¡Descarga Apunts de C&H vegetal y más Apuntes en PDF de Citología e Histología Vegetal y Animal solo en Docsity!
CAPÍTOL 1: CÈL·LULA VEGETAL
La cèl·lula vegetal té molts components: els cloroplasts i els plasts (on es
realitza la fotosíntesi), la paret cel·lular (que es troba per fora la membrana), etc.
Paret cel·lular (Vegetals) Mantenen la turgència dels teixits, deguda a l’inflament de les cèl·lules per osmosi. La cèl·lula vegetal té tendència a inflar-se. La paret cel·lular impedeix que la cèl·lula rebenti, es tracti d’una paret fina o gruixuda. Les cèl·lules meristemàtiques tenen una paret cel·lular molt fina, com una pel·lícula. Els troncs de les plantes, per exemple, estan formats per cèl·lules amb parets lignificades, degut al fenomen d’esclerificació: enduriment de teixits. Les parets fines són les primàries, i les gruixudes, esclerificades, són les secundaries.
Matriu extracel·lular: és el que substitueix la paret cel·lular en animals, és més complexa.
Cloroplasts: Són un tipus de plasts.
Vacúols
Emmagatzemen aigua i diferents soluts. Són els responsables de la turgència de la cèl·lula. No hi són presents en totes les cèl·lules. En les cèl·lules vegetals són relativament grans, però també depèn del tall.
TEIXIT FOLIAR
VEGETALS
Les cèl·lules estan plenes de cloroplasts. Aquests cloroplasts tenen un ordre
i una densitat diferents en cada fulla, degut a la incidència de la llum. Amb il·luminació intensa (30 minuts), observem que els cloroplasts estan
enganxats a la paret, mentre que amb menys il·luminació els trobem
dispersos en la cèl·lula. Aquest fenomen suggereix dues coses:
- Els cloroplasts es poden moure per dins el teixit cel·lular, com tots els orgànuls.
- Quan hi ha massa llum, l’eviten (L’eficàcia de la fotosíntesi correspon a una quantitat específica de llum).
PLASTIDIS (O PLASTS)
Cloroplasts (de color verd)
Amiloplasts (rics en midó, no són verds, no tenen color)
Cromoplasts
El midó té tendència a formar grans de midó amb tenen línies
concèntriques, que indiquen que el creixement d’aquest gra és discontinu.
La patata té arrels tuberoses, i aquestes són rics en amiloplasts.
CLOROPLASTS
Els cloroplasts tenen doble membrana, més gran que el mitocondri: la
membrana interna i la membrana externa. A part, tenen la membrana del
tilacoide o tilacoidal, on es realitza la fotosíntesi. També hi ha els
estromatilacoides (o tilacoides del estroma), els granatilacoides (tilacoides dels grana), l’espai intertilacoidal, l’espai intermembrana i l’estroma. Els
tilacoides dels grana formen piles, cada pila s’anomena grànum, i el conjunt
de grànum s’anomena grana. En el tilacoide hi ha una alta concentració de protons, mentre que a
l’estroma aquesta concentració és baixa. L’espai intertilacoidal és un espai
continu, els grana estan units als estromatilacoides. Un grànum normalment té 10, 12 o 15 granatilacoides. Cada grana és una mena de pàrquing amb
moltes plantes (granes), amb moltes sortides en forma de rampes
tilacoidals helicoidals, que són els estromatilacoides.
A la membrana, hi ha pigments, ATP sintetases, etc. Aquests pigments capten diferents λ. Els pigments poden ser altres que clorofil·les.
Les arrels acumulen reserves de sucre i de midó.
VEGETALS
D’acord amb models recents, les molècules de carotens (C) queden segrestades al nucli dels túbuls, la coberta dels quals consisteix en una monocapa de lípids polars (PL) en la que s’introdueixen unes proteïnes específiques, anomenades proteïnes associades als carotens (CAP). La funció de les CAPs és de protecció, fent d’escut, de
forma semblant a les oleosines dels cossos lipídics.
