



1
12
BIOLOGIA
CELULAR
3º GRADO BIOLOGIA
Raquel

























































































Prepara tus exámenes y mejora tus resultados gracias a la gran cantidad de recursos disponibles en Docsity

Gana puntos ayudando a otros estudiantes o consíguelos activando un Plan Premium

Prepara tus exámenes
Prepara tus exámenes y mejora tus resultados gracias a la gran cantidad de recursos disponibles en Docsity
Prepara tus exámenes con los documentos que comparten otros estudiantes como tú en Docsity
Encuentra los documentos específicos para los exámenes de tu universidad
Estudia con lecciones y exámenes resueltos basados en los programas académicos de las mejores universidades
Responde a preguntas de exámenes reales y pon a prueba tu preparación
Consigue puntos base para descargar
Gana puntos ayudando a otros estudiantes o consíguelos activando un Plan Premium
Comunidad
Pide ayuda a la comunidad y resuelve tus dudas de estudio
Ebooks gratuitos
Descarga nuestras guías gratuitas sobre técnicas de estudio, métodos para controlar la ansiedad y consejos para la tesis preparadas por los tutores de Docsity
Asignatura: Biología celular, Profesor: , Carrera: Biología, Universidad: UMU
Tipo: Apuntes
1 / 107

Esta página no es visible en la vista previa
¡No te pierdas las partes importantes!
Presentación seminario 1,25.
prácticas vale 1,5.
PROCARIOTAS
Las procariotas son las bacterias y las eucariotas son las células que forman parte de Protistas, plantas y animales. Las principales diferencias son:
El desarrollo del estudio de la célula se da con el desarrollo de la microscopia. Nuestro ojo es capaz de ver hasta 0’1 o 0’2 mm y las células presentan un tamaño de 5-10 μ. De manera que en función de la disminución del tamaño de las formas celulares tendremos 2 tipos de microscopio que nos permite estudiar células y componentes celulares.
Además de que las células son pequeñas, son transparentes, de manera que tienen poco contraste (pequeñas, acuosas y tienen átomos ligeros) y no se pueden observar fácilmente. Para poder ser estudiadas por un microscopio podemos modificar el contraste de la muestra (tinciones o podemos modificar la física del microscopio. En función de estos dos factores presentamos distintos tipos de microscopia.
LUZ BLANCA λ = 550nm AN= n · senα AN ACEITE= 1, 5
Cuanto mayor es la apertura numérica (AN) menor es el límite de resolución. Cuanto más cerca esté la muestra del objetivo menor será el límite de resolución. Con aceite de inmersión, el límite de resolución aumenta hasta 1000 veces con respecto al ojo humano. El microscopio de campo claro necesita aumentar el contraste de la muestra, para ello se tiñe la muestra de manera que para suministrar coloración se trabaja con material muerto, de manera que esta microscopía no permite material vivo salvo que tenga pigmentos naturales.
