¡Descarga Capítulo de Geneser histología y más Guías, Proyectos, Investigaciones en PDF de Histología solo en Docsity!
CAPÍTULO 14
Tejido nervioso
"Hermoso es lo que vemos. Más hermoso es lo que sabemo s. Pero mucho más hermoso es lo que no conocemo s". Ni el s Stee ns en
Las funciones normales del organismo dependen hacia uno o más órganos efectores (músculos de la recepción de estímulos desde el ambiente esqueléticos, músculos lisos o glándulas) o sólo externo (exterorrecepción) y propios (intero se almacene como información. El cerebro, en rrecepción), y de la generación de reacciones especial la corteza cerebral, utiliza la informa integradas dirigidas en respuesta a esos estímulos. ción almacenada en funciones "superiores" tales Esta actividad requiere un eslabón coordinador como el pensamiento abstracto y la memoria, que entre el estímulo y la respuesta, entre el órgano constituy en la base estructural y química para la receptor y el órgano efector. existen ci a consciente. En el organismo de los mamíferos se desaJTolla Algun as célu las nerviosas realizan sus fun ron dos sistemas coordinadores o in tegradores que ciones integradoras por medio de hormonas relacionan el estímulo con la respuesta: el sistema sinte ti zadas en el cuerpo celular y liberadas por endocrino y el sistema nervioso. Mientr as q ue el las te rminaciones nerviosas para ser transpor sistema endocrino por lo general media reacciones tadas por la sangre haci a los órganos diana. Se más difusas y prolongadas, el sist ema ne rv ioso dice q ue estas cél ul as nerv iosas tienen actividad representa la base estructur al para las re acciones neurosec re tora. precisas, rápidas y, casi siempre, más cortas. Tradicionalmente, el si stema nervioso se divi El sistema nervioso incluye todo el teji do de en sistema nervioso central y sistema nervioso nervioso del organismo y su func ió n principal periférico. es la comunicación ; las cél ul as nerviosas están El sistema nervioso central (SNC) está especializadas para ello, con sus prolongaciones compuesto por el encéfalo, encerrado en el crá muy largas y propiedades electr ofisiológicas neo, y su continuación hacia abajo, la médula especiales. En la célula nerviosa o neurona, las espinaL ubicada en el conducto raquídeo. En los funciones celulares especializadas de irritabilidad mamíferos, la mayoría de las células nerviosas se y conductividad alcanzan su máximo desarrollo. encuentran en el sistema nervioso central. Entre La irritabilidad indica la capacidad de un a c él ula las células nerviosas existen contactos celulares para reaccionar ante distintos estímul os. La con (^) de un tipo especial , la sinapsis (gr. synapsis, ductividad es la capacidad de enviar (transmitir) unión), a través de la cual una célula nerviosa hacia otras partes de la célula un a señal de que puede influir sobre la acti vi dad eléctrica de otra se ha producido a un estímul o. A esto se agrega cél ul a nerviosa medi ante sustancias químicas la capacidad de transferir con rapidez una señal transmisoras. El sistema nervioso central hu a otras células (transmisión de señales). Las cé mano con ti ene miles de mi llones de neuronas lulas nerviosas se irritan o se estimulan con gran relacionadas funcionalmente a través de sinapsis, facilidad, lo que genera una onda excitadora o a menudo con varios miles en cada neurona. Por impulso nervioso que puede transmitirse a través lo tanto, la complejidad de esta red de sinapsis de distancias considerables como una diferencia es muy grande. progresiva de potencial eléctrico a lo largo de la En el sistema nervioso central, la mayor parte fibra nerviosa. de los cuerpos celulares de las neuronas están Ante la acción de un estímulo determinado reunidos en grupos de distintos tamaños, denomi (p. ej., presión sobre la piel, fotones en el ojo, nados núcleos. Por lo general, las largas prolon ondas sonoras en el oído), estas distintas formas (^) gaciones de las células nerviosas están rodeadas de energía son transformadas (transducidas) en por una vaina más o menos desarrollada (véase actividad eléctrica por estructuras celulares espe más adelante, pág. 336) y se denominan fibras ciales, los receptores sensoriales. La actividad (^) nerviosas. A menudo, transcurren reunidas en un eléctrica se transmite como impulsos nerviosos haz desde una parte del sistema nervioso central a al sistema nervioso central, donde son modifica otra para formar untracto o vía de conducción. dos y elaborados por otras células nerviosas. El El sistema nervioso periférico (SNP) com resultado de esta elaboración puede ser que se prende todo el tejido nerviosofitera del sistema envíen mensajes en forma de impulsos nerviosos nervioso central. Está compuesto por grupos de
Tejido nervioso I 15
BOOK´S COPICENTRO USAC / 2016
