



Descargado en:
patatabrava.com
EVOLUCIO (UDG)
EVOLUCIO
VIDAL, ORIOL 12-13




























































































Prepara tus exámenes y mejora tus resultados gracias a la gran cantidad de recursos disponibles en Docsity

Gana puntos ayudando a otros estudiantes o consíguelos activando un Plan Premium

Prepara tus exámenes
Prepara tus exámenes y mejora tus resultados gracias a la gran cantidad de recursos disponibles en Docsity
Prepara tus exámenes con los documentos que comparten otros estudiantes como tú en Docsity
Encuentra los documentos específicos para los exámenes de tu universidad
Estudia con lecciones y exámenes resueltos basados en los programas académicos de las mejores universidades
Responde a preguntas de exámenes reales y pon a prueba tu preparación
Consigue puntos base para descargar
Gana puntos ayudando a otros estudiantes o consíguelos activando un Plan Premium
Comunidad
Pide ayuda a la comunidad y resuelve tus dudas de estudio
Ebooks gratuitos
Descarga nuestras guías gratuitas sobre técnicas de estudio, métodos para controlar la ansiedad y consejos para la tesis preparadas por los tutores de Docsity
Asignatura: Evolucio, Profesor: ORIOL ORIOL, Carrera: Biologia, Universidad: UdG
Tipo: Apuntes
1 / 109

Esta página no es visible en la vista previa
¡No te pierdas las partes importantes!
Descargado en:
patatabrava .com
EVOLUCIO (UDG)
EVOLUCIO
VIDAL, ORIOL 12-
3.5. Canvis en les freqüències gèniques III. Grandària real i grandària efectiva d'una població. Deriva genètica. Fixació d'al·lels. Efecte fundador i coll d'ampolla.
3.6. Diferenciació interpoblacional. Fragmentació en les poblacions: efecte Wahlund. Índex de fixació: Fst. Divergència genètica i diferenciació poblacional.
4. L'ESPÈCIE I EL SEU ORIGEN
4.1. L'espècie. Concepte històric d'espècie. Concepte morfològic. Concepte biològic. Concepte evolutiu. Semiespècies. Espècies críptiques.
4.2. L'aïllament reproductiu. Concepte. Mecanismes d'aïllament prezigòtic: ecològic, temporal, etològic, mecànic i gamètic. Mecanismes d'aïllament postzigòtic: inviabilitat i esterilitat dels híbrids.
4.3. Origen de noves espècies. Especiació al·lopàtrica o geogràfica. Zones híbrides. Especiació per efecte fundador. Els pinsans de Darwin. Especiació no geogràfica: especiació parapàtrica. Especiació simpàtrica.
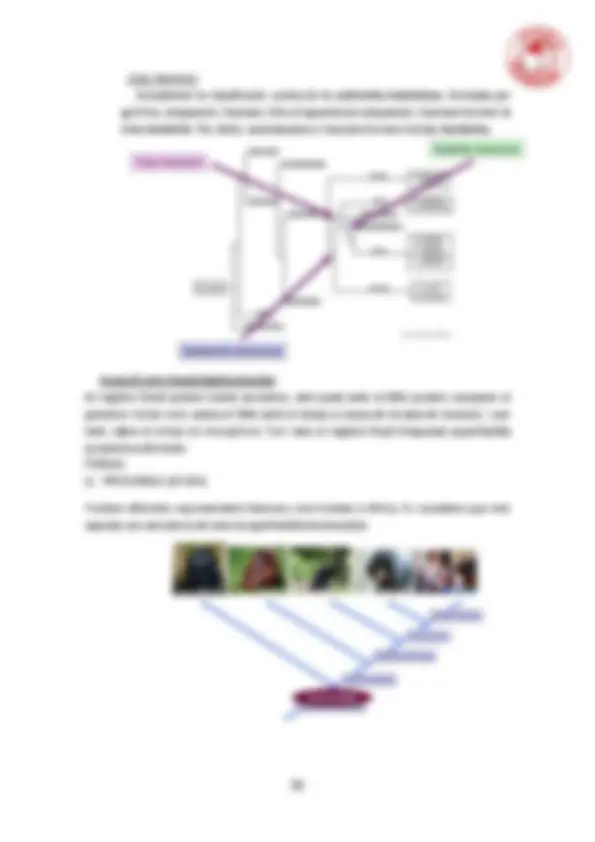
5.1. Definicions. Conceptes de taxonomia, sistemàtica i classificació. Anagènesi i cladogènesi. Caràcters ancestrals i derivats. Homologia i homoplàsia.
