¡Descarga Bloque 1 Biología Celular 1ºCurso Grado Biología UAM y más Apuntes en PDF de Biología Celular solo en Docsity!
BLOQUE 1. SUPERFICIE CELULAR
TEMA 2. MEMBRANAS CELULARES
Membrana plasmática : estructura que rodea a la célula, delimitándola, y encierra a sus componentes. Desarrolla funciones vitales para la célula como la interacción con el medio y con otras células, la protección y la especialización celular a través de estructuras externas (microvellosidades). Componentes: la membrana plasmática es una estructura delgada (7,5 nm) y que posee una composición entre lípidos (40%), proteínas (60%) y azúcares (10% asociados a proteínas y lípidos). LÍPIDOS: sustancias anfipáticas. Los más importantes son: Fosfoglicéridos: fosfatidilcolina, fosfatidilserina, fosfatidiletanolamina (aminoalcoholes) y fosfatidilinositol (inositol). Esfingolípidos: están basados en la esfingosina. Tipos: Esfingomielina (ceramida+fosfato+colina) Glicolípidos (ceramida+azúcar). En lugar del fosfato y del aminoalcohol, se une al grupo OH libre un azúcar. Dependiendo de la complejidad del mismo, se distinguen: o Cerebrósidos o Gangliósidos (azúcares complejos ramificados) Ambas son moléculas poseen una geometría cilíndrica.
Esteroles: se derivan de otro tipo de lípido. El principal esterol es el colesterol, que se trata de una molécula más pequeña con geometría plana. BICAPA LIPÍDICA La organización de una membrana plasmática tiende a la menor cantidad de energía libre. Los lípidos anfipáticos, por tanto, pueden formar dos estructuras: Micela: se forma debido a la geometría cónica de las moléculas que la forman. Bicapa: se forma debido a la geometría cilíndrica de los lípidos que la forman, cuya formación es la de menor energía libre. *Existe una capa de 1,5 nm donde se encuentran las colas hidrofóbicas en la que el agua no penetra. Características de la membrana Fluidez: los lípidos de la membrana se pueden mover dentro de la bicapa. Este movimiento se produce debido a la temperatura y el medio acuoso donde se
PROTEÍNAS: corresponden al 60% en peso de la membrana plasmática. (Las proteínas son moléculas más grandes que los lípidos, por lo que poseen mayor peso) Clasificación: EXTRÍNSECAS: no forman parte de la estructura ni unidas covalentemente a otros componentes, sino que se encuentran unidas por interacciones débiles (puentes de H, fuerzas de VdW) INTRÍNSECAS: forman parte de la membrana y permanecen en su interior o unidas covalentemente a algún componente de la misma. Tipos: Integrales: están ancladas a la bicapa de la membrana. Poseen una serie de dominios hidrofóbicos que atraviesan una o varias veces la membrana. También poseen dominios globulares hidrofílicos que permanecen en contacto con el medio extra o intracelular. Pueden poseer un solo dominio o varios, que atraviesan la membrana. Existen proteínas integrales no transmembrana, que están asociadas a una sóla hemimembrana. Los dominios transmembrana tienden a tener una configuración en α-hélice ó β-lámina. Estos dominios están constituidos por AA apolares. FORMACIONES: Barril-beta: formado por β-láminas, genera un poro en la membrana a través del cual se genera un canal acuoso que permite la interacción con el medio. Estos poros son fijos. Poros α-hélice: se deben asociar varias α-hélice para formar un poro, que se pueden abrir o cerrar.
Periféricas: unidas covalentemente a lípidos de la membrana o a otras proteínas de membrana. Aparecen en ambas hemimembranas. MOVIMIENTOS: las proteínas no aportan fluidez a la membrana, sólo siguen el movimiento de la bicapa debido a las asociaciones de la parte hidrofóbica con las colas hidrofóbicas de los lípidos. Tipos: Desplazamiento lateral en el plano de la membrana. Rotación ASIMETRÍA: existe una asimetría respecto de la ubicación de las proteínas en la membrana plasmática que hacen que se produzcan fenómenos tales como la bomba Na y K. (Las proteínas están colocadas de una sola forma, sin movimiento de Flip-flop) HETEROGENEIDAD: se forman agrupaciones de proteínas en las balsas lipídicas. Las proteínas con la parte hidrofóbica más larga se encuentran en balsas lipídicas, zonas más gruesas de la bicapa, donde la proteína se adhiere de forma más estable. De esta forma la parte hidrofóbica no entra en contacto con los medios extra e intracelular. Se agrupan en estas balsas por cuestiones funcionales o estructurales. FUNCIONES DE LAS PROTEÍNAS Transporte Reconocimiento celular Adhesión celular Enzimática- CATÁLISIS AZÚCARES Glicocáliz (Glicocálix): se trata de una estructura de la membrana plasmática con aspecto fibroso o granuloso localizada en la parte externa de la membrana. Se une a los lípidos y proteínas de la hemimembrana externa. Esta glucosilación sólo se produce en la parte externa. Las moléculas glucosiladas más importantes son: Lípidos glicosilados (glicolípidos): derivados de la esfingosina. Cerebrósidos y gangliósidos. Se produce en el aparato de Golgi.