cuerpos de células nerviosas, o ganglios, redes de fibras nerviosas, o plexos, y haces de fibras nerviosas de recorrido paralelo en la forma de nervios periféricos, Los nervios parten del encéfalo y de la médula espinal de a pares, uno para cada lado del organismo. Los nervios del encéfalo se denominan nervios craneales y los nervios que provienen de la médula espinal, nervios espinales. Los nervios periféricos comunican las neuronas del sistema nervioso central con todas las partes del organismo. Las fibras nerviosas que llevan los impulsos desde el sistema nervioso central a la periferia se denominan eferentes (la!. effero, sacar de) o motoras (lal. moveo, mover), mientras que las fibras nerviosas que conducen impulsos desde la periferia hacia el sistema nervioso central se denominan aferentes (la!. affero, llevar a) o sen sitivas (lat. sentio, sentir). Las fibras nerviosas son motoras o sensitivas, pero por lo general se encuentran entremezcladas en los nervios craneales y espinales. Los nervios se ramifican en su camino hacia la periferia, donde presentan terminaciones libres en los tejidos periféricos u órganos terminales especializados. Además de las neuronas, el sistema nervioso incluye también células especiales de sostén, la neuroglia (gr. gUa , pegamento) y tejido conec tivo. Este último conforma las meninges encefá' licas y espinales que rodean el sistema nervioso central, y también aparecen en escasa cantidad a lo largo de los vasos sanguíneos que ingresan en el SNC. El tejido conectivo interviene también en la formación de los nervios periféricos, los ganglios y los órganos sensoriales. A continuación se verán la histología y la citología de los elementos celulares del sistema nervioso central, es decir, la neurohistología.
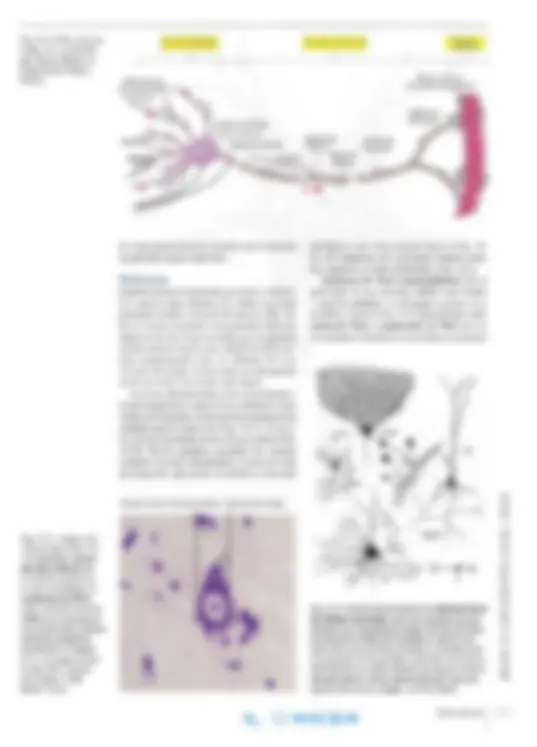
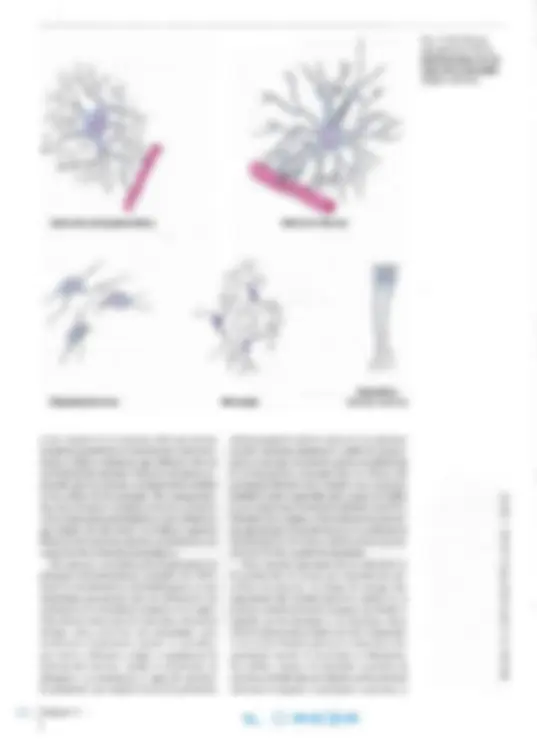
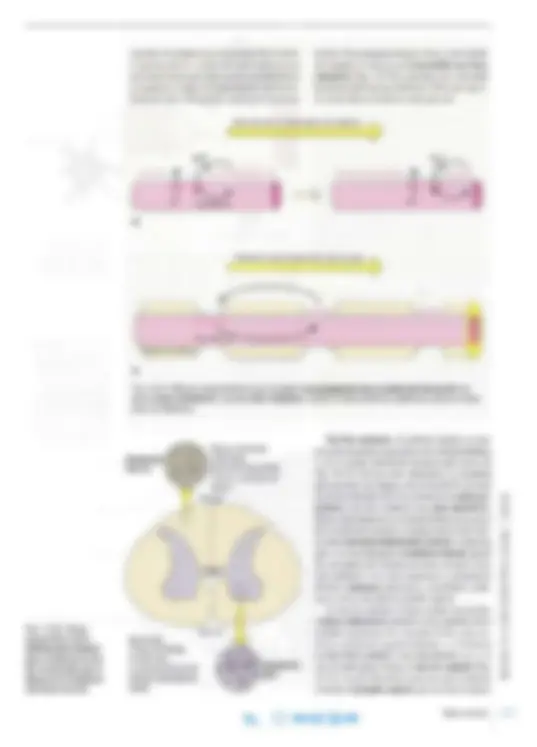
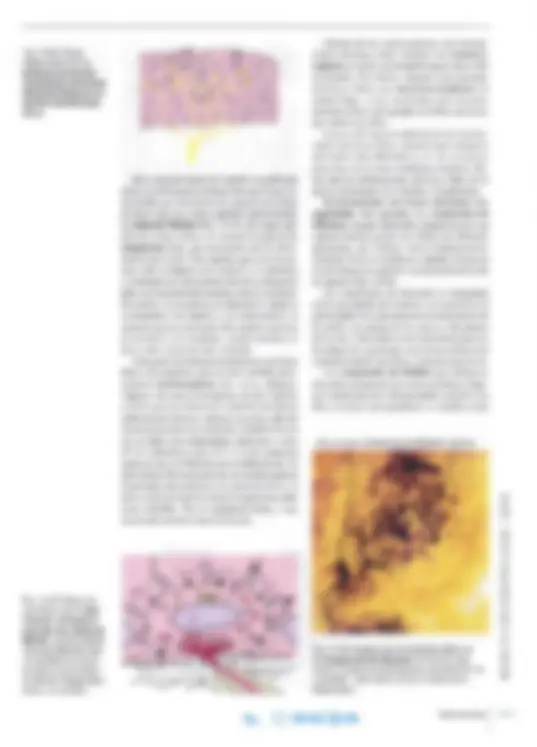
Soma Dendritas Espinas Axón
a^ f
31 I Capítulo 14
Neuronas
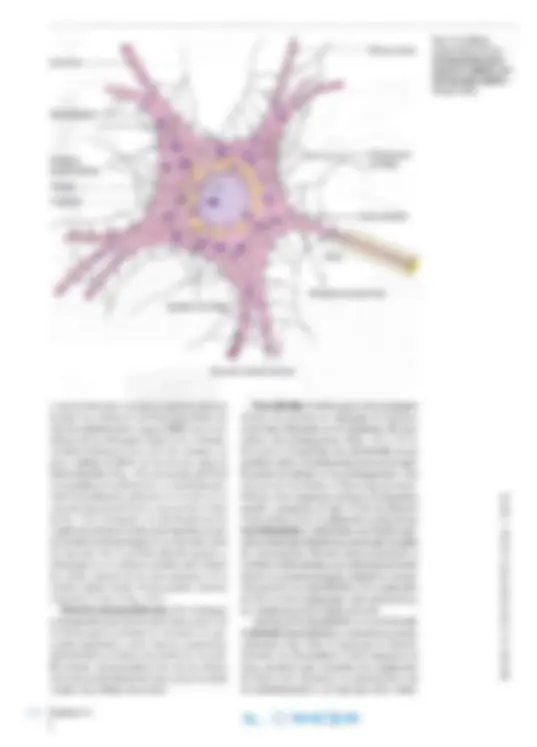
La neurona es el cuerpo de la célula nerviosa con todas sus prolongaciones. La fomla de la neuro na se visualiza mejor en cortes gruesos teñidos mediante técnicas de impregnación con metales pesados (Fig. 14-1a) o tras microinyección con una molécula marcada (Fig. 14-1 b; véase también el Capítulo 2, pág. 43). Todas las neuronas poseen un cuerpo celular o soma compuesto por el núcleo rodeado por cantidades variables de citoplasma (Fig. 14-2). El citoplasma que rodea el núcleo se denomina pericarion, de donde salen largas pro longaciones citoplasmáticas, de las cuales todas las neuronas poseen por lo menos una. A menudo exis ten numerosas prolongaciones cortas ramificadas, las dendritas (gr. dendrites , referente a árboles; es decir, ramas de árbol) y una prolongación larga, el axón (gr. axon, eje), que en algunos casos puede medir más de un metro de largo. Con frecuencia, el axón emite ramificaciones, las colaterales, a lo largo de su recorrido y además presenta ramifica ciones preterrninales justo antes de donde temuna la neurona y forma contactos sinápticos. Neuritas es la denominación común de las den dritas y los axones, y se utiliza cuando no es posible establecer si una fibra nerviosa es dendrita o axón.
Núcleo
El núcleo es de oval a redondo y grande en re lación con el tamaño celular; por lo general, se ubica en el centro del cuerpo celular. En células nerviosas grandes, los núcleos son claros, con distribución uniforme de cromatina de grano fino, y suelen poseer un nucléolo grande visible contra el fondo débilmente teñido (Fig. 14-3). En neuronas pequeñas, el núcleo es más oscuro, con granos de cromatina más gruesos. Desde el punto
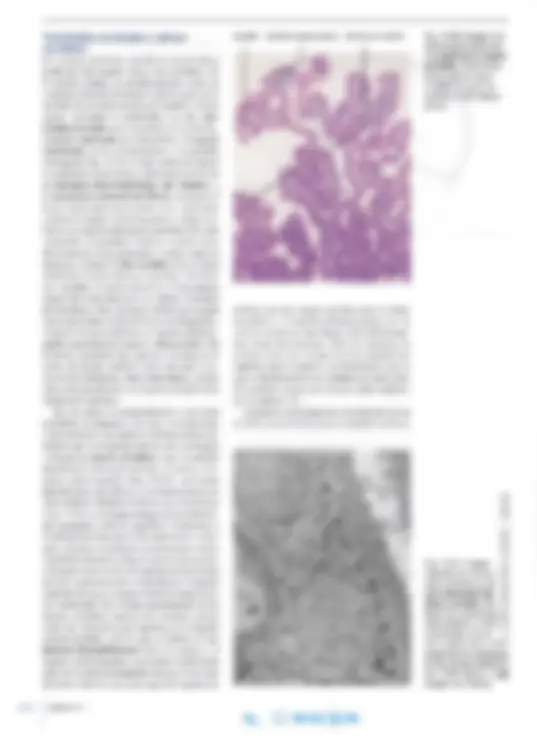
Fig.14-1.a Imagen con microscopio óptico de una neurona de la corteza cerebral. Pre parado por impregna ción según técnica de Golgi. x215. Barra: 50 ~m. b Imagen de una neurona en la que se efectuó microinyección (en la célula viva) de la enzima peroxidasa del rábano para luego determinar la enzima en el preparado, por método histoquímico. (Cedido por J. Zimmer).
BOOK´S COPICENTRO USAC / 2016
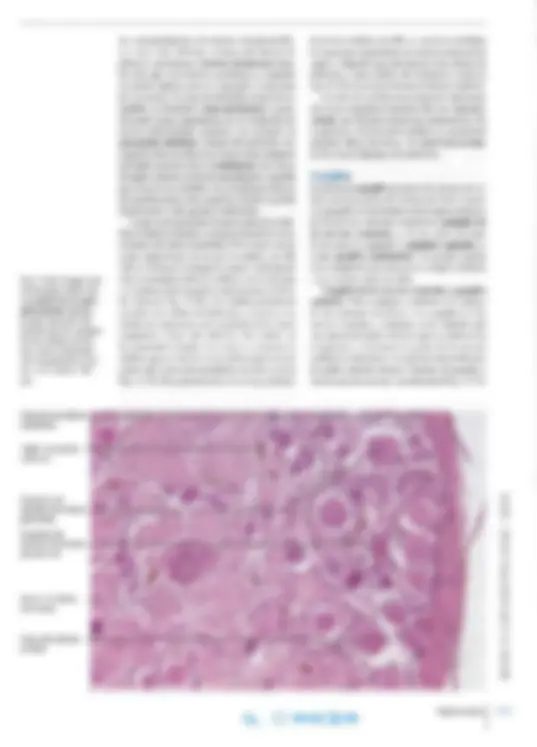
...•: (^) ¡,I--- --7"-- --- Mitocondrias Dendrita-------->,
Neurofibril las-"--...."---.l
Sinapsis ~~~~^ ~~^ .llfttlfj!;l--7-~?-===:-----..,orpúsculo de Nissl axosomáticas
Núcleo Nucléolo -7~--- Cono axónico
Sinapsis axodendríticas
vivas no coloreadas con microscopia de contraste de fase. Los corpúsculos de Nissl son cúmulos de retículo endoplasmático rugoso (RER) en el que, además de los ribosomas fij ados a las cisternas, se hallan ribosomas libres entre las cisternas. La gran cantidad de RNA así acum ul ada causa la intensa basofilia (Fi g. 14-6). La sustancia de Nissl se encuentra en el pericarion y en la pri mera por ción de las dendrita s, pero falta en el axón y en el cono de iniciación del axón o cono axónico (véase la Fig. 14-5). El tamaño y la distribución de los corpúsculos de Nissl ti enen gran impo rt ancia para la identificación histológica de los distintos tipos de neuronas. Por lo general, son más gruesos y abundantes en las neuronas gr an des, por ejem plo las células motoras de las ast as anteriores de la médula espinal , donde forman grandes cúmulos irregulares (véase la Fig. 14-3). Retículo endoplasmático liso. No se di stingue en preparados para microscopia óptica, pero con la microscopi a electrónica se enc uentra en can ti dades importantes, sobre todo en el pericario n, desde donde se extiende a las dendrit as y el axón. El retículo endoplasmático liso de las células nerviosas puede almacenar iones calcio de modo similar a las células musculares.
