



Biosíntesis de lípidos de
membrana, esteroides,
isoprenoides y eicosanoides










Prepara tus exámenes y mejora tus resultados gracias a la gran cantidad de recursos disponibles en Docsity

Gana puntos ayudando a otros estudiantes o consíguelos activando un Plan Premium

Prepara tus exámenes
Prepara tus exámenes y mejora tus resultados gracias a la gran cantidad de recursos disponibles en Docsity
Prepara tus exámenes con los documentos que comparten otros estudiantes como tú en Docsity
Encuentra los documentos específicos para los exámenes de tu universidad
Estudia con lecciones y exámenes resueltos basados en los programas académicos de las mejores universidades
Responde a preguntas de exámenes reales y pon a prueba tu preparación
Consigue puntos base para descargar
Gana puntos ayudando a otros estudiantes o consíguelos activando un Plan Premium
Comunidad
Pide ayuda a la comunidad y resuelve tus dudas de estudio
Ebooks gratuitos
Descarga nuestras guías gratuitas sobre técnicas de estudio, métodos para controlar la ansiedad y consejos para la tesis preparadas por los tutores de Docsity
Este trabajo aborda la biosíntesis de glicerofosfolípidos, que son componentes fundamentales de la membrana; los esfingolípidos, que se encuentran fundamentalmente en el tejido nervioso de los animales, sin embargo, también se encuentran en las membranas de las células vegetales y en los eucariotas inferiores, como levaduras; los esteroides y otros componentes isoprenoides, que actúan como componentes de la membrana, hormonas y vitaminas, y por ultimo por eicosanoides.
Tipo: Monografías, Ensayos
1 / 19

Esta página no es visible en la vista previa
¡No te pierdas las partes importantes!
Este trabajo aborda la biosíntesis de glicerofosfolípidos, que son componentes fundamentales de la membrana; los esfingolípidos, que se encuentran fundamentalmente en el tejido nervioso de los animales, sin embargo, también se encuentran en las membranas de las células vegetales y en los eucariotas inferiores, como levaduras; los esteroides y otros componentes isoprenoides, que actúan como componentes de la membrana, hormonas y vitaminas, y por ultimo por eicosanoides, un tipo de reguladores biológicos que se sintetizan a partir del ácido araquidónico.
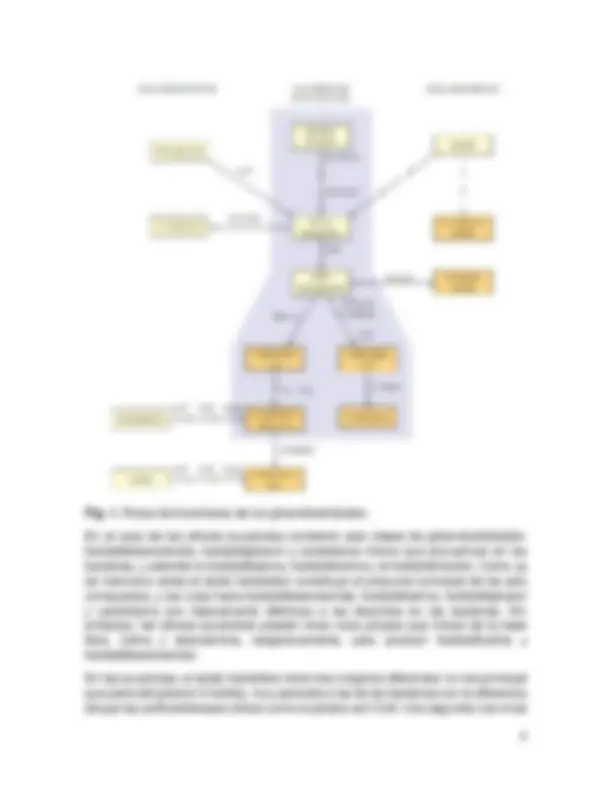
Se empezará mencionando la biosíntesis de algunos glicerofosfolípidos del reino de los procariotas, debido a la simplicidad de sus sistemas biológicos, además de que la ruta que estos siguen en las procariotas es igual que en las eucariotas. Sin embargo, también se mencionan otras rutas propias de las eucariotas. Utilizaremos como ejemplo a E. coli, ya que es el microorganismo mas estudiado, sus membranas contienen tres fosfolípidos en cantidades significativas: fosfatidiletanolamida, fosfatidilglicerol y cardiolipina. Como se muestra en la Fig. 1, el ácido fosfatídico constituye un punto de ramificación entre la síntesis de triacilgliceroles y la de los fosfolípidos. El cofactor energético para la biosíntesis de los fosfolípidos es la citidina trifosfato (CTP). El ácido fosfatídico se sintetiza por medio de dos acilaciones sucesivas del glicerol- 3 - fostato. En bacterias, los