hidrofílica dentro del poro, y la parte hidrofóbica en contacto con la bicapa. Se trata de un transporte de alta velocidad. Permeasas y bombas: permiten el transporte de manera más lenta, ya que no abren un poro acuoso, sino que interaccionen de manera más estrecha con la molécula transportada. En el transporte sufren un cambio conformacional. Se trata entonces, de un transporte específico. Gradiente: Gradiente químico o de concentración: las moléculas generan un gradiente a favor de las concentraciones menores, tendiendo a igualar concentraciones en ambos ambientes. Gradiente eléctrico: las moléculas con carga establecen un gradiente para igualar la carga en ambos ambientes alrededor de la membrana. Puede suceder que el gradiente químico y el eléctrico se contrarresten, surgiendo así el gradiente electroquímico. Este gradiente puede considerarse el sistema de regulación de entrada y salida de moléculas en la célula. *Normalmente en la célula existen mayor número de cargas negativas dentro y mayor número de cargas positivas fuera. *Normalmente existe una mayor concentración extracelular de Na+, Ca2+^ y Cl-; y una mayor concentración intracelular de K+^ y glucosa. Este tipo de gradientes determina el tipo de transporte de la célula. De ello se extraen tres tipos de transporte: Difusión simple: las moléculas atraviesan la bicapa con total libertad y siempre se produce a favor del gradiente de concentración Transporte pasivo o difusión facilitada: siempre se produce a favor del gradiente de concentración. No requiere un gasto energético, ya que se realiza a través de canales protéicos y permeasas. Transporte activo: se produce el transporte de una sustancia en contra de su gradiente de concentración, realizado por bombas.
TRANSPORTE PASIVO
Canales: cada canal es específico de la molécula que van a transportar. Están formados por dominios transmembrana de α- hélice con canales hidrofílicos. Tipos: Canales iónicos: Canales dependientes de ligando: estos canales sólo se estimulan ante la llegada de alguna sustancia química. (Acetilcolina) Ej.: canal de Na+^ dependiente de acetilcolina. Canales dependientes de voltaje: se estimulan porque detectan un cambio en la polaridad de la membrana. Su estimulante, por tanto, es una diferencia de potencial. Ej.: canal de Na+^ y de Ca2+^ dependientes de voltaje.
Acuaporinas: se trata de canales normalmente abiertos. Dejan pasar agua a favor del gradiente de concentración. Pueden cerrarse ante determinados estímulos (apertura regulada). Son proteínas del tipo α-hélice. ***** Los estímulos que abren los canales tienen una duración determinada regulada por otros estímulos que los cierran. En el caso de los canales dependientes de ligando, existen sustancias (enzimas) que destruyen el ligando. En canales de voltaje, el canal se cierra cuando el potencial se restaura. En algunos canales de voltaje, la apertura implica su cierre. Algunos canales de voltaje poseen un tercer estado, el inactivo, que lo hace insensible al estímulo que lo abre. Gracias a este estado intermedio, el impulso nervioso sólo se transmite en una dirección. Permeasas: están en continuo movimiento. Sufren un cambio conformacional, por lo que el transporte es más lento. Este transporte se produce a favor del gradiente. Las permeasas fluctúan constantemente entre dos posiciones (la extracelular y la intracelular) Las permeasas pueden interaccionar con su estimulador. El transporte más característico es el del transporte al interior de la célula de la glucosa, que una vez en el interior se degrada o se polimeriza formando glucógeno. Tipos: Uniporte: se produce el transporte de una única molécula en la permeasa. Cotransporte: se produce el transporte de varias sustancias. Tipos: Simporte: en la misma dirección. (Glucosa+Na+) Antiporte: en direcciones contrarias. (ADP/ATP de la mitocondria)
TRANSPORTE ACTIVO
Bombas: estructuralmente son prácticamente iguales a las permeasas. Las bombas son capaces de utilizar diferentes formas de energía, utilizada para transportar sustancias en contra del gradiente. También tienen esa fluctuación constante entre dos posiciones, como las permeasas. Son capaces de funcionar, también, en dos direcciones: trabajar en contra del gradiente o dejar pasar sustancias a favor del gradiente, generando energía. Existen varias fuentes de energía: ATP: se puede hidrolizar, generando la suficiente energía. LUZ: algunas bombas emplean la luz como fuente de energía. Ej.: bacteriorrodoxina (bomba que emplea la luz para el transporte de H+^ en las bacterias) POTENCIAL ELECTROQUÍMICO (GRADIENTE): el potencial más fuerte es el del Na+. La célula puede emplear este potencial para arrastrar a otras moléculas en contra de su gradiente. GRADIENTE REDOX Podemos clasificar las bombas según su estructura: Tipo P: transporte de