31 I Capítulo 14
Neurofibrillas. Se distinguen en los preparados teñidos con técnicas de impregnación argéntica como finos filamentos en el citoplasma del peri carion y las prolongaciones (Figs. 14-5 y 14-7). En el axón y las dendritas, las neurofibrillas corren paralelas entre sí. Se demuestran hasta en las rami ficaciones terminales de las prolongaciones. Con microscopia electrónica, se observa que las neuro fi brillas están compuestas por haces de filamentos grandes y pequeños de unos 10 nm de diámetro (véase la Fig. 14 -6). Los filamentos se denominan neurofilamentos y representan una familia espe cial de filamentos intermedios, por lo que son parte del citoesqueleto. Forman entrecruzamientos y confieren sostén mecánico, en particular en el axón, donde se encuentran en gran cantidad en orienta c ión paralela. Las neurofibrillas y los corpúsculos de Nissl son los componentes más característicos del citoplasma de las células nerviosas. Además de las neurofibrillas se encuentran mi crotúbulos (neurotúbulos), a menudo en grandes cantidades (Fig. 14-8). Al igual que los neurofi lamentos, los microtúbulos suelen disponerse en haces paralelos que circundan los corpúsculos de Niss l. Con frecuencia se entremezclan con los neurofilamentos y, al igual que éstos, contri-
Fig. 14-5. Dibujo esquemático de los componentes de la neurona visibles con microscopio óptico. (Según Barr).
BOOK´S COPICENTRO USAC / 2016
Fig. 14-6. Imagen de Aparato^ de^ Golgi^ Mitocondrias^ Neurofilamentos una parte de la neuro na, captada con mi croscopio electrónico, en la que se observan zonas localizadas de retículo endoplasmático rugoso. Además se observan el aparato de Golgi, numerosas mito condrias pequeñas 'fila mentosas y pequeños haces de neurofilamen tos entre las zonas de retículo endoplasmático rugoso. x18.000. Barra: 500 nm. (Cedido por E. Westergaard).
Fig. 14-7. Imagen con microscopio óptico de una neurona de la médula oblongada (bulbo raquídeo) que muestra las neurofibri lIas en el citoplasma. Tinción de Cajal. x760. Barra: 10 IJm.
buyen a dar rigidez y estabilizar la morfología de la célula. Esto es especialmente válido para el axón, donde haces paralelos de microtúbulos contribuyen a mantener la forma extendida. Los microtúbulos del axón adquieren su rigidez por estar unidos entre sí o con la membrana celular u otras partes del citoesqueleto mediante dos proteínas asociadas con los microtúbulos (MAP). Todos los microtúbulos de las dendritas y el axón de la neurona tienen la misma orientación, con el extremo minus dirigido al cuerpo celular. Esto
Neurofibrillas
crea la base para otra función muy importante: el transporte axónico (véase más adelante). Por último , en la neurona se encuentran filamentos de actina, correspondientes a los filamentos de actina en otras células no muscu lares. Forman parte del citoesqueleto y tienen un diámetro de unos 7 nm. Aparato de Golgi. Sólo se encuentra en el pericarion, donde se visualiza como una red fila mentosa en preparados teñidos por los métodos de impregnación con cromo u osmio. Esta red se extiende como un anillo alrededor del núcleo (véase la Fig. 14-5); con microscopia electrónica, en el sitio correspondiente, se distinguen peque ños rimeros de cisternas de Golgi rodeados por vesículas de escaso tamaño (véase la Fig. 14-6). Mitocondrias. Se encuentran en cantidades importantes en el pericarion y en todas las prolon gaciones (Fig. 14-5). Las mitocondrias de las célula~ nerviosas son pequeña~, de menos de 111m , y suelen ser filamentosas y extendidas (Fig. 14-6). Con mi croscopia electrónica se distingue la configuración habitual, salvo que las crestas a veces son alargadas. Centrosoma. En los precursores embrio narios de las neuronas, que todavía proliferan, se observa un centrosoma con un par de cen trÍolos. Después de un examen minucioso , en las neuronas maduras se distingue un único centríol0 de importancia desconocida, dado que las neuronas maduras no se dividen.
Tejido nervioso I^319
BOOK´S COPICENTRO USAC / 2016
Fig. 14-9. Imagen cap tada con microscopio electrónico de un corte transversal de un axón del sistema ner vioso central. x65.000. Barra: 100 nm. (Cedi do por L. Heimer, en Flickinger.)
a las influencias de distintos neurotransmisores. En casos aislados, las dendritas forman sinapsis dendrodendríticas entre sí (véase más adelante). Axón. Nunca sale más de un axón de cada neurona. Por lo general, el axón surge de una pequeña saliente del cuerpo celular o de la primera porción de una dendrita. Este cono de iniciación o cono axónico se caracteriza por su falta de sustancia de Nissl (véase la Fig. 14-2). El axón suele ser mucho más largo y más delgado que las dendritas de la misma neurona, con un diámetro más uniforme. A lo largo de su recorrido, puede emitir ramas colaterales que abandonan el tron co principal en forma casi perpendicular (véase la Fig. 14-2), mientras que las dendritas suelen ramificarse en ángulos agudos. Las colaterales se encuentran sobre todo en el sistema nervioso central; en ocasiones, son muy numerosas. De este modo, la neurona puede establecer con tacto con muchas otras neuronas. Cerca de la región terminal , a menudo el axón se divide en un ramillete de ramificaciones preterminales que suelen terminar en un bulbo de gran tamaño denominado bulbo terminal o botón sináptico (véase la Fig. 14-5). El citoplasma del axón o axoplasma es conti nuación del citoplasma del pericarion y contiene mitocondrias, túbulos alargados de retículo en doplasmático liso, microtúbulos y gran cantidad de neurofilamentos, pero ningún corpúsculo de Nissl (Fig. 14-9). El plasmalema que rodea el axón se denomina axolema. Muchos axones están
rodeados por una vaina de mielina (véase más adelante) que no es parte de la neurona. La reacción de las neuronas ante los estímulos que recibe el árbol dendrítico es transmitida a través del axón como un potencial de acción que se difunde por medios electroquímicos según el principio del todo o nada. La primera porción del axón, desde el cono de iniciación hasta el comienzo de una eventual vaina de mielina, se de nomina segmento inicial (véase la Fig. 14-2). El pericarion y las dendritas poseen una membrana que se excita ante estímulos eléctricos, pero por lo general el potencial de acción se desencadena en el segmento inicial, que presenta un umbral menor para la excitabilidad eléctrica y que, además de los canales de sodio rápidos activados por voltaje, cuenta con una cantidad especial de canales de potasio y de calcio. Esto permite que la frecuen cia de descarga de los potenciales de acción se corresponda con el grado de despolarización del axolema en el segmento inicial, es decir que el grado de despolarización de las dendritas y el soma de la célula nerviosa se transforman en una frecuencia de descarga (una forma de conversión de analógico a digital). Además de la difusión de los potenciales de acción a lo largo de la membrana axónica tiene lugar un transporte axónico, es decir, el desplazamiento de sustancias dentro del axón. Como se mencionó antes, el axón no contiene retículo endoplasmático rugoso, por lo que no hay síntesis local de proteínas de membrana. En
Tejido nervioso I 321
BOOK´S COPICENTRO USAC / 2016
consecuencia, las proteínas y las estructuras de membrana, por ejemplo las enzimas, los recep tores del axolema y los o rg ánulos li mitados por membrana, só lo se sintetizan en el cuerpo de la célula nerviosa y la parte proximal de las dendri tas. Sin embargo, pequeñas cantid ad es de algun as proteínas del citos ol son sintetizadas en el propio axón (con la ayuda de ribosomas libres), entre ellas, ciertas proteín as de los neurofilamentos. En consecuencia, las proteínas y los orgánulos d el axón deben ser transportados ca si en su totalidad desde el cuerpo celu lar hacia el axón, fen ómeno que se observa direc tamente en los cultivos de tejido como fl ujo axó ni co de vesíc ulas. La inyección local de aminoácidos marcados cerca del pericarion de la célula ne rv iosa, seguida por radioautografía, demuestra que en el pericarion tiene lugar la síntesis de las proteínas que luego son transportadas hasta las termi naciones axó nicas. También la constricción axónica con fi rma la existencia de esta forma de transporte, dado que hay acum ul ación de material a ambos lados de la constricción , c om o signo del transporte en ambas direcciones. El transporte axónico lento ocurre en direc ción anterógrada (hacia la periferia de la célul a) con una velocidad de 0,2-8 mm por día y abarca la mayor parte de la s p ro teínas sujetas a transporte axónico. De este modo, se entregan al axón los componentes necesarios para su manterumien to , por ejemp lo, moléculas de tubulina, cie rt as subunidades de neurofilamentos y otras pro te ínas solubles, como las enzima s. Con el transporte axónico r ápido hay trans porte anterógrado (centr ífugo) de or gánulos limitados por membrana, por ejemplo, mitocon drias, fragmentos de ret íc ulo endoplasmático liso y vesículas, con una velocidad de 100-400 mm por día. El transporte axónico rápido incluye también a las enzimas que catalizan la síntes is de las sustanc ias transm isoras en los bulbos terminales. El transporte axónico rápido también es en dirección retrógrada (centrípeto). Este transporte desde las term in aciones axónicas en dirección al cuerpo celular incluye los compo nentes celulares desgastados con el fi n de su degradación y posible reutilización de s us consti tuyentes. Además, en las terminaciones tam bi én se captan sustancias que son transportadas hacia el cuerpo celular. Esto permite la información retrógrada (p. ej., a los genes del núcleo celular) sobre el estado de las terminaciones axónicas y las condiciones en su entorno para que la célula pueda efectuar ajus te s ( p. ej ., modificaciones de la síntesis de proteínas) como adaptación an te los procesos que ocurren en estas ter minaciones axónicas (véase más adelante, en terminaciones axónicas y sinapsis).