5.2. Classificació biològica. Les escoles evolutiva, genètica i cladista de classificació biològica. Monofiletisme, parafiletisme i polifiletisme.
5.3. Filogènia. Tipus de dades. Mesures de distància. Distància genètica. Estadístics d'agrupament. Arbres sense arrel. Determinació de la polaritat. Màxima parsimònia.
5.4. Biogeografia evolutiva. Patrons geogràfics de distribució. Causes de les distribucions geogràfiques. Dispersió versus vicariança. Cladística. Deriva continental. Intercanvi de la fauna americana.
5.5. Origen de les novetats evolutives. Gradualisme filètic. Equilibri puntuat. Estasi i especiació. Fòssils vivents. Embriologia, desenvolupament i evolució. Estructures vestigials.
Darwin va anunciar 4 postulats : (els títols dels postulats no els va anomenar així, ja k aquests termes encara no eren coneguts com a tals)
Darwin va escriure la seva autobiografia quan ja era gran, i va ser ajudat pel seu fill. També va escriure sobre altres temes, com per exemple de la reproducció dels coloms o de les orquídies. També sobre els cirrípedes, on va recol·lectar fòssils i va fer la primera descripció d’un hermafrodita.
Els 3 llibres que van tenir molta importància en Darwin van ser :
Alfred Russel Wallace (1823-1913) enviava des de Malasia mostres d’ individus a museus i d’ aquesta manera cobrava. És a dir, feia el mateix que Darwin. Al 1858, Alfred va enviar a Darwin un article anomenat “La tendència de les varietats, d’ allunyar-se de la forma original” per tal que s’ el llegís i li donés l’ opinió. Darwin va veure-hi reflectit moltes de les seves pròpies idees, de manera que al 1859, va publicar la “ teoria de l’ origen de les espècies ” al Journal of the Linnean Society. No va tenir una gran repercussió a nivell de societat ja que ho llegia molt poca gent. És per això, que va fer un llibre perquè ho pogués entendre tot el món (1859), i va ser d’aquesta manera, que va tindre una gran repercussió. Després de la publicació del llibre, al 1860 es va revolucionar l’ església. Per això, la primera teoria de l’ evolució es diu és de Darwin-Wallace. Aquesta va ser reformada més tard, rebent el nom de teoria moderna de l’evolució.
El viatge amb el Beagle :
x El Beagle va sortir el 27.12. 1831 i va tornar el 2.10. 1836. x Quan el vaixell parava a un lloc per tal de prendre les mesures cartogràfiques , que era el seu objectiu, Darwin baixava per fer la seva feina. El que feia els dibuixos era Conrad Martens. x A val paraíso va viure un terratrèmol. Va fer una excursió als Andes abans i després d’aquest, i això li va permetre veure els canvis geològics. x En una expedició, es van emportar amistosament algun natiu, i els van portar a la civilització amb l’objectiu de culturitzar-los.
El viatge al Beagle li va donar l’oportunitat a Darwin de poder observar i pensar sobre el gran rang de diversitat dels organismes i de les formacions geològiques. Durant el viatge, ell portava els PRINCIPIS DE GEOLOGIA de Lyell. Una experiència que va tenir un gran impacte sobre el pensament de Darwin sobre l’evolució va ser l’estada a les illes Galápagos , on va recollir animals, els quals destaquen els “ pinsones ” (ocells) que va portar dissecats el 1841. Altres animals k va trobar són tortugues gegants i iguanes.
El vaixell Beagle: Era un bergantí de 28m, 1817. Va fer 3 viatges al món. El que va anar-hi en Darwin era el segon. Després d fer el tercer viatge, el vaixell va ser destinar a fer treballs fins que va ser abandonat al desguàs, i per això no es pot veure en l’actualitat.