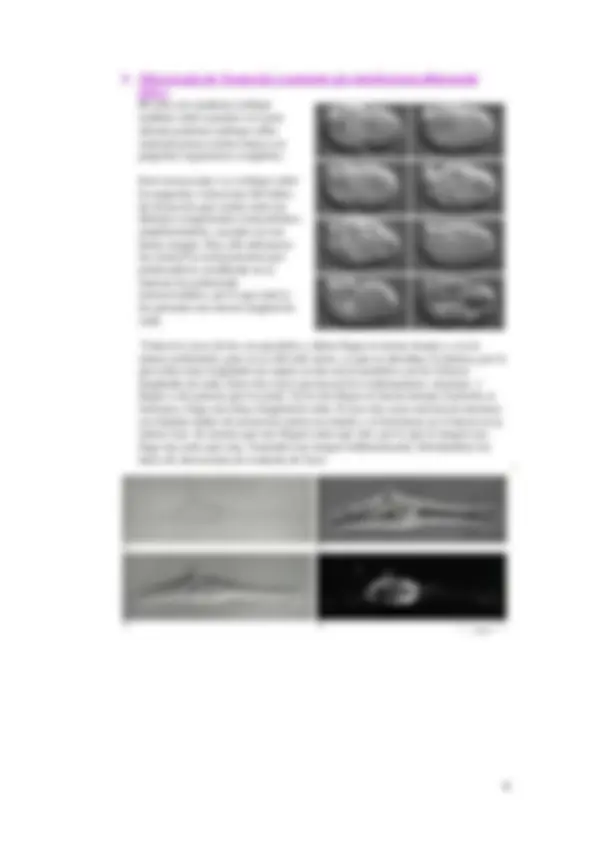
El microscopio de campo oscuro presenta una ventaja y es que permite trabajar con material vivo, que no tiene por qué estar coloreado. Lo que se hace es variar la física de la luz de manera que nos ofrece en el material vivo unos contornos brillantes con un fondo oscuro, ya que entre el condensador y la fuente de luz se pone un cilindro que bloquea toda la parte central de la luz, solo llegando la parte periférica de la luz haciendo que toda la luz llegue inclinada hacia la muestra y toda aquella luz que no es dispersada por la muestra no la vemos y vemos la dispersada por ello vemos brillo sobre un campo oscuro. En este caso no se permite estudiar con mucho detalle, es decir, no tiene buen límite de resolución, pues su apertura numérica no pasa de 1.
El microscopio de contraste de fase es muy usado, y nos permite estudiar material vivo debido a su física, ya que se introduce un condensador de anillo y un plato de fase en el microscopio. El condensador solo permite que pase un cilindro de luz hueco en el centro que luego hace que una vez que la luz llega sobre la muestra se produzca un pequeño desfase en la luz (de un cuarto) que llega a través de la célula produciendo una refracción muy pequeña, pero al colocar el plato de fase hace que la longitud de onda que va desfasada la vuelve a retrasar (otro cuarto) de manera que se retrasa la mitad la longitud de onda y esta mitad si se puede recoger y ver. Su poder de resolución es bajo pero nos permite distinguir el contorno celular, el núcleo, etc.
En epifluorescencia el haz de luz llega por arriba incide sobre la muestra y vuelve de nuevo hacia arriba. Estos microscopios se basan en la fluorescencia, que es la capacidad de ciertas moléculas de absorber luz de una determinada longitud de onda y emitir luz a mayor longitud de onda durante un periodo de tiempo. Esto produce la emisión de energía que provoca esta fluorescencia por acción de los fluorocromos. La parte del fluorocromo que produce esta fluorescencia es el fluoroforo. Cada fluorocromo tiene un espectro de excitación con un máximo y va a tener un espectro de emisión de mayor longitud de onda con un máximo también. Estos espectros pueden solaparse en parte y pueden darnos imágenes que no se adecuen a la realidad. En este tipo de microscopio se cambian las fuentes de iluminación puesto que tenemos una luz ultravioleta y se parte con una longitud de onda baja para emitir longitudes mayores y puedan ser visibles, por eso se usa la luz ultravioleta (340-500 nm). Estos microscopios presentan una serie de filtros que no deben porque estar en el microscopio sino que se añaden a un microscopio cualquiera al que se le añade la fuente de luz ultravioleta y quita la luz. Hay un filtro que será el de excitación al que le llegará una luz y se elige la longitud de onda que queremos que atraviese ese filtro y que excite al fluorocromo. Después tenemos los
filtros de barrera. Estos eliminan todos los fluorocromos que se puedan solapar y pueden ser de 3 tipos: Filtro de paso de banda , que puede pasar un rango más o menos estrecho de longitudes de onda. Puede pasar la longitud de onda de una franja. Filtro de paso corto , donde pasan todas las longitudes que sean menores de 600 nm por ejemplo, y las que no lo sean, no pasan. Filtro de paso largo donde pasan todas las longitudes de onda que sean mayores a X (por ejemplo, 600nm), mientras que las menores no.