iones. Bomba Ca2+^ y la bomba Na+/K+. Tipo F: transporte de iones. Bomba de H+^ mitocondrial. Tipo ABC: transporte de sustancias de mayor tamaño (pequeñas moléculas) Azúcares, aminoácidos, nucleótidos y agentes tóxicos. Bomba de Na+/K+ La bomba introduce en la célula 2 moléculas de K+^ por cada 3 de Na+^ que saca, ambos en contra del gradiente. Este transporte viene propiciado por un gasto de energía en forma de ATP. Tiene como funciones principales: Principal generador del gradiente electroquímico de la célula. El balance total cuando la bomba actúa, es de una carga negativa en el interior de la célula, generando un gradiente. Es importante, ya que el gradiente generado también se emplea como fuente de energía para el trasporte de otras sustancias. Osmolaridad: determina la regulación del volumen de la célula. Se basa en el computo total de las concentraciones de
Macropinocitosis Vesículas desnudas Vesículas recubiertas de caveolina Vesículas recubiertas de clatrina: conocida también como endocitosis mediada por receptor. Una serie de componentes de la célula intervienen de manera secuencial en la incorporación de una sustancia de gran tamaño al interior celular. (LDL, partícula típica) La entrada de la LDL (lipoproteína que incorpora el colesterol a la célula) viene propiciada por una proteína específica transmembrana (ligando) que interaccionan con la partícula. Estas partículas chocan en su viaje por el medio extracelular con la membrana. Cuando se topan con su proteína receptora, quedan asociadas a ella. Estos receptores también interaccionan con otras proteínas del citosol. La primera proteína, que reconoce al receptor es la ADAPTINA (interacciona por fuerzas débiles) Esta adaptina interacciona con otra proteína interior, llamada CLATRINA. Cuando se produce la interacción del ligando con la molécula que entrará, se producen cambios confomacionales en las proteínas implicadas, que hacen que la membrana se curve. Las interacciones entre las proteínas implicadas en esta pinocitosis actúan en cadena provocando cambios que curvan la membrana. La clatrina forma un entramado, polimerizándose, que hace que se curve la membrana. El monómero de clatrina tiene forma de trisquelion (aspa tridimensional) La dinamina es la proteína responsable de estrangular la vesícula y separarla de la membrana.
Con la vesícula ya formada, el revestimiento de clatrina y la adaptina se desprenden, quedando la vesícula con los receptores y las moléculas de ligando en el interior. Después, esta vesícula sigue una ruta celular que la pondrá en contacto con el ENDOSOMA (compartimento membranoso que almacena las vesículas que llegan por endocitosis) El ENDOSOMA PRIMARIO posee en su membrana proteínas de membrana que bombean protones desde el citosol al interior reduciendo su pH (=6,5). Este cambio de pH hace que las proteínas receptoras cambien su forma, soltando las partículas que transportan. El ENDOSOMA INTERMEDIO recibe a los ligandos que son transportados desde el primario. Este compartimento tiene un pH menor (=5,5), propiciado por la entrada de protones. El ENDOSOMA SECUNDARIO también recibe a los ligandos en un ambiente de menor pH (=4,5). El paso por todos los compartimentos del endosoma transforma los componentes que se han introducido a la célula. Después las sustancias viajan al LISOSOMA a través de vesículas, donde se digieren (hidrolasas) (pH=4). Las proteínas receptoras de la membrana que se fusiona con los compartimentos del endosoma, son recicladas y devueltas a la membrana plasmática. *La célula indentifica estos compartimentos del endosoma, a través de los componentes de su membrana. Uno de los principales señalizadores es el fosfatidilinositol. Esta molécula se puede fosforalizar de maneras diferentes, sirviendo de identificación para cada uno de los compartimentos. También intervienen proteínas en la señalización, como las proteínas RAB. Fagocitosis: partículas de gran tamaño. El proceso es parecido al de la pinocitosis, pero con diferencias: No se produce un recubrimiento de clatrina, por lo que la vesícula de fagocitosis se forma proyectando la membrana plasmática hacia fuera captando la partícula y
TRANSCITOSIS Se lleva a cabo por vesículas formadas por endocitosis que no terminan en el lisosoma, sino que pasar a ser vesículas destinadas a la exocitosis. Estas vesículas sufren un tránsito a través de la célula. La transcitosis sirve para que determinadas células que absorben sustancias que no van destinadas a ellas mismas, pueden expulsarlas por el lado opuesto (cruzar vasos sanguíneos, células del intestino) Por ejemplo, las IgA de la leche materna, cuando llegan al intestino del lactante no pueden ser digeridas, sino asimiladas y llevadas al torrente sanguíneo, por lo que deben atravesar las células del intestino sin sufrir cambios.