1.Creacionisme :
El catastrofisme accepta el creacionisme, és a dir, l’aparició de noves espècies del no res. Cuvier defensava el catastrofisme, ja que va ser el fundador de l’ anatomia comparada, anti-evolucionista.
3. Fixisme (Linné, finals s. XVIII) : - Defensa el fet de que les espècies són fixes, úniques i invariables. També defensa l’ immutabilitat i l’ invariació entre els individus d’ una mateixa espècie.
Desenvolupa el concepte de Linneo, el concepte tipològic d’espècie (sistema binomial).
4. Progressionisme. - Defensa que tot evoluciona de l’ésser més simple cap més complexa. Per això a l’home el pinten com el ésser superior i més evolucionat que hi ha hagut mai.
El progressionisme no té sentit, ja que l’ atzar juga un paper molt important. Un argument de que aquesta teoria no és bona, és el fet de que si en l’ època dels dinosaures no hi hagués hagut totes les catàstrofes, l’ésser humà no hagués aparegut.
1.variabilitat morfològica: Un exemple de variació discontinua serien les papallones de color fosc i les blanques (contaminació Æ unes són afavorides xk els seu depredador no les veu, etc). Un altra exemple seria el dimorfisme sexual, el color dels pètals d’ una planta, la cresta dels galls (són d’ 1 tipus o altra) ... Un exemple de variació contínua seria l’ alçada en els individus, la variació gènica (ja que es pot calcular la freq. al·lèlica).
Les variacions a nivell d’ àcids nucleics es poden mirar mitjançant la seqüenciació d’ ADN (PCR), ja que així s’ obté l’ ordre de les bases i es pot observar si hi ha hagut alguna modificació en l’ ordre.
Les tècniques per a comprovar variacions a nivell de proteïnes són varies. En el següent full s’ en mostren algunes :
Tipus de variacions/variabilitat :
Tècnica Material de prova Usat per identificar diferències entre
Limitacions
Electroforesi Proteïnes
Individus
Poblacions
Espècies
Mètode indirecte
Subestima la quantitat de variabilitat genètica, ja que detecta 1/3 de les mutacions (substitucions d’aminoàcids)
Immunològica (antigen - anticòs)
Proteïnes Espècies Mètode indirecte
Seqüenciació d’aa Proteïnes Espècies Mètode indirecte Ens permet saber l’ordre dels aa d’una proteïna. Seqüenciació d’ADN: nuclear, mitocondrial o cloroplàstic.
ADN codificant i
ADN no codificant
Població
Espècies
Mètode directe Cost i temps elevats (PCR)
Empremptes dactilars moleculars (minisatèl·lits o VNTR = repeticions de nº variable amb tandems.
Regions altament variables d’ ADN i pròpies de cada individu
Individus
Només utilitza una part del genoma.
RiFLiPS Æ polimorfisme en els fragments de restricció
S’ usen enzims que tallen el genoma (enzims de restricció), aconseguint talls de DNA
DNA mitocondrial, cloroplàstic o nuclear.
Individus
Espècies
Només utilitza una part del genoma, aquella que els enzims han reconegut i tallat.
Hibridació (DNA-DNA) ADN
Espècies.
Mesura la distància genètica entre espècies
El grau d’ hibridació dependrà de la proximitat filogenètica entre individus. Com + hibridació hi hagi, + semblants seran genèticament
- Una repetició en tàndem seria en grups. Ex : CCTCCTCCT = (CCT) 3 - Una repetició que no és en tàndem és quan una certa seqüència es repeteix però de forma dispersa, és a dir, no consecutivament.
Cal dir que hi ha un volum de dades i arguments + ampli que el model clàssic.
A la natura, la majoria de poblacions presenten una dinàmica de la població + semblant al model d’ equilibri.
VARIABILITAT CROMOSÒMICA:
Polimorfismes cromosòmics :
Causats per inversions i translocacions.
x Inversions = un fragment d’ un cromosoma és invertit, de manera que es modifica l’ ordre.
Exemples :
1. Drosophila suboscura (mosca de la fruita) Æ Els polimorfismes cromosòmics es solen fer en insectes, ja k al tenir cromosomes gegants, és + fàcil la detecció de polimorfismes (solen ser inversions). 2. Keyacris scurra (escarbat) Æ Trobem inversions en 2 cromosomes diferents. En el cromosoma anomenat CD i en l’ anomenat EF. Parlarem del cromosoma EF : - Cromosoma Sr = estàndard. - Cromosoma EF = cromosoma Sr que ha patit una inversió.