32~ I^ Capítulo 14
El mecanismo para el transporte axónico rápido está condic io nado por la presencia de m i crotúbulos, ya que la colchicina puede inhibir el transporte; la colchicin a se fij a a los dímeros de tubulina e impide su polimerización a microtú bulos. Los microtúbulos del axón siempre tienen la misma p ol aridad, con el extremo plus dirigido haci a la terminación del axón. P or eje mplo, el transporte de una vesícula tiene lugar cuando moléculas de cinesina o dine ín a cito pl asmática se fi ja n al orgánulo con la "cola" y luego migran hacia la super fi cie del microtúbulo por un. ión de las cabezas (v éanse también microtúbulos, Capítulo 3, pág. 105, Y la Fig. 3 -5 0). Dado que las moléculas de cinesina siempre migran h ac ia el extremo p lu s d el microtú bu lo mientras que las moléculas de dineí na citoplas máticas lo hacen hacia el extremo minus, el transporte anterógrado está condicionado por la cinesina, mientras que el transporte retrógrado hacia el cuerpo celular depende de la acción de la dineína citoplasmática. El mecanismo del transporte axónico lento no está aclarado en forma de l1n itiva. Las más recientes investigac iones sugieren. que bá si camen te es igual al transporte axónico rápido pero con cortos períodos de transporte rápido interrumpido po r pausas de longitud variable, por lo que la velocidad en promedio es unas 100 veces menor que la del transporte axónico rápido.
Tipos de neuronas y su presentación
Las neuronas pueden clasificarse de acuerdo con la c an ti dad de prolongaciones y la lon gitud de los axones. Según la cantidad de prolongaciones, las neuronas se clas ifi c an en unipolares , bipolares y multipolar es (Fig. 14- 10). Las neuronas unipola res ti enen sólo una prolong ac ión. Son escasos los ejemplos verdaderos de es te tipo (véase neuronas seudounipolare s) , pero se encuentran en el núcleo mesencefálico del nervio trigémino. Las neuro nas bipolares emiten una prolongación desde cada extremo del cuerpo celular, que tiene forma ahusada. Las neuronas bipolares se encuentran en la retina, el ganglio vestibular y el ganglio espiral del oído in te rno (r el acionados con el nervio ves
.--~---~
~ -- ------'- -------
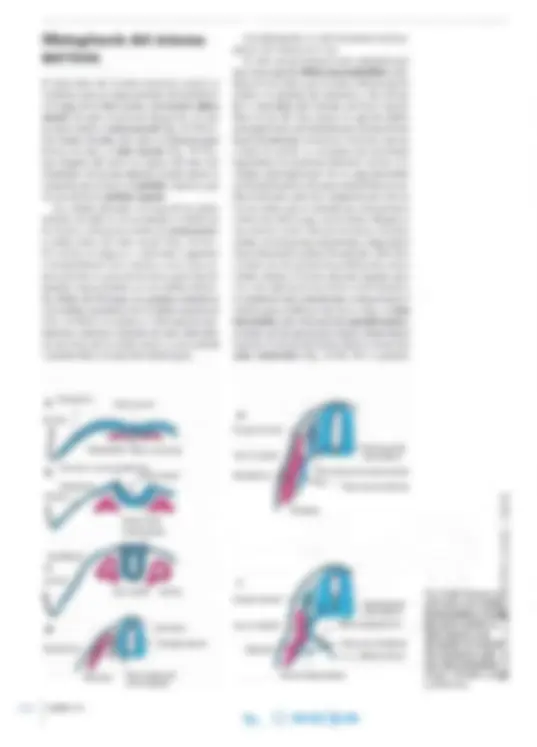
Fig. 14-10. Dibujo esquemático de los 4 tipos en los que pueden clasificarse las neuronas sobre la base de la cantidad de prolongaciones.
BOOK´S COPICENTRO USAC / 2016
Terminaciones axónicas y sinapsis
Las neuronas del sistema nervioso y sus prolonga ciones conforman una red muy interrelacionada. Los contactos para la transferencia o transmisión de los impulsos nerviosos están estructurados de manera tal que la transmisión del impulso sólo puede producirse en una dirección. En lugar de actuar directamente sobre la célula vecina, el impulso nervioso que se desplaza por el axón y llega hasta la terminación nerviosa produce la liberación de una sustancia transmisora. Un neurotransmisor puede definirse como una sustancia química liberada por exocitosis de una terminación nerviosa como reacción ante el potencial de acción del axón, y que transmite la señal a otra célula (postsináptica) (neurona u órgano efector) que, debido a ello, es excitada o inhibida. La sustancia transmisora se difunde a través del espacio intercelular hasta la célula adyacente (postsináptica), donde se fija a molé culas receptoras específicas sobre la membrana superficial de esta célula. Entonces, la reacción entre el transmisor y las moléculas receptoras causa un cambio del potencial de membrana en la célula siguiente. La sinapsis es la zona especializada de con tacto donde tiene lugar la transmisión del impulso eléctrico, mediada por un neurotransmisor. La
324 ICapítulo 14
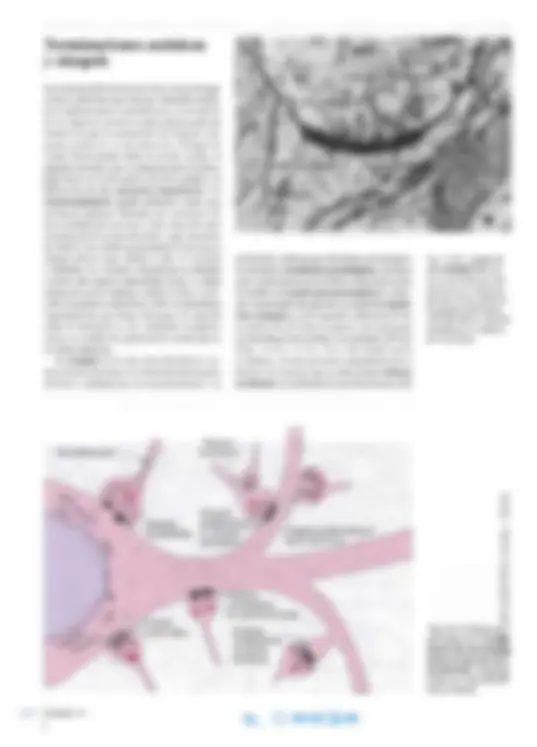
porción del axolema que interviene en la sinapsis se denomina membrana presináptica, mientras que el plasmalema de la célula contactada recibe el nombre de membrana postsináptica. La bre cha extracelular interpuesta se denomina hendi dura sináptica y, por lo general, mide unos 25 nm de ancho. Por lo tanto, es apenas más ancha que las hendiduras intercelulares circundantes (20 nm) (Figs. 14-12 y 14-13). En el sitio donde ocurre la sinapsis , el axón presenta ensanchamientos o botones (fr. bouton), que se denominan botones terminales si conforman la porción terminal del
Fig. 14-12. Imagen de una sinapsis del sis tema nervioso central, captada con microsco pio electrónico. Nótese el aspecto asimétrico. x75.000. Barra: 100 nm. (Cedido por L. Heimer, en Flickinger).