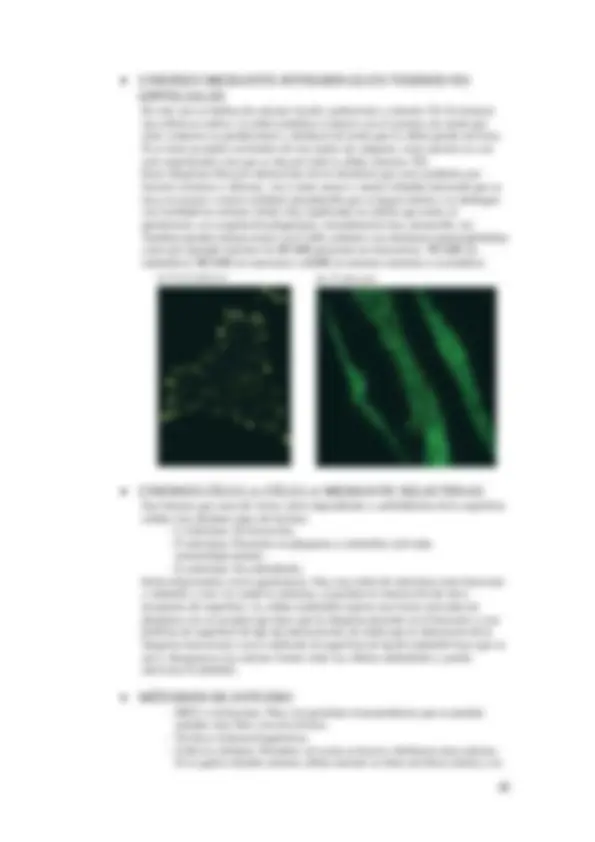
En esta microscopia se pueden estudiar tanto material vivo como material muerto. Se puede usar para determinar una muestra concreta, para la presencia de una enzima, de ADN, proteínas, etc. Además se pueden utilizar distintos fluorocromos en la misma muestra, generalmente se trabaja con equipos rutinarios con verde, rojo y azul. El problema es que las muestras que se colorean no se saben con exactitud donde se encuentran cada estructura, por lo que se deben comparar con otras imágenes de la misma muestra con distinto microscopio para ver de qué estructura se trata.
A la hora de trabajar con microscopia de fluorescencia debemos saber que:
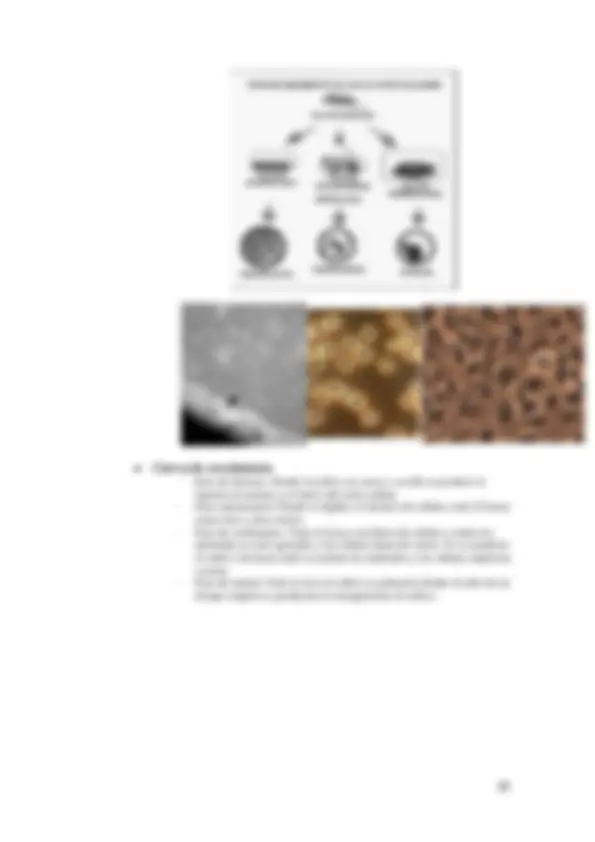
Se utiliza para estudiar la ultraestructura. Tenemos un microscopio con alto poder de resolución (1 nm). Esta microscopía es similar al microscopio óptico, se trabaja a alto vacio. Esto hace que los electrones se aceleren a un ánodo perforado pasando los electrones por el vacio por lentes electromagnéticas hasta llegar a la muestra y a través de unas lentes llega a una cámara digital.