grupos acilo se transportan por la proteína transportadora del acilo. Se involucran en el proceso dos enzimas aciltransferasa diferentes, que ponen de manifiesto en parte sus diferencias de especificidad por los acil ACP: aproximadamente el 90% de los grupos acilo esterificados en la posición 1 están saturados, por otra parte, el 90% de los de la posición 2 están insaturados. Enseguida el ácido fosfatídico se activa metabólicamente al reaccionar con CTP. Esta reacción da como producto CDP-diacilglicerol, que contiene un enlace anhidrido que une los fosfatos de los ácidos citidílico y fosfatídico. El CDP- diacilglicerol está ya activado para la síntesis de los grupos de cabeza polares. Entonces el CMP se intercambia por serina, para dar fosfatidilserina, que inmediatamente sufre una descarboxilación a fosfatidiletanolamida. Otra ruta transfiere el glicerol- 3 - fosfato a acido fosfatídico, seguido por una reacción fosfatasa para dar fosfatidilglicerol. Al reaccionar con otro mol de fosfatidilglicerol se obtiene difosfatidilglicerol o también llamado cardiolipina.
Fig. 1. Rutas de biosíntesis de los glicerofosfolípidos. En el caso de las células eucariotas contienen seis clases de glicerofosfolípidos: fosfatidiletanolamida, fosfatidilglicerol y cardiolipina mismo que encuentran en las bacterias, y además la fosfatidilserina, fosfatidilcolina y el fosfatidilinositol. Como ya se menciono antes el ácido fosfatídico constituye el precursor principal de los seis compuestos, y las rutas hacia fosfatidiletanolamida, fosfatidilserina, fosfatidilglicerol y cardiolipina son básicamente idénticas a las descritas en las bacterias. Sin embargo, las células eucariotas poseen otras rutas propias que inician de la base libre, colina y etanolamina, respectivamente, para producir fosfatidilcolina y fosfatidiletanolamida. En las eucariotas, el ácido fosfatídico tiene tres orígenes diferentes: la ruta principal que parte del glicerol- 3 - fosfato, muy parecida a las de las bacterias con la diferencia de que las aciltransferasas utilizan como sustratos acil-CoA. Una segunda ruta inicia
La ruta de salvamento para la fosfatidiletanolamina utiliza las mismas reacciones, partiendo de la etanolamina en vez de la colina. La misma enzima realiza la fosforilación de la colina y de la etanolamina, sin embargo, las reacciones posteriores las efectúan enzimas distintas en las dos rutas. La ruta alternativa para fosfatidiletanolamida y fosfatidilcolina empieza con la conversión de la fosfatidilserina en fosfatidiletanolamida, catalizada por una de dos enzimas diferentes. La fosfatidilserina descarboxilasa es una enzima mitocondrial que descarboxila la fosfatidilserina a fosfatidiletanolamina. La otra enzima es una transferasa activada por el calcio, la fosfatidiletanolamina serinatransferasa, que intercambia la etanolamina libre por la porción serina de la fosfatidilserina, produciendo fosfatidiletanolamina y serina. Luego, la fosfatidiletanolamina sufre tres metilaciones sucesivas, catalizadas probablemente todas ellas por la misma enzima, para dar como resultado fosfatidilcolina (Fig. 3). Fig. 3. Ruta alternativa para fosfatidiletanolamida y fosfatidilcolina.
