adjacent amb els extrems N-terminal. Es diferencien en dos grups: les clàssiques, que uneixen cèl·lules i també són transmissores de senyals, i les no clàssiques amb seqüències diferents que actuen principalment al cervell. o Selectines: les interaccions que estableixen són heterofíliques ja que s’uneixen a glucoproteïnes o glucolípids de la membrana cel·lular més propera. Les adhesions es duen a terme al torrent sanguini per regular la circulació de leucòcits. Diferenciem les L-,P- i E- selectines, les quals tenen com a domini extracel·lular una cadena de
TEMA 6 COMPARTIMENTS CEL·LULARS I TRÀFIC DE PROTEÏNES
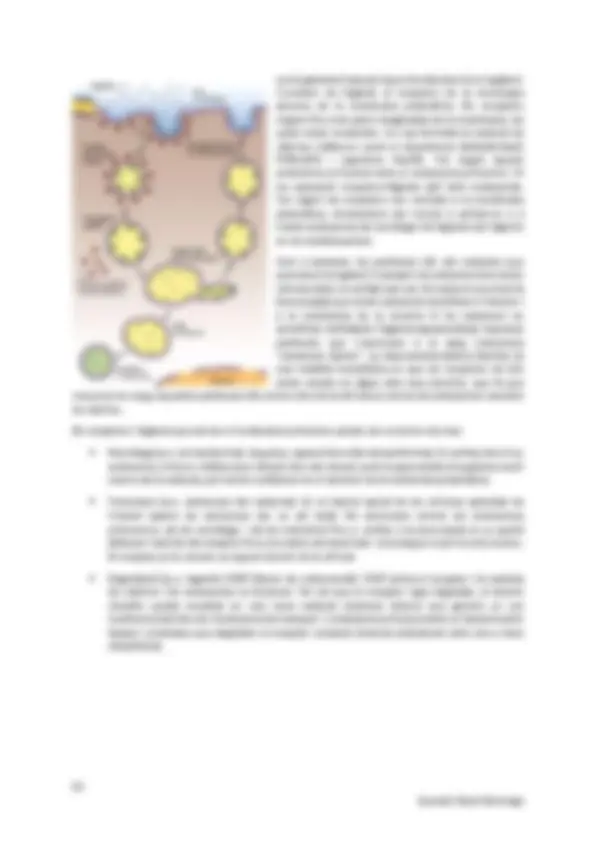
La compartimentació dins de les cèl·lules permet per una banda augmentar la superfície de la membrana, i això permet més superfície per poder dur a terme processos enzimàtics i reaccions. També permet crear orgànuls que tenen diferències en composició i funció. Aquest conjunt de sistemes de l’interior cel·lular eucariota s’explica mitjançant vaires teories. Origen del nucli: En algun moment, es produeix una invaginació de la membrana en que s’envolta el material genètic, formant l’embolcall nuclear, que és una doble membrana. D’aquí sorgeixen el reticle endoplasmàtic, l’aparell de Golgi, lisosomes, endosomes,... Teoria endosimbiòtica: Mitocondris: una cèl·lula protoeucariota anaeròbica acull per simbiosi una cèl·lula procariota aeròbica i es produeix intercanvi de gens entre la procariota i el nucli protoeucariota. Es defineix la dependència i la cèl·lula simbiòtica adquireix forma i funcions de mitocondri. La cèl·lula protoeucariota esdevé una cèl·lula aeròbica. Cloroplasts: una cèl·lula eucariota aeròbica acull una procariota fotosintètica, es produeix intercanvi de gens i es produeix dependència. Es forma el cloroplast i apareix la cèl·lula eucariota fotosintètica. Tràfic intracel·lular de proteïnes: Les proteïnes es sintetitzen totes al mateix lloc i degut a la compartimentació, calen unes rutes de transport perquè arribin a cada orgànul i compleixin les seves funcions.
Importació postraduccional Importació cotraduccional Síntesi als ribosomes i alliberament al citosol Acaben la síntesi al RE o altres compartiments Residents al citosol, al nucli, mitocondris, plastidis, peroxisomes Residents al RE, membrana nuclear, via biosintètica-secretora (a l’aparell de Golgi, membrana plasmàtica, exterior, lisosomes, endosomes)... Tipus de transport: o Regulat per porus: a través dels porus nuclears, les proteïnes solubles arriben al citosol per la via postraduccional. o Transmembranal: el fan les proteïnes postraduccionals dels peroxisomes, plastidis i mitocondiris i també les cotraduccionals del RE. o Vesicular: no travessen el RE i per tant es mouen de RE a AG o compartiments similars. Senyalització de proteïnes: és important per tal que les proteïnes arribin al seu. Son un conjunt de seqüències i senyals que conté la pròpia proteïna o mRNA. Seqüències senyal: es troben als extrems (poden eliminar-se un cop arribi a lloc) o al mig de la cadena Regions senyal: trossos de cadena proteica que un cop la proteïna es plega, s’ajunten formant la regió (no es pot eliminar).