Al chocar los electrones sobre la muestra, estos se dispersan en todas las direcciones. En esta microscopía se recogen los electrones que no se dispersan por lo que en la imagen se verá en tonos negros, blancos y grises. Se trata de muestras sin nada de agua, estabilizadas y que estén contrastadas, pues todos los componentes celulares tienen muy baja densidad y eso no ofrece resistencia a los electrones. Se usan técnicas de tinción que nos permitan identificar estructuras y que sean muy electrodensos, siendo capaces de ser reflejados los electrones viendo así estructuras que sin contrastar no veríamos. Se obtendría una imagen similar a las de campo claro pero sin color. Además, para favorecer la penetración y transmisión de estos electrones la muestra debe ser muy fina. Se aumenta la imagen 200.000 veces.
En este caso no estudiamos los electrones transmitidos, sino los dispersados. Esta microscopía nos permite estudiar la superficie, y su resolución es menor a la anterior (10nm), trabajando a 10.000 o 20.000 aumentos.
Una vez que los electrones inciden sobre la muestra se recogen los electrones dispersados. En este caso las muestras se metalizan (material electrondenso) para hacer que los electrones se dispersen, pudiendo estudiar la superficie.
Hay distintos tipos de fijadores:
Lo que se usa más que fijadores simples son mezclas de fijadores, normalmente se utiliza:
dificulta el procesado por lo que se elimina tras su acción y para ello lo sumergimos en etanol de 70.
Inclusión : Una vez fijada se inicia la inclusión que sirve para hacer un bloque que permita hacer cortes de la muestra. Se utiliza para hacer el bloque materiales no hidrosolubles, como parafina o resinas, por lo que la muestra se queda sin agua. Primero se deshidrata con pasos progresivos de etanol al 70% (50-70-90-96-absoluto) 1- 2h cada uno. Tras la eliminación del agua se hace el aclaramiento pues la sustancia para realizar el bloque no es miscible en la muestra, por lo que se usa Xilol que si lo es. Tras ello se hace la impregnación , se impregna todo con parafina liquida (60oC) y una vez que lo tenemos impregnado se incorpora en un molde, se le añade parafina y queda la muestra secuestrada en el molde, quedándonos un bloque de parafina.
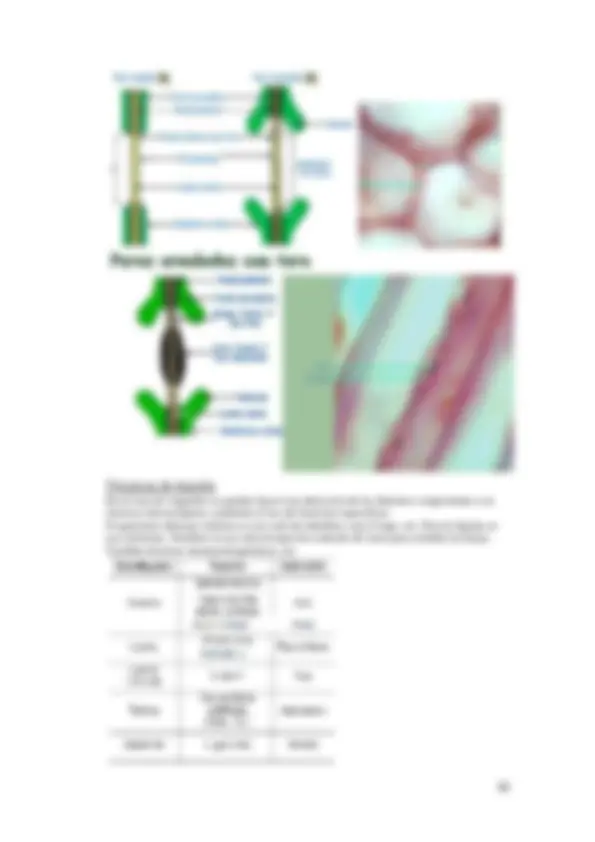
Corte : Este corte puede hacerse de distinta forma, puede ser a mano alzada si se tiene una estructura ya endurecida como se hace en histología vegetal. En el caso de tejido con el que se usa microtomo, primero hacemos un devastado del bloque (se dibuja como un trapecio alrededor de la muestra que será lo que se corta) y mediante un microtomo de parafina cortaremos la muestra en tamaños de 3 a 10 μm y una vez que tenemos los cortes estos van saliendo en fila y se van colocando en un portaobjetos. Para ello nos ayudamos de poli-L-lisina o albumina. Se pueden hacer cortes individuales o bien seriadas (se usa mucho para localizar en la misma célula dos reacciones o dos componentes, etc.). También hay criomicrotomos que se usan para hacer cortes por congelación, se hacen cortes de 5 a 30 μm y funcionan los microtomos igual que los anteriores.
Tinción : Una vez que el colorante ha reaccionado con el material biológico, el resultado es que ese colorante quede fijado en el material biológico. Los colorantes son soluciones acuosas de modo que no se puede impregnar con el material hasta que no se produzca la desparafinacion, se hidrate la muestra y ya por último se realice la tinción. Tras esto se realiza el montaje mediante el uso de un material hidrosoluble como la glicerina pero es hidrosoluble por lo que no se conserva por mucho tiempo, también se puede mantener por material no hidrosoluble, como es el DPX que nos lo puede mantener por más tiempo, pero para este caso se debe deshidratar de nuevo. Este montaje se realiza para mantener la muestra (en el caso de DPX) y además como su índice de refracción es mayor que el del aire nos permite ver imágenes con mayor índice de refracción.
mas típicas que se ponen de manifiesto son esterasas, fosfatasas, peroxidasas y diaforasas. Nos debemos asegurar de que la reacción es específica. Se usara contraste de fondos para identificar el fondo de nuestra muestra. Muchas de las enzimas que identifica solo presentan una localización, por lo que no solo se usa para poner de manifiesto una enzima, sino que se utiliza para localizar una estructura como son los lisosomas porque la mayoría de las enzimas estudiadas con esta técnica están allí. Hay otras enzimas que están solo en el Golgi, otras en el RE, etc.
al anticuerpo. La inmunocitoquímica puede ser directa cuando se une el anticuerpo con el marcador o bien indirecta cuando se usa primero un anticuerpo frente al antígeno y después un anticuerpo secundario frente al anticuerpo primario y este secundario es el que va marcado, amplificando así la señal. Como marcador se suele usar: Una enzima, para MO, como puede ser: Peroxidasa : La peroxidasa usa un sustrato que produce un precipitado de color marrón oscuro pero el problema que presenta es que no se sabe si la reacción es debida a la que va unida al anticuerpo o bien a la peroxidasa de dentro de la célula. La peroxidasa tiene un punto débil y es que se inhibe de forma irreversible por el sustrato, por lo que si la peroxidasa, una vez que se desparafina y se hidrata, se somete a alta concentración de peróxido de hidrogeno se inhibe, para así eliminar el exceso de peroxidasa quedando solo el que está unida al sustrato. Además hay que tener en cuenta que existe otro inhibidor de la peroxidasa que es la azida sódica. Fosfatasa alcalina : No se inhibe por sustrato, pero se hace una inhibición de la fosfatasa endógena. Se puede añadir sustrato de forma continuada con la fosfatasa alcalina no como en la peroxidasa. La señal se puede amplificar con métodos como: PAP : Donde tenemos un anticuerpo primario que reconoce al anticuerpo secundario, y después esta la estructura PAP que presenta un anticuerpo terciario que presenta peroxidasa, revelándose al igual que antes. ABC : En este caso el anticuerpo secundario va marcado con biotina, después se pone avidina en el sistema ABC, uniéndose proteínas de forma específica a biotina que lleva unida peroxidasa.
Se pueden utilizar bien con peroxidasas o bien con fluorocromos, de manera que se puede visualizar mediante microscopia de fluorescencia o de barrido o citometría de flujo. Si se quiere usar MET lo que se usara para marcar al anticuerpo partículas de oro coloidal. También se puede realizar una inmunocitoquímica simple o bien múltiple (detectar varios antígenos al mismo tiempo). Para ello debemos tener una precaución, se deben tener anticuerpos generados en especies distintas para que no haya reacción cruzada. Para identificar varios antígenos de una misma especie usando diferentes sistemas de revelado, pero el antígeno debe expresarse en amplia cantidad.