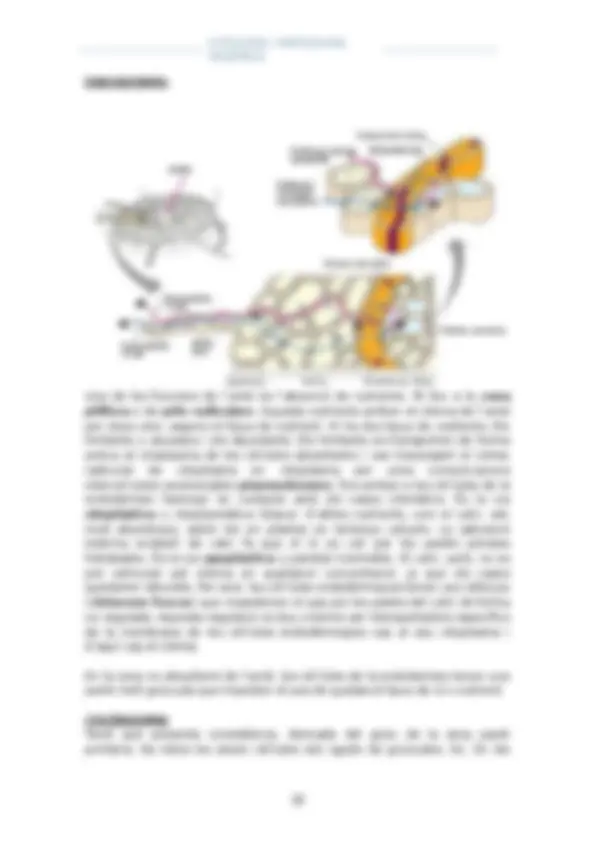
INTERCONVERSIÓ PLASTIDIAL
A: Proplast B i C: cloroplast D: Cloroplast en divisió E i F: etioplast G: amiloplast H: cromoplast
Els colors no verds dels vegetals són pigments no verds, tancats en vacúols o en cromoplasts. (Com les flors i els fruits)
El protoplast correspon a les cèl·lules de l’embrió. Té midó. L’embrió, es va desenvolupant i té la radícula, la tija embrionària i els cotiledons. Els cotiledons són les primeres fulles i diferenciaran els cloroplasts. En la radícula no interessa que hi hagi cloroplasts, sinó que hi ha amiloplasts.
Evolució secundària
Dels cloroplasts dels cotiledons sorgeixen els cromoplasts en els fruits. El
cromoplast pot tornar a ser cloroplast.
E I F: etioplasts. Són un tipus de plastidis que es donen en els mateixos
cotiledons o en òrgans verds de la planta que se’ls hi priva de llum. Desapareix el cloroplast. Presenten una variant de clorofil·la que no és de
color verd. Etiolació.
Tenen doble membrana i membranes internes en forma de túbuls, i aquests túbuls tenen aquest pigment clorofíl·lic no verd. No agafen cap tipus de
color.
La etiolació és reversible, és a dir, que si ha una planta blanca li tornem a
incidir llum, la clorofil·la normal torna a aparèixer i apareix color verd.
La etiolació també la podem fer a partir de l’embrió, a part de a partir dels
cotiledons. Llavors els cotiledons d’aquella planta no seran de color verd.
VEGETALS
MITOCONDRIS
La principal funció de la cadena respiratòria dels mitocondris, en cèl·lules
animals o vegetals, es convertir el poder reductor en forma de NADH o
succinat en molècules d’ATP. Una cadena respiratòria esta formada per tota una sèrie de molècules inserides en la membrana interna mitocondrial per
les quals hi ha un transport electrònic, transport que inclou una colla de
reaccions redox exergòniques, l’energia despresa de les quals s’inverteix en bombejar protons a l’espai intermembrana i crear així un gradient protònic
que servirà per a fabricar ATP. En tot mitocondri hi ha doncs grans
complexes proteics, multienzimàtics, entre els quals té lloc un transport
electrònic, transport en el que intervenen molècules més petites i mòbils com la ubiquinona (UQ) i el citocrom c (en blanc en la figura). Els complexes
grans són la NADH deshidrogenasa (I), la succinat deshidrogenasa (II) que
utilitza com a coenzim font d’electrons la FADH2, el complex III o complex citocrom b/c i el complex IV o citocrom c oxidasa (COX), que lliura els
electrons que han viatjat per la cadena a l’oxigen per a formar aigua. El
complex V és l’ATP sintetasa. En els mitocondris vegetals hi ha uns components addicionals que permeten
rutes alternatives de transport d’electrons (en negre en la figura). Dues
NADH deshidrogenases (40 i 41) que oxiden el NADH en la matriu i en el
citosol, respectivament, i no bombegen protons. Ambdues són resistents a la rotenona. I una oxidasa alternativa (AOX) (44) que com l’altra oxidasa
també
lliura els electrons de la cadena a l’oxigen per a formar aigua. Aquesta oxidasa alternativa tampoc bombeja protons i és resistent al cianur.