Fig. 14-13. Dibujo es quemático de la clasifi cación de las sinapsis sobre la base de su localización. (Según Bailey, en Copenhaver, Kelly y Wood).
Neurofilamentos
en espina dendrítica
BOOK´S COPICENTRO USAC / 2016
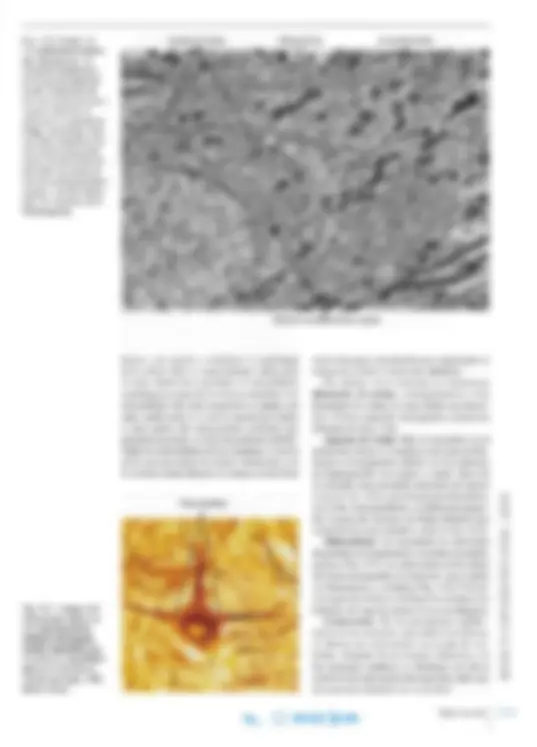
Fig. 14-14. Imagen con microscopio óptico de un axón del sistema nervioso central con numerosos botones de pasaje. Las fibras se tiñeron por método in munohistoquímico con anticuerpo monoclonal contra el neurotransmi sor serotonina. x540. Barra: 10 ~m.
Botón de pasaje
f>
'.
axón (Fig. 14-5 ) o botones de paso (fr. boutons en passant) si se encuentran a lo largo del axón (Fig.14-14). Mediante microscopia electrónica , se dis tinguen las terminaciones axónicas en contacto sináptico con los somas y las dendritas de las neuronas vecinas (véase la Fig. 14-11). Las terminaciones contienen escasas mitocondrias y numerosas vesículas sinápticas pequeñas , por lo general con un diámetro de unos 50 nm (véase la Fig. 14-1 2). Algunas de las vesículas están muy cerca de la membrana presináptica, que allí presenta un a condensación de materiaL citopLasmático sobre La supelficie interna. Esta zona se denomina zona activa. dado que es aquí donde se fijan las vesículas sinápticas y liberan los neurotransmisores a la hendidura sináptica. Frente a ésta también hay una condensación sobre la superficie interna de La membrana postsináptica. En algunas sinapsis también pue de aparecer material extracelular especializado entre las membranas de la hendidura sináptica. Este material comprende los dominios extrace lulares de las proteínas transmembrana de las membranas celulares presináptica y postsináp tica y por moléculas de la matriz extracelular. Estos componentes de la hendidura sináptica (p. ej. , moléculas de adhesión de células nerviosas [NCAM] e integrinas) representan la base de la fuerte unión entre las dos membranas de la sinapsis. Mediante la técnica de congelación y fractura, se ha demostrado que la condensa ción citoplasmática presináptica se organiza en forma de un patrón hexagonal con columnas de proteínas que se extienden hacia el interior del citoplasma. Se cree que las vesículas sinápticas son guiadas entre las columnas hacia sitios de anclaje y de liberación específicos (véase más adelante).
La presencia de vesículas sinápticas exclusi vamente en la región presinática implica que la sinapsis sólo transmite en una dirección, desde eL axón hacia la célula contactada. Asimismo, se ha demostrado que los transmisores se liberan en cantidades determinadas de algunos miles de moléculas y que las vesícul as sinápticas contie nen los neurotransmisores que se concentran, se envasan y se almacenan protegidos contra la degradación por enzimas citoplasmáticas. Las vesículas liberan el neurotransmisor a la hendidura sináptica por fusión con la membra na presináptica (exocitosis). La liberación del neurotransmisor es muy rápida comparada con la exocitosis en otros tipos celulares, dado que una porción mínima de las vesículas sinápticas, denominada fondo común libe rabIe , se loca liza junto a la membrana presináptica, a la que se fija para vaciarse de inmediato (Fig. 14-15). Se cree que las vesículas se unen justo frente a los canales iónicos de calcio activados por voltaje, los cuales se abren cuando un potencial de acción llega a la terminación axónica. Dado que la concentración de iones calcio es mucho mayor en el espacio extracelular que dentro de la terminación axónica, hay una difusión hacia adentro de iones calcio, lo que induce la fusión de la membrana de la vesícula sináptica con el plasmalema y la liberación del contenido de transmisor. Un mayor fondo común de reserva de vesículas sinápticas se localiza en el interior de la terminación , donde las ves.ículas se unen a filamentos de actina. Este depósito de reserva suministra vesículas sinápticas al fondo común liberable después de la exocitosis. Cuando las vesículas están llenas del neurotransmisor, son transportadas a la zona activa y allí se fijan al plasmalema. El proceso de transporte y fijación se denomin a acoplamiento de las vesículas. Varias moléculas proteicas intervienen en la unión exacta de las vesículas frente al sitio acti vo, la exocitosis y la posterior neoformación de vesículas, el denominado ciclo de las vesículas sinápticas. Se forman vesículas con cubierta de c1atrina por invaginación del plasmalema de la terminación axónica cerca de la sinapsis para reutili zar membrana en la producción de vesícu las sinápticas (véanse las Figs. 14 -12 y 14-15). Las vesículas se fusionan y forman un endosoma temprano, luego de perder la cubierta de clatrina. Luego se liberan nuevas vesículas sinápticas del endosoma y se unen a los filamentos de actina, tras lo cual captan neurotransmisores. Este ciclo local de vesículas sinápticas dura alrededor de I minuto en total, tras lo cual la exocitosis dura menos de 1 ms. Además del ciclo de las vesículas sinápticas descrito, hay fuertes indicios de que en algunos
Tejido nervioso I^325
BOOK´S COPICENTRO USAC / 2016
Receptores metabotrópicos y neuromodulación
Un ejemplo típico de un receptor metabotró pico es el receptor para acetilcolina (de tipo muscarínico) en el marcapasos cardíaco (nó du lo sinusal). Aquí, la unión de la acetilcolina con el receptor activa una proteína G y abre los canales iónicos de potasio. Dado q ue la concentración de iones potasio es mayor dentro de la célula que en el espacio extracelular, esto produce un fiujo de iones potasio hacia el exterior y un aumento del potencial de membran a. En con secuencia, di sminuye la frecuencia cardíaca. Como se mencionó en el Capí tu lo 7, el efec to es mediado por la proteína G por acc ió n sob re la aden il ato cic lasa y la consecue nt e modificación de la cantidad de AMP cíclico, que actúa como segundo mensajero dentro de la cél ul a. Los neu rotransmisores de tipo amínico actúan por unión con receptores metabotrópicos de esta clase. En ocasiones, los receptores metabotrópicos pueden actuar sobre la actividad de transcripción de la cél ul a a través del sistema intracelular de transducción de señales. De este modo, la célula sinte ti za proteínas que se incl uyen en la mem brana celular y así modifican sus carac te rísticas. Las modificaciones muy prolongadas, en algunos casos tal vez permanentes, de la sensibilidad postsináptica de la neurona f re nte a distintas acc iones representan el fundamen to de fenómenos como el aprendizaje y la me moria. Como se mencionó antes, muchas de las sustanc ias transmiso ras son péptidos, o neuropéptidos , que actúan to do s a través de receptores metabotrópicos. Lo mismo es vá li do para la mayor parte de los ne ur otrans misores amínico s. Tambi én es ca racte ríst ic o de los neurotransmisores q ue actúan sobre los recep tores metabotrópicos, que se liberan en forma extrasináptica, es decir de zonas del plas ma lema ter mi nal fu era de la condensación p re sináptica. Además, los receptores sobre los cuales actúan también son de localización extrasináptica, ya sea cerca de la condensación postsi náptica o a una distancia considera bl e. Por lo tanto, las moléculas de transmisores deben difundirse una distancia corta o más larga pa ra llegar a los receptores, como es el caso de las aminas libe radas de las varic os idades de las fi bras nerviosas aminérgicas (v éase también la in ervación del músc ulo liso, Capítulo 13, pág. 3 12). Así, en la mayo r ía de los casos, en el encéfalo las vari cosidades de las fi bras aminérgicas no forman contactos sinápticos. La transmisión sináptica lenta con ac ti va ción de los receptores metabotrópicos se denomina
también neuromodulación , dado q ue la acción no crea un potencial de acción en la célu la postsináptica, pero puede modificar su sensibi li dad f re nte a otras acciones de corta duración mediadas p or transmisión s in áptica rápida. Antes de demostrarse su presencia en el sistema nervioso central, muchos de los neuropéptidos eran conocidos como hormonas con efecto fuera del encéfa lo , por ej emplo, las hormonas del tubo digestivo gastrina y colecistocinina (véase con mayor detalle en el Capítulo 18) y las hormonas hipofisari as adrenocorticotrófica y lu teinizante (véase el Capí tu lo 21). Además, el e nc éfalo, el tubo di gestivo y el sistema inmunológico contienen receptores es pecíficos para sustancias similares a la morfina, denominados re c ept ores de opiáceos. Muchos de los péptidos ne uroacti vos endógenos se unen a los receptores de opiáceos y tienen efectos s imila res a los de estos com puestos. En la actualidad, se denomi na endorfinas ("morfina endógena") a este grupo de péptido s, de los cuales dos son pentapéptidos, las encefalinas (gr. , enkephalos, en la cabeza), idénticas salvo el aminoácido terminal. En la MET-encefalina ( met-enk ) el aminoácido termin al es metionina; en la LEU-encefalina (leu-enk), es leucina. Se ha demostrado la presencia de e nc efalinas en las terminaciones nerviosas de los si stemas nerviosos central y periférico, con efecto anal gésico después de su in yección en el tronco del encéfalo. Actúan como ne urotransmisores en un sistema endógeno de inhibición del dolor en el tronco encefá li co y la médu la espina l, donde modu lan la transmis ión de los impulsos dolorosos. Algunas endorf in as más grandes también se unen a los receptores de opiáceos y ti enen propiedades an algésicas. Además de su relación con la modulación o la transmisión del dolor, es probable que las encefalinas y las endor fi nas intervengan en un espectro mucho más amplio de funciones encefá licas, entre ell as , efectos sobre reacc io nes emocionales y sensaciones de placer. Tamb ié n moléc ul as muy pequeñas, ciertos tipos de gases, son una forma no característica de neurotransmisores. Por el momento , esto es válido para el óxido nítrico (NO) (ing. n itric o xi de) , una molécula de vi da media corta. El NO es s in teti zado a partir del aminoácido arginina, en una reacció n ca tal izada por la e nz ima NO-s in tetasa, y se ha demostrado q ue causa dilatación de los vasos sanguíneos. No ha y acumu lación en las vesfculas sinápticas,