Los esfingolípidos incluyen la ceramida, la esfingomielina y una familia de esfingolípidos que contienen hidratos de carbono denominados glucoesfingolípidos neutros y ácidos; estas últimas sustancias incluyen los cerebrósidos y los gangliósidos. La ceramida actúa como precursor tanto de la esfingomielina como de los glucoesfingolípidos. En los animales, la ruta de la ceramida inicia con la síntesis de un derivado de la esfingosina, la esfinganina, a partir de palmitoil-CoA y serina como se puede
apreciar en la Fig. 4. Luego tras la reducción del grupo ceto resultante, el grupo amino de la esfinganina se acila y produce una ceramida. La unidad esfinganina de este compuesto se desatura a continuación para dar una ceramida con una base esfingosina. La transferencia de una unidad fosfocolina procedente de la fosfatidilcolina da esfingomielina y diacilglicerol. Fig. 4. Biosíntesis de esfingolípidos.
ácidos biliares, las vitaminas liposolubles, entre otros. Gran parte del estudio sobre los isoprenoides se centra en un único compuesto esteroideo, el colesterol, este lípido es un componente principal de las membranas de las células animales en donde participa en la modulación de la fluidez de la membrana. En los animales también actúa como precursor de todas las hormonas esteroideas, de la vitamina D, y de los ácidos biliares, que facilitan la digestión de las grasas. La biosíntesis del colesterol se considera en tres fases: Fase 1: formación del mevalonato. La fase 1 inicia con la condensación de dos moléculas de acetil-CoA para dar acetoacetil-CoA. La acetoacetil-CoA reacciona con una tercera molécula de acetil- CoA para dar 3-hidroxi- 3 - metilglutaril-CoA por medio de la HMG-CoA reductasa que cataliza la reducción de cuatro electrones, dependiente de NADPH, que transforma la HMG-CoA en mevalonato (Fig. 6). Fig. 6. Formación de mevalonato. Fase 2: síntesis de escualeno a partir de mevalonato. En primer lugar, se activa el mevalonato mediante tres fosforilaciones sucesivas. La tercera fosforilación, en la posición 3, sienta las bases para una descarboxilación a través de una eliminación trans para dar isopentenil pirofosfato. Una molécula de esta sustancia se isomeriza a dimetilalil pirofosfato (Fig. 7).
Fig. 7. Conversión del mevalonato en isopentenil pirofosfato y dimetilalil pirofosfato. Entonces el dimetilalil pirofosfato reacciona con otra molécula de isopentenil pirofosfato para dar geranil pirofosfato y posteriormente otra molécula de isopentenil pirofosfato reacciona con este producto para dar farnesil pirofosfato (Fig. 8).
Fig. 9. Conversión del farnesil pirofosfato en escualeno. Fase 3: ciclación del escualeno a lanosterol y su conversión en colesterol. La ciclación del escualeno a lanosterol y la conversión del lanosterol en colesterol se muestra en la Fig. 10. La formación de lanosterol, que tiene el núcleo esterol de cuatro anillos, se produce en dos pasos. En primer lugar, una oxidasa de función mixta introduce una función epóxido en los carbonos 2 y 3. La protonación de este grupo funcional inicia una serie de cambios trans 1,2 de los grupos metilo y los iones hidruro, para producir el lanosterol. A continuación, se producen una serie de unas 20 reacciones, en las que intervienen reducciones de dobles enlaces y tres desmetilaciones; se elimina un grupo metilo de C-14 y dos de C-4. El penúltimo producto es el 7 - deshidrocolesterol, que sufre una reducción final para dar colesterol.
Fig. 10. Conversión del escualeno en colesterol.
Fig. 11. Rutas de biosíntesis que van desde la pregnenolona a otras hormonas esteroideas.