El significat d’aquests compostos alternatius està discutit. Donat que
l’energia despresa en el transport d’electrons entre aquests components no s’inverteix en bombejar protons, es perd en forma de calor. Són doncs rutes
alternatives termogèniques, que en òrgans florals poden ser útils per a
volatilitzar compostos olorosos, cosa que facilita l’atracció dels insectes
pol·linitzadors.
VEGETALS
TRANSPORTADORS DEL TONOPLAST
Canals: iònics i aquosos (aquaporines, TIPs) Co-transportadors: antiporters i simporters Bombes protòniques: ATPàsiques i pirofosfatàsiques Transportadors ABC (ATP Binding Cassette)
SISTEMA
D’ENDOMEMBRANES
La cèl·lula vegetal conté un sistema d’endomembranes ben
representat. Els materials són
transportats cap als diferents vacúols mitjançant la ruta secretora, per la qual es vehiculen els
components de la paret.
TIPS VACUOLARS
La naturalesa vacuolar dels orgànuls de reserva ve donada per la presència en la seva membrana de TIPs (Tonoplast Integral Proteins).
- Les lletres greques corresponen a les diferents TIPs i així els vacúols de reserva proteica (PSV) i els vacúols lítics (LV) estan limitats per tonoplast amb llurs TIPs específics.
- Els orgànuls amb δ-TIP representen vacúols no de reserva.
- En l’esquema hi figura també la ruta secretora constitutiva (default) Aquaporines (TIPs): canals d’aigua, importants i diferents en cada
vacúol. Tenen una estructura molt complexa. Ex: El ronyó és riquíssim en aquaporines.
Els vacúols com a dipòsits de reserva proteiques emmagatzemen ATP. Una
acumulació de reserves significa una acumulació de proteïnes.
La síntesi de proteïnes es fa al reticle endoplasmàtic o al citosol. Després,
passen per l’aparell de Golgi, d per les vesícules i finalment poden anar cap
VEGETALS
a fora la cèl·lula, cap a dins la cèl·lula o quedar-se a la membrana. És a dir:
secreció, membrana o interior cel·lular. Si es queden a l’interior cel·lular, les proteïnes van als lisosomes (animal) o als vacúols lítics (vegetal). Aquests
són uns vacúols de reserva proteica, de sucre, que acumulen proteïnes per
digerir, anomenades hidrolases àcides. Els vacúols d’aigua permeten
augmentar la pressió osmòtica, i canviar la forma de la cèl·lula. Hi ha dos tipus de proteïnes, unes que van directament cap al vacúol i unes que no.
Tot i la diversitat de vacúols, tots estan limitats pel seu tonoplast. Els
vacúols es poden fusionar i com a conseqüència es secreten proteïnes.
DRUSES D’OXALAT
En molts teixits verds de la planta, com a conseqüència del fenomen de la
fotorespiració (una desviació del procés fotosintètic) es produeix l’àcid
oxàlic, altament tòxic per a la cèl·lula.
Per això, aquest àcid s’acumula i s’immobilitza als vacúols on, degut a la concentració relativament alta de calci, precipita en forma de cristalls
d’oxalat càlcic molt característics.
PARET PRIMÀRIA
Growing Plant Cell Wall (en català l’anomenem paret primària ). Totes les cèl·lules vegetals tenen partit primària. Es diposita quan la cèl·lula creix. Està fora la cèl·lula. És un producte de secreció de la cèl·lula, des de les tires de col·lagen. Tots els components es sintetitzen a la cèl·lula mitjançant diferents mecanismes. La majoria de les cèl·lules només tenen paret primària, però alguns teixits tenen paret secundària , com el tronc dels arbres Fa entre 100 i 200 ηm. Podem descriure-la com una esponja, un gel hidratat, adaptable al creixement i que té porositat.