Si l’ individu ha patit una inversió, serà heterozigot per l’ inversió, de manera que tindrà un cromosoma Sr i un EF. L’ efecte és que l’ insecte és més petit.
Si l’ individu pateix dues inversions, serà homozigot per l’ inversió, de manera que tindrà dos cromosomes EF. L’ efecte és que l’ insecte mort, ja que l’ homozigosi per l’ inversió és letal.
3. Perognathus goldmani (ratolí de camp) Æ Es van trobar 5 polimorfismes cromosòmics diferents, és a dir, 5 tipus de cromosomes en individus de la mateixa espècie. Aquests cromosomes es diferenciaven entre ells ja que uns eren acrocèntrics, altres metacèntrics, i altres submetacèntrics. - Acrocèntric = té el centròmer a l’ extrem. - Metacèntric = té el centròmer al mig. - Submetacèntric = té el centròmer al mig, però té 1 braç més llarg que l’ altre.
El que van observar és que cada tipus de modalitat cromosòmica, corresponia a un conjunt d’individus que ocupaven una zona geogràfica concreta. Amb això van arribar a la conclusió de que hi havia una de les poblacions que era l’ ancestral.
Bandeig cromosòmic = tècnica que es pot usar per demostrar que entre 2 cromosomes s’ ha donat una fusió cèntrica, o per demostrar que 1 cromosoma ha patit una inversió.
BLOC 3: GENÈTICA DE POBLACIONS
Característica Individus Poblacions Temps de vida 1generació (xk l’ individu neix, es reprodueix i mort)
Moltes generacions (fins que l’espècie s’ extingueix)
Continuïtat espacial (massa) Limitada al volum corporal Pot arribar a ser molt gran
Característiques genètiques Genotip Freqüències al·lèliques
Variabilitat genètica Cap (però si que la té si l’ individu és heterozigot)
Considerable
Caract. evolutives No evoluciona Pot evolucionar (i si ho fa, ho farà en les freq. al·lèliques)
Podem parlar de 2 tipus de poblacions, les naturals i les Mendelianes :
1.Poblacions naturals : conjunt d’individus salvatges units entre si per vincles de reproducció i de parentesc.
Quan un home manipula a una població, aquesta deixa de ser natural.
2.Població Mendeliana: és la unitat de treball de la genètica de poblacions.
La població Mendeliana ideal presenta una sèrie de característiques:
o La mida poblacional (N) és constant al llarg del temps (al llarg de les generacions). o És de generacions no solapades en el temps. / El temps de generació és l’edat a la que es té el primer fill. o La població ha d’estar formada per individus ideals , els quals són aquells que s’aparellen aleatòriament entre ells , és a dir, que no hi ha cap factor que provoqui amb + intensitat la reproducció.
Si hi ha alguna cosa que pertorbi a la població, les freqüències al·lèliques i les genotípiques queden modificades.
Tot i k sembli k les poblacions naturals no segueixen aquests supòsits, resultat k si k segueixen el mdoel de H-W.
Aparallament no aleatori-consanguinitat :
Els aparellaments selectius/classificats canviaran les freqüències genotípiques, però no alteraran els al·lels transmesos a la descendència, perquè seran els mateixos. És a dir, les freqüències al·lèliques no canviaran.
a. Aparallament classificat/selectiu positiu Æ Quan s’ aparellen mes alts amb altes que no pas els esperats per atzar. b. Aparallament selectiu negatiu Æ Quan s’ obtenen menys aparallaments d’ alts amb altes que no pas dels que s’ esperarien a l’ atzar.
Per tal de calcular el coeficient d’endogàmia o consanguinitat : f(individu) i F(població)
F= He-Ho/He = 2pq - Ho/2pq
He: heterozigots esperats Ho: heterozigots observats
Aquest índex mesura el grau d’endogàmia per una població, i pot prendre valors de 0-1. Si tenim una població amb un índex 1 , voldrà dir que tindrem consanguinitat màxima , que serien casos d’ autofecundació de plantes o casos d hermafroditisme (autozigosi). Si obtenim valors de 0 , voldrà dir que no hi ha endogàmia (sinó que hi ha exogàmia) i que per tant, s’ha produït un creuament a l’atzar. // L’ aïllament d’ 1 població farà varia la F.