Los terpenos se biosintetizan en última instancia a partir del isopentenil pirofosfato (C5) y del dimetilalil pirofosfato (C5). Cuando éstos se combinan para producir el geranil pirofosfato (C10), los terpenos formados se denominan monoterpenos. Cuando se forma un compuesto a partir de 1 mol de farnesil pirofosfato (C15), el producto se denomina un sesquiterpeno. Los triterpenos (C30) se forman a partir de 2 moles de farnesil pirofosfato. El geranilgeranil pirofosfato (C20) produce o bien diterpenos (C20) o bien tetraterpenos (C40).
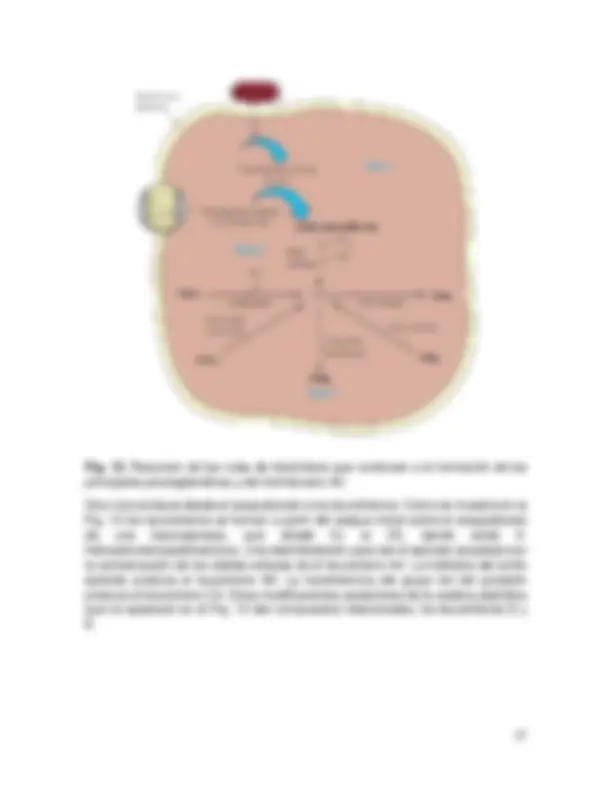
Los eicosanoides se caracterizan por sus potentes propiedades fisiológicas, sus bajas concentraciones en los tejidos, su rápido recambio metabólico y su origen metabólico común. Los más importantes de estos compuestos son las prostaglandinas; otras sustancias de esta clase son los tromboxanos y los leucotrienos. Las rutas de síntesis de las prostaglandinas y tromboxano se producen en tres fases diferentes (Fig. 12): (1) liberación del ácido araquidónico a partir de los fosfolípidos de la membrana, (2) oxigenación del araquidonato para producir PGH2, un endoperóxido de prostaglandina que actúa como precursor de otras prostaglandinas, y (3) en función de las enzimas existentes en una célula, la conversión de la PGH en otras prostaglandinas o en Tromboxano A2. La liberación de ácido araquidónico en la fase 1 se produce como consecuencia de estímulos específicos de los tejidos por hormonas como la bradiquinina o la adrenalina, o por proteasas como la trombina. Puede producirse una liberación patológica si se alteran las membranas. La liberación comporta evidentemente la acción de una fosfolipasa A2 específica sobre la fosfatidilcolina o la fosfatidiletanolamina, dando araquidonato, o la acción de una fosfolipasa C sobre el fosfatidilinositol dando un diacilglicerol, que a su vez sufre una rotura para dar araquidonato libre. En la fase 2, el araquidonato libre sufre la acción de la PGH sintasa, una enzima bifuncional con dos actividades en una única cadena polipeptídica que contiene hemo. La primera, una ciclooxigenasa, introduce dos moléculas de O2, una para formar el anillo y otra para formar un grupo hidroperoxido en C-15. La segunda actividad comporta una reducción de dos electrones del peróxido, para dar PGH2, con un grupo hidroxilo en C- 15. En la fase 3, una serie de enzimas específicas convierten la PGH2 en otras prostaglandinas y en el tromboxano A2.
Fig. 13. Biosíntesis de los leucotrienos.