Tejido nervioso I 327
BOOK´S COPICENTRO USAC / 2016
dado que el NO se difunde al exterior de la célula a la par de la síntesis (el NO atraviesa sin dificultad el plasmalema). El efecto tampoco se desencadena por unión a receptores tradiciona les en las membranas postsinápticas. El efecto del NO sobre las células musculares lisas es de relajación. Durante mucbo tiempo se supo que este efecto era causado por la acetilcolina, pero ahora se ha demostrado que el mecanismo ocurre a través de las células endoteliales (las células que tapizan la superficie interna de los vasos ; véase con mayor detalle en el Capítulo
- y el NO. Las células endoteliales poseen receptores para aceti1colina de tipo muscarí nico, es decir, receptores metabotrópicos con
COj o Salvo los péptidos, todos son compuestos de bajo peso molecular que se sintetizan en la terminación axónica. En cambio, los péptidos se sintetizan en el retículo endoplasmático rugoso del cuerpo celular y son transportados desde allí por transporte axónico anterógrado. La unión del transmisor con el receptor de la membrana postsináptica modifica la permeabili dad de esta última para ciertos iones, lo cual causa una variación del potencial eléctrico a través de la membrana celular. El efecto se denomina ex citador si el potencial de membrana en la célula postsináptica se hace menos negativo, dado que aumenta la probabilidad de que se inicie un po tencial de acción. Esto ocurre cuando el receptor es un canal iónico de sodio activado por neuro transmisor, puesto que la unión con el neurotrans misor induce la apertura del canal con ingreso de iones sodio y la consecuente disminución del potencial de membrana. Los ejemplos más importantes de transmisión sináptica excitadora de este tipo son el efecto de la acetilcolina sobre el receptor de acetilcolina (de tipo nicotínico) en la placa motora terminal (véase también el Capítulo 7, pág. 197) y el efecto del glutamato sobre los receptores acoplados a canales iónicos en el sistema nervioso central. El glutamato y el aspartato son los principales neurotransmisores excitadores del sistema nervioso central. El efecto del transmisor se denomina inhibi torio si el potencial de membrana se hace más negativo, ya que disminuye la probabilidad de formación de un potencial de acción en la célula postsináptica. Esto ocurre por unión del trans misor a los receptores asociados con canales ió nicos de cloro activados por neurotransmisores. De este modo, se produce la apertura del canal iónico y el ingreso de iones cloro. Los ejemplos
32 I Capítulo 14
proteína G como intermediario. En este caso. el IP3 actúa como segundo mensajero que favorece la liberación de iones calcio desde el retículo endoplasmático liso y la consecuente unión de estos iones a la calmodulina (véase el Capítulo 7, pág. 201). A continuación , el com plejo ca1cio-calmodulina activa la NO-sintetasa que cataliza la síntesis de NO, y éste se difunde a las células musculares lisas adyacentes en la pared vascular. Aquí, el NO activa la enzima guanilato ciclasa, que cataliza la formación de GMP cíclico, el cual , como segundo mensajero en la célula muscular, activa la proteína cinasa C. Entonces , esta enzima produce relajación muscular y dilatación del vaso sanguíneo.
más importantes de transmisión inhibitoria de este tipo son las uniones de GABA y glicina a los canales iónicos de cloro en el sistema ner vioso central. En conjunto, los ejemplos nombrados de transmisión excitadora e inhibidora por unión directa del neurotransmisor a los receptores asociados con canales iónicos se denominan transmisión química rápida, puesto que la transmisión de una señal sólo tarda unos pocos milisegundos debido al mecanismo directo. En cambio, en la transmisión química lenta, la transmjsión de la señal tarda varios cientos de milisegundos y la respuesta dura más tiempo, desde segundos a minutos. Esto se debe a que en este caso son receptores acoplados a proteína G, donde el receptor ejerce una acción indirecta sobre el canal iónico, mediada por la proteína G. Este tipo de receptores activados por neuro transmisores reciben el nombre de receptores metabotrópicos (véase también el Capítulo 7, pág. 199). En muchos casos , hay colocalización de dos neurotransmisores distintos en la misma termi nación axónica, a menudo una amina o un ami noácido más un neuropéptido. Por el contrario, es muy infrecuente la presencia de dos transmisores de bajo peso molecular en la misma terminación, pero hay colocalización constante de GABA y serotonina, entre otros. El aspecto de las vesículas sinápticas varía según el tipo de neurotransmisor que contienen. La mayor parte de las terminaciones axónicas del sistema nervioso central contienen vesículas esféricas de alrededor de 50 nm de diámetro con interior límpido (véase la Fig. 14-12). En ocasiones, en estas terminaciones también se encuentran desde escasas hasta abundantes vesÍ
BOOK´S COPICENTRO USAC / 2016
Factores de crecimiento nervioso
Un ejemplo de sustancia neurotrófica es el fac tor de crecimiento nervioso (NGF) (ing. nerve growth factor), una proteína que entre otros efectos es un estimulante del crecimiento de los axones de células ganglionares simpáticas. El NGF se forma en los órganos diana para los axones y se une a los receptores específicos en las terminaciones axónicas para después ser movilizado por transporte retrógrado hacia el cuerpo neuronal, donde ejerce sus efectos. Existen muchos otros factores de crecimien to nervioso relacionados que ejercen efecto similar sobre el sistema nervioso central. En conjunto, estos factores conforman la familia de las neurotrofinas, de las cuales , además del NGF, en el encéfalo de mamíferos se han identificado el factor neurotrófico derivado del encéfalo (BDNF) (iug. brain derived neurotrophic factor) y dos neurotrofinas de-
La doctrina de la neurona también presupone que la transmisión sináptica siempre es unidi reccional, es decir, desde la célula presináptica a la célula postsináptica. Esto ocurre en casi todas las sinapsis de los mamíferos, en los cuales la transmisión sináptica es mediada por una sus tancia transmisora química, es decir, en sinapsis químicas. La contrapartida son las sinapsis elec trotónicas (o sinapsis eléctricas en la forma de los nexos descritos en el Capítulo 6). Las sinapsis electrotónicas se encuentran rara ve z en los mamÍ feros, entre las intemeuronas de varias regiones de la corteza cerebral y en la retina (véase el Capítulo 24). Los canales intercelulares de los conexones (véase también el Capítulo 6, pág. 176) permiten el pasaje directo de iones pequeños de una célula a otra, por lo que la variación del potencial de membrana en una célula se difunde a la siguiente. Esto se denomina acoplamiento electrotónico, y el agente mediador es la corriente eléctrica (cabe recordar que, en las células vivas, la corriente eléctrica sólo es transportada por iones pequeños, p. ej. , iones de sodio o de cloro). La transmisión es mucho más rápida que en las sinapsis químicas, dado que sólo depende de la velocidad de los movimientos de los iones pequeños. La diferencia es menos pronunciada en los mamíferos, debido a la elevada temperatura corporal. Por último, a causa de su estructura y su función, la sinapsis electrotónica es bidireccional, es decir, permite la transmisión en ambas direcciones. Una función importante de las sinapsis electrotónicas sería la
330 I Capítulo 14
nominadas neurotrofina 3 (NT-3) Y NT-4. El efecto de los factores de crecimiento nervioso no sólo es retrógrado, sino también anterógrado por traslado a la célula postsináptica; además, los factores tienen efecto autocrino sobre la propia célula. En este sentido , parece que tienen importancia en el mantenimiento de las neuronas, también en el sistema nervioso del adulto. Cada neurona recibe influencia de varios de los distintos factores de crecimiento y, a la inversa, los efectos de los factores no se limitan a un único tipo de neurona. El interés por los factores de crecimiento recibe fuerte estímulo de las evidencias experi mentales que sugieren la posibilidad de que los factores de crecimiento podrían ser de valor para el tratamiento de distintas patologías neurológi cas , por ejemplo, la enfermedad de Parkinson y la enfermedad de Alzheimer.