TEMA 4. SUPERFICIE CELULAR
ADHESIÓN CELULAR
Las células son capaces de adherirse firmemente a otros componentes que las rodean, como otras células o el medio (matriz extracelular). Para llevar la adhesión a cabo, la célula posee proteínas (transmembrana) situadas en su membrana, existiendo cuatro familias principales: Cadherinas: se trata de proteínas de membrana que se asocian formando un dímero. Son capaces de asociarse con otras cadherinas presentes en células contiguas, por lo tanto, se trata de una interacción homófila. Estas proteínas aparecen en: tejidos de tipo epitelial, en el sistema nervioso y en tejidos musculares cardíacos. Selectinas: proteínas que forman monómeros. Intervienen en la adhesión célula-célula, asociándose a otro tipo de componentes (interacción heterófila) como los azúcares de las proteínas de células vecinas. Integrinas: forman dímeros, pero con unidades diferentes, por lo que se trata de heterodímeros. Intervienen en la adhesión dual (célula-célula/célula-matriz). Poseen una interacción de tipo heterófilo, ya que cuando proveen la adhesión con una célula vecina, interaccionan con una proteína de tipo Ig-CAM. Cuando Son dependientes de Ca 2+ , por lo que la adhesión sólo se produce si existen unos determinados niveles de este ión en el ME endi de
proveen la adhesión con la matriz, interaccionan con componentes de la misma, de tipo fibroso como la fibronectina y laminina. Ig-CAM: forman monómeros, mediando en la asociación celula-célula. Poseen una interacción homófila con la Ig-CAM de otra célula y heterófila con las integrinas. Forman parte de las inmunoglobulinas, apareciendo normalmente en células de la sangre y del sistema inmune. Habitualmente las interacciones que se producen entre las proteínas de adhesión o estas con los azúcares son muy versátiles y fácilmente regulables, ya que son débiles y se pueden deshacer a través de cambios en el medio. La desventaja de su debilidad se ve compensada con el “efecto velcro” (muchas interacciones juntas) TIPOS DE UNIONES CELULARES Transitorias: se establecen entre células cuyo estado fisiológico propicia la unión con una célula determinada. Un cambio en este estado propicia la desunión y la interacción con otra célula. Por ejemplo: neuronas (cadherina y las Ig-CAM), células de la sangre con los vasos sanguíneos (un glóbulo blanco se encuentra rodando por los vasos hasta que, si se produce una infección, queda adherido más fuertemente a esa zona, permitiendo que el glóbulo atraviese el vaso y se fije en el foco de la infección), formación de trombos por las plaquetas (mecanismo de adhesión entre plaquetas para formar un coágulo Permanentes: se establecen entre células vecinas o entre una célula y la matriz que la rodea. Esta unión tiene una duración casi vital y vienen asociadas a estructuras específicas de la célula o uniones celulares. Existen tres tipos de uniones celulares: OCLUYENTES: las células que revisten el intestino (o enterocitos) están caracterizadas por el desarrollo de todos los tipos de uniones celulares. La unión ocluyente se produce en la zona apical (en la punta) que da a la luz del intestino. Esta unión se produce gracias a que las membranas de células vecinas se aproximan, ocluyéndose, y desapareciendo el espacio entre ellas. Estas uniones se producen por proteínas