El tonoplast comunica amb altres cèl·lules veïnes. Entre dues parets
cel·lulars hi ha la làmina mitjana.
Està constituïda principalment per polisacàrids:
VEGETALS
galactosa que té un grup carboxil i s’anomena àcid galacturònic. Les
pectines tenen moltes càrregues negatives, per tant agafen aigua de solvatació i la paret esdevé un gel. Les pectines ramificades tenen
Ramnogalacturonano. També tenen grups acetil i metil. Així doncs, la paret
cel·lular dóna un mitjà aquos a la cèl·lula.
PLASMODESMATA
Hi ha d’haver una comunicació intracel·lular, tot orgànul aïllat està mort. Per tant hi ha uns ponts o canals que permeten una continuïtat de citoplasma i
de membrana. Aquests braços citoplasmàtics reben el nom de
plasmodesmes , i sempre van en grup. A més també han de fer la funció de canalitzar el reticle endoplasmàtic d’una cèl·lula a un altre, mitjançant un
tub anomenat desmotúbul. Els plasmodesmes són anàlegs als connexons
de les cèl·lules animals.
A la cèl·lula no hi ha res estàtic. Els plasmodesmes poden estar més o
menys tancats: oberts (més comunicació) o tancats (menys comunicació). El
que obra o tanca el plasmodesma és un dipòsit de sucre que rep el nom de
cal·losa , que té enllaços β 1-> 3. Aquests enllaços fan la cèl·lula boteruda, una massa que no té forma. Normalment els plasmodesmes es presenten
agrupats en camps de porus primaris (“primary pit fields”).
Solvatació : La solvatació és un fenomen que es produeix en una dissolució i que consisteix en l'atracció, mitjançant forces intermoleculars ió-dipol o
dipol-dipol, de les molècules de dissolvents i els ions o molècules de solut.
La pectina, concretament els àcids galacturònics que la formen, tenen un grup carboxil ionitzat amb una càrrega negativa, a més a més d’altres
càrregues provocades per altres molècules. Per tant, quantes més
càrregues, més aigua de solvatació.
PARET SECUNDÀRIA O GRUIXUDA:
Paret cel·lular que es deposita quan la cèl·lula ha crescut. Totes les cèl·lules amb paret cel·lular secundaria esdevenen mortes, al final només queda la paret. És una paret gruixuda. És un producte de secreció. Es forma a base d’uns estrats interns molt més gruixuts. Estrat S1 (el primer que es deposita), S2 i S3. El creixement és en la direcció interna. La membrana sempre queda per dins.
En la paret secundària no parlem de microfibril·les, ja que no està en creixement continu. Tenim doncs, cel·lulosa, poques hemicel·luloses i
poques pectines. La cel·lulosa està organitzada en fibra de cel·lulosa: un feix
de microfibril·les, que al seu torn formen la fibril·la, que pot tenir diferents
VEGETALS
orientacions. Aquesta paret pateix un procés de lignificació, ja que els seus
porus estàn ocupats per lignina, i la paret esdevé no porosa. Així doncs la cèl·lula es mor perquè no té aigua, només queda la paret secundària.
LIGNINA
La lignina no és un polisacàrid, és una xarxa formada per un polímer, els
monòmers del qual són els monolignols: compostos de pes molecular baix,
tenen grups alcohols i grups fenols. Aquests monolignols s’uneixen per formar el polímer tridimensional, que es cola per les fibres de cel·lulosa de
la paret secundària, impermeabilitzant la paret. És un producte de secreció
de la cèl·lula que es forma quan la cèl·lula està molt activa. La cèl·lula forma els monolignols i aquests es segreguen cap a fora. Van en sentit contrari al
de creixement, així doncs passen a través dels S1, S2 i S3. Però primer es
lignifica la paret primària, després l’S1, l’S2 i finalment l’S3 ( a vegades l’S no es lignifica).
PUNTUACIONS
Les vies de comunicació entre les cèl·lules que tenen paret secundària són
puntuacions , interrupcions de la paret secundària que constitueixen vies de comunicació entre dues cèl·lules veïnes amb aquesta paret. La paret primària (en groc) roman intacta en el sí de la puntuació, però la seva porositat permet el pas d’aigua i soluts entre ambdues cèl·lules. Les puntuacions solen coincidir amb el camp de plasmodesmes inicials primaris.