[Actualment tots els conills d’ Austràlia són una població endogàmica, ja k tots han derivat d’ 1 grup petit de conills que es van portar d’ Espanya]
Coeficient d’ endogàmia de la descendència Consanguinitat entre germans 1/ 4 Consanguinitat entre tio i fiol 1/ Consanguinitat entre cosins germans 1/ Consanguinitat entre cosins germans amb 1 generació de diferència
1/
Consanguinitat entre cosins segons 1/
3. Canvis en les freqüències gèniques I. MIGRACIÓ. Models de migració. Flux gènic.
Una migració afecta les freqüències al·lèliques de la població.
Perquè una migració sigui considerada com a tal, haurà d’ anar acompanyada de la reproducció entre la població ja existent en el lloc i la població migradora.
Exemple : cas + senzill : 1 locus amb 2 al·lels :
Població 1 : f(A) = p1 f(e)= q
Població 2 : f(A) = p2 f(e)= q
Una porció (m) de la població 2 emigra cap a la població 1, de manera que considerem que la població 1 és la receptora dels immigrants, i per tant la població 1 seran els natius (1- m). Per tant :
Població mesclada : f(A) = p1(1-m) · p2·m f(e) = q1(1-m) · q2·m
És a dir, la freqüència al·lèlica de la població 1(receptora) pren un nou valor, ja que s’ ha de tenir en compte la freq. al·lèlic dels que entren i també la taxa de migració (m).
Abans de la migració
En humans casi cap poliploïdia és viable, xo en plantes si que n’ hi ha de viables.
Un heterozigot per inversió és aquell individu que té un cromosoma amb l’ ordre normal, i l’ altre en l’ ordre invertit.
Asa de inversión : ho fan els cromosomes per poder-se aparallar quan hi ha hagut una inversión, ja que sinó no poden aparallar-se.
Equilibri mutacional - Ex :
x Tenim 1 locus amb 2 al·lels, A1 i A2. x μ = taxa de mutaió Æ indica el canvi de l’ al·lel A1 a A2. x v = taxa de retromutació Æ indica el canvi de l’ al·lel A2 a l’ A1. x Suposant que no hi ha cap altra força evolutiva afectant el sistema : o μ · p = quantitat d’ al·lels que passen de A1 a A2. o v · p = quantitat d’ al·lels que passen de A2 a A1. x ∆q = μp – vq ∆q = μ (1-q) - vq ∆q = μ – q (μ + v) ∆q = 0
Mutació puntual (transicions i transversions)
Substitució de parells de bases en una seqüència d’ ADN
Possibilitat d’errors durant la síntesi d’ ADN causats per l’ADN polimerasa, o durant la reparació d’ ADN danyat per agents kímics,mutàgens o radioactius
Nous al·lels
Inversió cromosòmica
Salt amb volta (inversió) d’un segment cromosòmic
Talls en la doble hèlix per radiacions ionitzants.
Tendència al lligament perquè els heterozigots no recombinin per a la inversió. Duplicitat gènica Duplicació de talls curts de ADN
Aparellament desigual durant la meiosi.
Gens duplicats que poden mutar lliurement i potser guanyar una nova funció. Poliploidia (mutació de tot el genoma)
Addició d’un joc complet de cromosomes
Errors en meiosi o errors en mitosi.
Possiblement donin lloc a una nova espècie.
o Quan ∆q = 0 , significa que no hi ha variació , i per tant vol dir que s’ ha arribat a l’ equilibri mutacional , i per tant podrem calcular les freqüències al·lèliques en l’ equilibri :
A1 en equilibri : A2 en equilibri :
EX 12. (secció de problemes)
L’efecte de les mutacions en el canvi de freq. al·lèliques en les poblacions naturals s’observarà al cap de moltes generacions, és a dir, tenen un efecte a llarg termini. Per tant, en una única generació, l’efecte de la mutació serà imperceptible. Tot i així, si el tamany de la població és petit, l’ efecte de la mutació es veurà amb menys generacions.
En les mutacions recurrents es pot determinar la freqüència al·lèlica al cap de “t” generacions mitjançant la següent formula:
En les mutacions bidireccionals es pot determinar la freqüència al·lèlica al cap de “t” generacions mitjançant la següent formula: