BIOLOGÍA CELULAR
Tema 3.
Mecanismos de regulación y coordinación del tráfico vesicular
Regulación y coordinación del tráfico vesicular
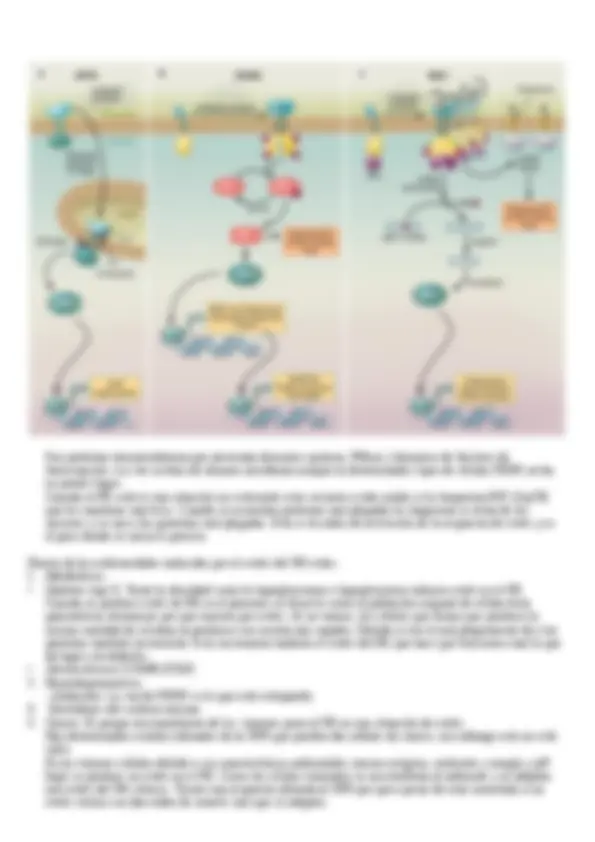
Las GTPasas monoméricas tiene un papel muy importante de las distintas etapas del tráfico intracelular, lo
regulan y lo coordinan. No solo se encargan del ensamblaje de las cubiertas sino también de su movimiento, de
su fusión con los compartimentos aceptores etc. En mamíferos hay diferentes tipos de GTPasas que funcionan
en dos estadios distintos de configuración unidad a GDP (inactivas) unidas a GTP (activas).
Dentro de las GTPasas hay diferentes familias. El tráfico intracelular está controlado por Rho, Rab y Sar1/Arf.
1. Rho. Esta asociadas también a los cambios en el citoesqueleto de actina. Hay distintos tipos de endocitosis
que necesitan de la intervención de este citoesqueleto y de la reestructuración del mismo para producir los
efectos adecuados.
En el caso del tráfico intracelular actúan. Rho A, Rac 1 y Cdc42
2. Rab. También son fundamentales en los procesos de tráfico intracelular. Controlan todas las etapas del
tráfico vesicular. Participan en el brote de las vesículas, en la pérdida de su revestimiento, en su movimiento
a través del citoplasma y el los procesos de fusión de las vesículas con sus compartimentos
correspondientes. En cada uno de los compartimentos y en cada una de las vesículas o carriers encontramos
distintos tipos de proteínas Rab. La inhibición de estas proteínas tiene efectos muy drásticos en las células.
3. Sar1/Arf. Sar1 es al que se encarga de reclutar el revestimiento de las vesículas COP II y así un montón más
que están en apuntes anteriores claramente detalladas.
Cosas que no se saben
• Cuantos tipos de COP II salen del RER ni si éstas están especializadas en la carga que transportan.
• Cuántos tipos de COP I se generan en el Golgi y en el ERGIC y como llegan estas vesículas a su
compartimento de destino.
• Cuánto tarda el desensamblaje de los revestimiento de las vesículas y cual es la orden in vivo para que esto
se produzca.
Alteraciones relevantes para la salud humana
La maquinaria completa necesaria para que se produzca un tráfico de membranas en una célula es
aproximadamente de 2000 proteínas. Cualquiera de estas 2000 proteínas puede ser una diana de marcaje o
terapéutica.
Hay múltiples situaciones en las que un mal funcionamiento puede tener consecuencias graves para la célula.
1. Salida del RER hasta el ERGIC.
2. Desde el ERGIC al retículo
3. Modificaciones del complejo de Golgi. Tanto en su funcionamiento como en las proteínas.
4. Salida desde la red trans-Golgi hacia los diferentes destinos.
5. Procesos de endocitosis.
Esto quiere decir que hay múltiples puntos en los que la célula tiene que tener un control muy exhaustivo para
que no haya un problema patológico
Salida del RER hasta el ERGIC
Problemas que puede haber por esta circunstancia.
1. Retención de la carga. La carga es retenida en el RE y degradada por el proteasoma por que las proteínas
no están bien sintetizadas. Esto depende del ERAD y suele ocurrir cuando las proteínas no consiguen
plegarse después de varios ciclos.
En la actualidad hay 67 enfermedades humanas que se asocian con la vía ERAD. Es decir que hay
determinadas proteínas que se degradan por esta vía y que no alcanzan su destino correcto.
La que está mejor estudiada es la fibrosis quística.
Existe un canal transmembrana que se llama CFTR que es un transportador ABC. Lo que hace este
transportador, en condiciones normales, es liberar Cl al exterior, en contra de gradiente. Estos iones de Cl
son fundamentales para la hidratación en determinados tipos de epitelios. Cuando este canal no funciona de
manera correcta, la ausencia de Cl en la superficie hace que se produzca un mucus de mucha densidad.