VEGETALS
CAPÍTOL 2: SISTEMA HISTOLÒGIC
DÈRMIC
Tot òrgan vegetal té sistemes externs, protectors, interns, etc.
A la superfície hi ha el sistema histològic dèrmic. A l’interior hi ha els teixits vasculars, que porten soluts i formen el sistema
histològic vascular.
Entre el superficial i l’intern, hi ha el sistema histològic fonamental.
Cada sistema està format per varis teixits, aquesta classificació deriva de la
topografia.
CÈL·LULES DE L’EPIDERMIS EN ÒRGANS DE CURTA DURADA
EPIDERMIS DE LA FULLA.
(1) Si tallem tangencialment, és a dir, fem un tall paral·lel a la superfície, secció periclinal, veiem aquesta imatge: L’epidermis amb la forma de les cèl·lules. L’epidermis és un teixit superficial. Tipus de cèl·lules: cèl·lules oclusives dels estomes i cèl·lules epidèrmiques ordinàries. L’Epidermis té una sola capa de cèl·lules.
VEGETALS
(2) En canvi, si fem un tall perpendicular a la superfície, secció transversal, veiem unes cèl·lules més grans a dalt de tot, la forma de les quals és aplanada i a més entre cèl·lula i cèl·lula no hi ha espai intercel·lular. El fet que no hi hagi espai intercel·lular no evita la pèrdua d’aigua, i aquí entra en joc una substància impermeable: un lípid, concretament un cèrid. La cutícula és l’impermeable lípid, preserva els teixits interns com a òrgans. Hi ha uns
porus que serveixen per intercanviar gasos.
Cèl·lules epidèrmiques ordinàries. A part de la paret cel·lulòsica, hi ha una altra paret primària: una cutícula que cobreix tota la superfície. La feina
d’aquestes cèl·lules és segregar aquesta cutícula. Són cèl·lules vives amb
nucli que no fan la fotosíntesi, per tant, no tenen plastidis.
Cutícula : dipòsit de tipus lipídic situat per fora de les parets periclinals de
les cèl·lules epidèrmiques ordinàries. La cutícula presenta dues regions:
reticulada i lamel·lar. Per fora de la cutícula hi ha un dipòsit epicuticular de cera , producte de secreció d’aquestes cèl·lules. Les ceres epicuticulars
són de naturalesa parafínica, totalment hidròfobes i no tenen llocs
d’hidratació. Aquestes ceres adopten diferents formes com vírgules o túbuls i plaques. Aquesta cera no està present en tots els òrgans, depèn del clima.
I així, des de les cèl·lules fins a la superfície, s’estableix un gradient de
menys a més hidrofòbia. La cutícula està formada per cutina.
La cutina és un polièster que sorgeix de la unió covalent entre un alcohol i
un grup carboxil d’àcid orgànic amb la pèrdua de H 2 O. És una xarxa formada
per àcids grassos polihidroxílics i interesterificats, és a dir, hidroxils i carboxils esterificats, però no tots els hidroxils s’esterifiquen, així que
queden cadenes polars. Contra més esterificació, més hidròfoba i menys
aigua. Segons els ambients, més o menys secs, el grau d’hidratació de la cutina variarà, així com el gruix de la cutícula.
Cèl·lules oclusives dels estomes. Tenen forma de mongeta, de ronyó. Coexisteixen amb les cèl·lules epidèrmiques ordinàries. Potencialment dues
cèl·lules oclusives es poden obrir molt quan estan aïllades i provocar una
gran pèrdua d’aigua; així que en els estomes, aquestes cèl·lules oclusives
estan envoltades per unes cèl·lules annexes que limiten aquesta obertura.
VEGETALS
Suberina La suberina és una substància lipídica hidròfoba de composició complexa,
que forma un dipòsit per l’interior de totes les parets cel·lulòsiques de les
cèl·lules del suro. Aquestes cèl·lules són doncs cèl·lules mortes. Al TEM
aquest dipòsit presenta lamel·les, degut a que la suberina s’interestratifica amb ceres.