sincronización de la actividad en las neuronas que relacionan. Los nexos que unen las células musculares cardíacas y las células musculares lisas y median el acoplamiento electrotónico entre ellas (véase el Capítulo 13 , pág. 307) son idénticos s en estructura y función a las sinapsis electrotónicas del tejido nervioso.
Neuroglia
El tejido nervioso se compone de neuronas y células de sostén no neuronales denominadas neuroglia (gr. glia, pegamento), cuya cantidad a menudo es muy superior a la de las neuronas. Los nervios periféricos, cuya porción neuronal está representada por axones, poseen además un componente de tejido conectivo que no se encuen tra en el sistema nervioso central, en el cual sólo escasa cantidad de tejido conectivo acompaña los vasos sanguíneos. La neuroglia o glía comprende las células neuróglicas que se encuentran entre las neuronas del sistema nervioso central y el epéndimo que tapiza la superficie las cavidades del encéfalo y de la médula espinal. A menudo, se denomina neuroglia periférica a las células de Schwann de los nervios periféricos y a las células satélite que rodean los cuerpos neuronales de los ganglios espinales (véase la Fig. 14-33) Y de los ganglios de los nervios craneales.
BOOK´S COPICENTRO USAC / 2016
Células de la neuroglia
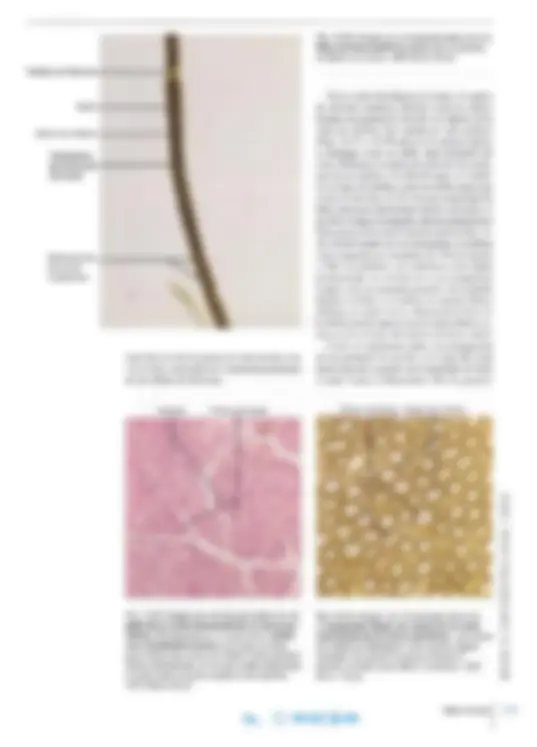
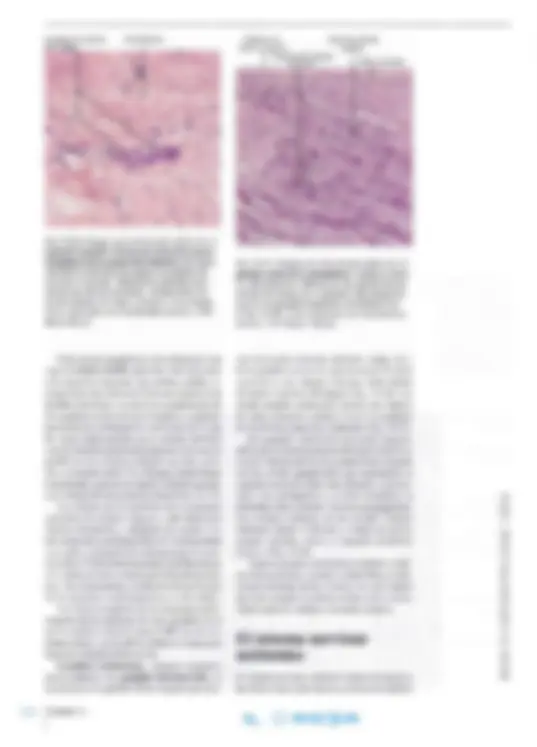
En los cortes histológicos habituales del sistema nervioso central, las células nerviosas y sus pro longaciones siempre están rodeadas por pequeños núcleos dispersos pertenecientes a las células de la neuroglia (véanse las Figs. 14-3 y 14-17). Además, las células neuróglicas pueden esuldiarse por métodos de coloración selectivos mediante los cuales es posible demostrar el cuerpo celular y sus prolongaciones. Se diferencian astrocitos, oligodendrocitos (en conjunto denominados ma croglia) y microglia. Astrocitos. Los astrocitos son células con forma de estrella ( gr. astron, estrella) y numero sas prolongaciones citoplasmáticas (Figs. 14- y 14-19a). Algunas de las prolongac iones están en contacto con un vaso sanguíneo en la forma de pies perivasculares característicos (véanse las Figs. 14-17 Y 14- 19a). El núcleo de los astro citos es más claro que el de los demás tipos de células de la neuroglia, y el ci toplas ma contiene numerosos fi lamentos y grá nu los de glucógeno. Los fi lamentos son de tipo in termedi o, compues tos por proteína ácida fibrilar glial (GFAP) (ing. glial fihrillary acidic prote in ) que sólo se encuentra en los astrocito s. En consecuencia, se utiliza la determinación inmunohistoquímica de GFAP para demostrar la prese nc ia específica de astrocitos (véase la Fi g. 3-54, pá g. 109). El contenido de f il amentos neuróglicos es mu y importante en un o de los ti pos principales de as trocitos, los astrocitos fibrosos (Fi g. 14-18), que se encuentran sobre t od o en la sustancia blanca
Fig. 14-17. Dibujo esquemático de los distintos tipos de neuroglia del sistema nervioso central y sus relaciones con las neuronas y los vasos. La mitad de la izquier da del dibujo muestra el aspecto tras la colora ción con distintos mé todos para neuroglia, mientras que la mitad de la derecha muestra el aspecto tras la tin ción de Nissl (p. ej., con tionina). FA astrocito fibroso; Mi microglia; 01 oligodendrocito; PA astrocito protoplasmá tico. (Según Chandler Elliott).
y presentan menos prolongaciones más largas y menos ramificadas que el otro tipo principal, los astrocitos protoplasmáticos (Fig. 14-18), que se hallan fundamentalmente en la sustancia gris y poseen prolongaciones de forma muy variable. Ambos tipos de astrocitos emiten pies perivas culares y prolongaciones que entran en contacto con las superficies de las neuronas (pies perineu ron ale s ), por ejemplo las terminaciones nerviosas y las dendritas, fuera de las zonas de sinapsis. Además, algunas de las prolongaciones forman procesos pediculares aplanados que establecen contacto con la piamadre (la capa interna de las meninges). Aquí, los procesos pediculares (pies subpiales) están relacionados mediante numero sos desmosomas y nexos y forman en conjunto una capa denominada membrana limitante glial externa o superficial. Una membrana similar se genera a la altura de los pies perivasculares y, en conjunto, estas membranas separan a las neuronas del sistema nervioso central de la piamadre y de los vasos, respectivamente. Antes se creía que la principal función de los astrocitos era mecánica, como "armazón" que separa las neuronas y sus prolongaciones entre sí. Pero se ha demostrado que intervienen en forma mucho más activa en la interacción funcional con las neuronas. Es cierto que no presentan excitación eléctrica (como todas las células de la neuroglia), es decir, carecen de capacidad para generar po tenciales de acción, y se comunican a través de señales químicas. Por lo tanto, son estimulados en forma no eléctrica por las sustancias liheradas
Ci)
(1) 01 0 D^ ~ e Q)^ PA Neurona
¡9 G^ [jj)^^01 El (^) liI Mi (!)
él Q¡ G
t)
S
E) f)
C> (^) o (^) o <::> FAO 01 O o o
o Q,
Tejido nervioso I 331
BOOK´S COPICENTRO USAC / 2016
Fig. 14-19. a Imagen con microscopio óptico de astroglia, demostra da mediante la técnica de sublimación áurica de Cajal. Nótense los procesos pediculares perivasculares. x270. Barra: 20 IJm. (Cedido por J. Zimmer). b Ima gen con microscopio óptico de un oligoden drocito, demostrado por método inmunohis toquimico con anticuer pos contra la proteína Rlp, específica para los oligodendrocitos. x11 OO. Barra: 10 IJm. (Cedido por B. Finsen).
Astrocitos Procesos pediculares perivasculares Oligodendrocito
lactato, que se transfiere a las neuronas a través del espacio extracelular. Las células nerviosas pueden consumir lactato en lugar de glucosa, y algunas neuronas incluso parece que prefieren el lactato a la glucosa como fuente de energía. Los astrocitos y las otras células de la neu roglia no forman sinapsis (como excepción, se han constatado sinapsis neuróglicas en un tipo de precursor de oligodendrocitos). Las células están relacionadas hasta cierto punto mediante nexos, pero estos no actúan como sitios de sinapsis elec trotónicas , dado que no hay potenciales de acción transmisibles. De todos modos. hay indicios de que se produce una comunicación entre los astrocitos a través de los nexos. Cuando se estimula un astro cito, aumenta la concentración intracelular (citosó lica) de iones calcio (por liberación de estos iones desde el REL) , lo cual desencadena la reacción del astro cito a la estimulación. El incremento de la concentración de iones calcio puede difundirse a nivel local en el astrocito y al astrocito vecino a través de los nexos en la forma de una ola de cal cio. Así, la reacción del astrocito a la estimulación puede difundirse a una cantidad muy grande de sinapsis, puesto que un único astrocito puede estar en contacto con más de 100.000 sinapsis. A través de los nexos, los astrocitos conforman un espacio fluido continuo,lo cual incrementa la probabilidad de actuar como amortiguador en la regulación del medio iónico extracelular. Los astrocitos también tienen funciones muy importantes durante el desarrollo del sistema nervioso central, donde aparecen como neuroglia radial, que dirige la migración de las neuronas recién formadas y también dan origen a neuronas y astrocitos. Además, la neuroglia radial o una de sus variantes al parecer intervienen en la neoformación de neuronas en regiones aisladas del encéfalo maduro (más detalles sobre neurogha radial en
histogénesis). También se cree que los astrocitos liberan sustancias tróficas con efecto sobre las neuronas y los oligodendrocitos, además de esti mular la formación de los contactos oclusivos en el endotelio de los vasos sanguíneos y, por lo tanto, el desarrollo de la barrera hematoencefálica (más detalles sobre esta barrera más adelante). En caso de lesión del tejido nervioso, ya sea por traumatismo mecánico o como consecuencia de isquemia debida a trombosis o rotura de vasos, los astrocitos aumentan la captación de los iones y los neurotransmisores filtrados de las neuronas dañadas. Los astrocitos también conforman las células cicatrizales del sistema nervioso. Durante la cicatrización, ocupan los defectos tisulares creados y se hacen más ricas en fibras. En consecuencia, el tejido c.icatrizal suele ser de consistencia más firme que el muy blando tejido del SNC, por lo que la formación de cicatrices se denomina esclerosis (gr. se/eros, seco, duro). Oligodendrocitos. Los oligodendrocitos (gr. oligo." pocos) poseen menos prolongaciones y menos ramificaciones que los astrocitos (véanse las Figs. 14-18 y 14-19b). En los preparados histo lógicos habituales, los núcleos son más pequeños y oscuros que los de los astrocitos; el cuerpo celular también es más pequeño y no contiene filamentos ni gránulos de glucógeno. Los oligodendrocitos satélite se encuentran adosados al cuerpo de las células nerviosas de la sustancia gris (véase la Fig. 14-.17). Los oligoden drocitos satélite no presentan especializaciones ultraestructurales donde están en contacto con los cuerpos celulares neuronales. Es posible que la estrecha relación sea señal del intercambio de sustancias, pero aún no se ha aclarado la función de estas células. Los oligodendrocitos interfas ciculares se encuentran sobre todo en la sustancia blanca (véase la Fig. 14-17) en la forma de hileras
Tejido nervioso I 333
BOOK´S COPICENTRO USAC / 2016
entre las fibras nerviosas. Forman la mielina en el sistema nervioso central. por lo que son homólo gos a las células de Schwann del sistema nervioso periférico (véase más adelante). Microglia. Las células de la microglia son pequeñas, con un núcleo reducido y oscuro, y delgadas prolongaciones con finas espinas (véanse las Figs. 14-18 y 14-20). La microglia se encuentra por todo el sistema nervioso central y es más numerosa en la sustancia gris. Representa el 5-20% del total de las células de la neuroglía en el sistema nervioso central. Mientras que las demás células de la neuroglia son de origen neuroectodérmico (véase la pág. 357), al igual que las neuronas la microglia es de origen mesodérmico. Se generan durante el desarrollo embrionario a partir de monocitos fetales o sus precursores , y llegan al sistema nervioso centr al por el torrente sanguíneo. Los monocitos migrantes sufren posibles divisiones y una diferenciación celular con aparición de prolongaciones y desarrollo de los típicos micro gliocitos residentes. En caso de daño del tejido nervioso, los mi crogliocitos residentes pueden transformarse en microgliocitos reactivos, con fagocitosis activa, que actúan como células presentadoras de antígeno profesionales. La microglia son las primeras célu las que reaccionan en minutos ante una lesión del sistema nervioso central, con divisiones celulares, cambios morfológicos y liberación de moléculas de señal (citocinas). Recién después (quizá indu cidas por las moléculas de señal de la microglia) reaccionan la astroglia y la oligodendroglia. La activación de la microglia es gradual , por lo que la respuesta se adapta al daño. Recién en la muerte celular neuronal la microglia se diferencia a cé lulas fagocíticas activas. Además de la división celular, el aumento de células de la microglia se debe a migración de mono citos provenientes del torrente sanguíneo y al ingreso de microgliocitos desde regiones limítrofes del SNC. Unos días después del inicio de la reacción, se detecta la muerte de algunas de las células de la microglia por apoptosis, entre ellas, las células Ü1gocíticas incapaces de volver al estadio de residentes en reposo, como parece que es el caso de las demás células de la microglia activadas. Este equilibrio entre reclutamiento con proliferación o ingreso desde el torrente sanguíneo y muerte celular por apoptosis también se observa en el sistema nervio so central normal, donde tiene lugar un recambio lento constante de los microgliocitos residentes. En principio, una reacción inflamatoria del sistema nervioso central tiene el mismo objetivo que en cualquier otro tejido, pero transcurre de otro modo. Así, nunca o sólo rara vez se observa ingreso de granulocitos neutrófilos y la reacción
334 I Capítulo 14
Microglia
inmunológica es más lenta y menos evidente. Probablemente sea la expresión de un sistema protector por el cual el t ej ido nervioso no se ve ex pu esto a una acción muy intensa de las sustan ci as cito tó xi cas secretadas por los granulocitos neut ró fi los y los macrófagos activados. La antes mencionada activación gradual de la microglia en relación con el grado de gravedad del traumatismo es una señal similar de protección, dado que el edema relacionado con la inflamación podría te ner rápidas consecuencias catastróficas debido al aumento de la p re sió n intracraneal. Cabe destacar que la mi croglia también produce varios factores neurotró fi cos, entre ellos N GF, lo cual sustenta la supervivencia de l as neuron as. Incluso después de traumatismos importantes, la microgliosis reac tiva disminu ye al cabo de alrededor de un mes. En apariencia, la reacción inflamatoria menos intensa en el sistema nervioso central se debe a la escasa vigil ancia inmunológica, puesto que sólo hay escasos linfocitos T recirculantes entre la sangre y el tejido nervioso (véanse más detalles sobre recirculación y linfocitos en el Capítulo 16). Aquí, los microgliocitos residentes actúan como células presentadoras de antígeno para los linfocitos T. Además, se cree que la menor tendenc ia a desencadenar reacciones inmunológicas en el sistema nervioso centr al se debe a la ausencia de vasos linfáticos que drenen en los ganglios linfáticos regionales. Sin embargo , al parecer los antígenos llegan a los ganglios linfáticos del cuello por drenaje a través del espacio perivascular a lo largo de los vasos que atraviesan la lámina cribosa hasta llegar a la mucosa de la cavidad nasa l. Allí, los antígenos pasan a los vasos linfáticos y luego a los ganglios linfáticos de drenaje. Por esta vía, los antígenos solubles provenientes del encéfalo inducen la producción de anticuerpos en los
Fig. 14-20. Microfoto grafía de microglia de cerebro de ratón, en el que previamente a la extracción del espécimen se provocó una lesión experimen tal. Impregnación con sulfuro de plata. x375. Barra: 20 ].Jm. (Cedido por J. Zimmer).
BOOK´S COPICENTRO USAC / 2016