transmembrana, asociadas a la célula vecina, que interaccionan muy estrechamente. Las proteínas que aquí intervienen son de la familia de las claudinas y las ocludinas. Para que se produzca un aislamiento total, se produce un sellado completo del borde de la célula, asociándose varias proteínas y formando un conjunto que parece un “pespunte” Son estructuras que se extienden por todo el perímetro celular, denominadas zonula ocludens. Aparecen varias hileras de proteínas entre las células vecinas. Las uniones en la parte apical de la célula contribuyen a impermeabilizar al organismo, sin permitir que cualquier sustancia penetre en él. Las proteínas o estructuras de adhesión están asociadas en el interior a elementos del citoesqueleto. En el caso de las ocluyentes, se encuentran asociadas a filamentos de actina. Las uniones entre las proteínas y el in en
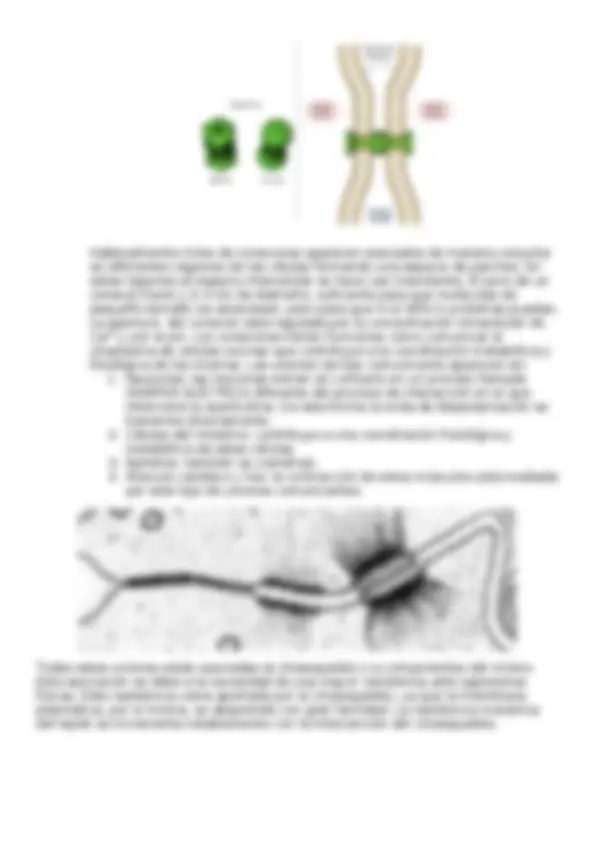
Contactos focales: la proteína transmembrana (INTEGRINA) interactúa con la matriz. En la parte interna de la célula, interaacciona con el citoesqueleto de filamentos de actina. Las proteínas puente en este caso son la talina y la vinculina. Hemidesmosomas: la proteína transmembrana es la INTEGRINA. Se trata de una estructura similar al desmosoma, pero es la mitad de la estructura, por tanto, sólo se forma un “disco”. La integrina interacciona con la matriz extracelular y está unida en el interior de la célula a la PLECTINA. La plectina la une al citoesqueleto de filamentos de queratina. UNIONES COMUNICANTES: median la comunicación entre células vecinas (unión célula-célula). Aparecen asociadas 6 CONEXINAS en la membrana de la célula formando una estructura con un canal en el centro. Esta estructura se denomina CONEXÓN, que establece un puente de conexión entre células vecinas.
Habitualmente miles de conexones aparecen asociados de manera conjunta en diferentes regiones de las células formando una especie de parches. En estas regiones el espacio intercelular se hace casi inexistente. El poro de un conexón tiene 1,5-3 nm de diámetro, suficiente para que moléculas de pequeño tamaño los atraviesen, pero para que ni el ARN ni proteínas puedan. La apertura del conexón está regulada por la concentración intracelular de Ca2+^ y por el pH. Los conexones tienen funciones como comunicar el citoplasma de células vecinas que contribuye a la coordinación metabólica y fisiológica de las mismas. Las uniones de tipo comunicante aparecen en:
- Neuronas: las neuronas entran en contacto en un proceso llamado SINÁPSIS ELÉCTRICA diferente del proceso de interacción en el que interviene la acetilcolina. De esta forma la onda de despolarización se transmite directamente.
- Células del intestino: contribuye a una coordinación fisiológica y metabólica de estas células.
- Epitelios: también se coordinan.
- Músculo cardíaco y liso: la contracción de estos músculos está mediada por este tipo de uniones comunicantes. Todas estas uniones están asociadas al citoesqueleto o a componentes del mismo. Esta asociación se debe a la necesidad de una mayor resistencia ante agresiones físicas. Esta resistencia viene aportada por el citoesqueleto, ya que la membrana plasmática, por sí misma, se desprende con gran facilidad. La resistencia mecánica del tejido se incrementa notablemente con la intervención del citoesqueleto.