Lenticel·les Les lenticel·les són les estructures de comunicació entre el teixits vius
interns i l’exterior, separats pel suro, teixit mort i impermeable. Són llocs
que trenca el suro, les cèl·lules queden soltes i deixen espais intercel·lulars per on es realitza l’intercanvi gasos.
CAPÍTOL 3: SISTEMA HISTOLÒGIC FONAMENTAL
PARÈNQUIMA
El parènquima és un teixit del sistema histològic fonamental que
topogràficament separa la epidermis dels feixos vasculars. Conté la major
part de la massa de les cèl·lules, és el teixit que majoritàriament ocupa tot el volum d’un òrgan. Les cèl·lules d’aquest teixit tenen paret primària molt
prima. Segons l’òrgan, pot tenir diferents funcions i pot ser ric en diferents
orgànuls. Així en les fulles i de fet en qualsevol teixit verd de la planta, el parènquima, anomenat mesòfil o clorènquima , és ric en cloroplasts o
plastidis verds. En les arrels el que domina és la presència de plastidis rics
en midó, o amiloplasts, i aleshores parlem de parènquima amilaci. No obstant això, l’habitual és que el parènquima sigui senzillament de
replenat , contribuint amb la seva turgència cel·lular a la consistència
mecànica del teixit.
1. Parènquima de replenat : Cèl·lules que no tenen gaires plasts ni
reserves, però donen molta turgència cel·lular al teixit. Tenen un vacúol
central i paret primària molt prima. Són com bombolles, de forma polièdrica. Aquestes cèl·lules, en un òrgan allargat com les tiges i les arrels, tindran
forma allargada.
2. Parènquima clorofíl·lic, clorènquima o mesòfil : Les seves cèl·lules
tenen moltíssims cloroplasts. Cèl·lules allargades i molt estretes. Amb dos o
VEGETALS
tres capes. Els estomes estan ubicats al revers perquè tenen més protecció
en contra de la pèrdua d’aigua, però hi ha excepcions. Hi ha espais en els quals s’acumula el CO 2.
Palissada: anvers Esponjós: revers
El mesòfil en palissada, propi de l’anvers de les fulles de dicotiledònia, és un
teixit compacte, amb parets cel·lulòsiques molt juntes, formant una massa
tissular molt hidratada. Per contra, el mesòfil esponjós del revers de la fulla,
presenta grans espais intercel·lulars plens d’aire, espais a prop dels estomes. Això fa que l’anhídrid carbònic, gas essencial per la fotosíntesi, es
retingui amb més facilitat dins d’aquests espais. No seria possible això si el
teixit del revers fos compacte.
3. Parènquima en arrels. No tenen cloroplasts, tenen plasts de reserva de
tipus glúcid, midó. Les arrels es dediquen a absorbir.
VEGETALS
costelles longitudinals de les tiges, hi ha molta colènquima. VEURE
COL·LÒCIT I COL·LENQUIMATITZACIÓ.
Tipus de col·lènquima
- Col·lènquima angular (engrossit en els punts de contacte intercel·lular).
- Col·lènquima tangencial (cèl·lules disposades en files ordenades i
engrossides a la cara tangencial de la paret cel·lular).
- Col·lènquima llacunar (tenen espai intercel·lular i engrossiment proximal cap a l'espai intercel·lular).
Les cèl·lules col·lenquimàtiques es troben més freqüentment adjacents a l'exterior dels teixits de creixement, al càmbium vascular.
ESCLERÈNQUIMA
Les seves cèl·lules estan mortes, per tant doncs són molt especialitzades. Estan lignificades, i per tant només els queda la carcassa de la paret.
Aquesta estructura dóna consistència a la fusta, a les crostes de les llavors,
etc. Hi ha dos tipus de cèl·lules:
Cèl·lules allargades : són fibres, es troben en els òrgans allargats, sota la parènquima de cèl·lules allargades. Les fibres esclerenquimàtiques formen la fusta.
Cèl·lules no allargades : esclereides. D’aquestes ultimes n’hi ha de varies formes. Braquisclereides (en forma de pilota): es troben en les llavors. Astrosclereides (en forma d’estrella): es troben en les parènquimes aerífer, que tenen espais buits plens d’aigua. Donen suport a aquestes cèl·lules. El nenúfar té molta parènquima aerífer.
La lignificació és la polimerització dels monolignols. Aquests es sintetitzen
en la cèl·lula i la polimerització té lloc a la paret. Normalment aquesta és
primària i secundària. La lignificació comença en la paret primària i centrípetament es va extenent cap els estrats de la paret secundària mes
interns o més propers a la membrana. El polímer s’intrueix per entre les
microfibril·les de cel·lulosa. La lignificació és doncs una modificació de la
paret per incrustació. Per contra, la cutinització i la suberificació són modificacions de la paret per adcrustació.
VEGETALS
CAPÍTOL 4: SISTEMA HISTOLÒGIC VASCULAR
El xilema i el floema són feixos vasculars. En la imatge, les cèl·lules que tenen la paret gruixuda són les del xilema. En un creixement secundari (troncs i altres) el xilema i el floema estan separats, a diferencia de en un creixement primari. El xilema i el floema són teixits compostos.
FLOEMA
El floema consisteix en una sèrie de tubs que porten els productes de la
fotosíntesi en direcció descendent, anomenats fotoassimilats. No obstant, aquesta direcció pot ser ascendent quan vinguin de les fulles i de les arrels,
les quals actuen com a òrgans de claveguera. Les cèl·lules que en formen
part són vives. Donat que el sistema vascular està format per conductes
cribosos , a vegades molt llargs, les diferents cèl·lules que formen aquests conductes han d’estar a la força comunicades entre sí. En el cas del floema,
teixit conductor encarregat del transport de la solució aquosa de
fotoassimilats o saba elaborada, les cèl·lules dels conductes, anomenades elements cribosos , estan comunicades entre sí per uns plasmodesmes
modificats anomenats porus cribosos, agrupats en àrees criboses en les
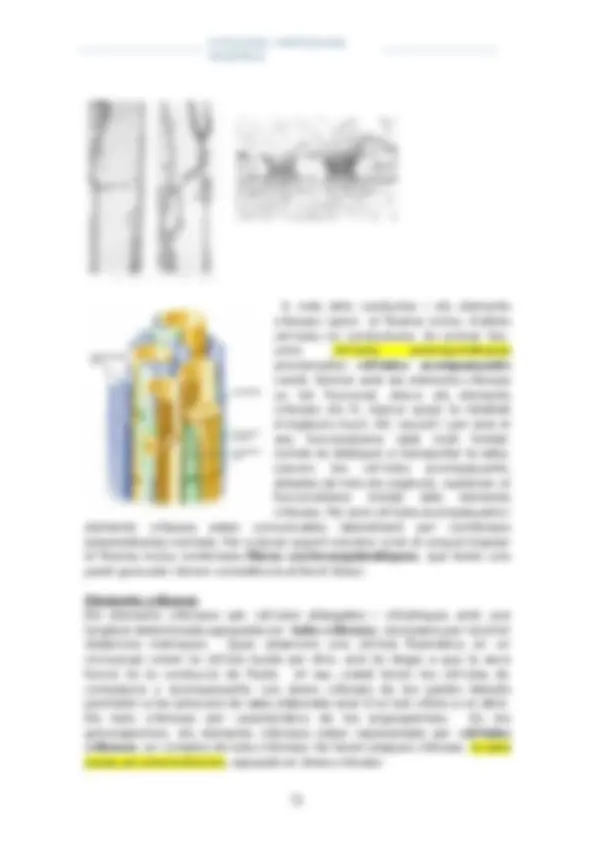
parets transversals anomenades plaques criboses. En la electromicrografia de l’esquerra s’observen les plaques criboses d’ambdós
conductes cribosos; les fletxes assenyalen un precipitat proteic exclusiu dels
elements cribosos. Es tracta de la proteïna F o P. En la electromicrografia de
la dreta es mostra un detall de dos porus cribosos on s’ha concentrat la proteïna F. Les estructures més fosques són acumulacions de sacarosa i
proteïnes F (phloem protein). Aquestes proteïnes coagulen i tapen les
plaques criboses en condicions anormals (quan fem un tall histològic, trenquem el floema i malmetem els tubs cribrosos, i com a mostra de
seguretat, la proteïna coagula i tapa les plaques criboses).
VEGETALS