¡Descarga Células natural killer - NK - y más Apuntes en PDF de Inmunología solo en Docsity!
Células Natural Killer (NK)
1. Introducción
Las células natural killer, células linfocíticas innatas , son un subtipo de ILC del tipo I que no solo desempeñan funciones importantes en las respuestas inmunitarias innatas, sino también representan un nexo muy importante entre la inmunidad innata y la inmunidad adaptativa. Las células NK median mecanismos microbicidas durante las primeras etapas de los procesos infecciosos. Cumplen un papel destacado en: El control de las infecciones virales. La eliminación de células tumorales. Los mecanismos de defensa frente a las bacterias y los parásitos intracelulares. La determinación del perfil en respuesta inmunitaria adaptativa que se articula contra un patógeno o célula tumoral. Las células NK participan en la respuesta inmunitaria a través de dos mecanismos: Producen citocinas y quimiocinas. Destruyen células infectadas o neotransformadas (tumorales) por citotoxicidad o por la activación de receptores de muerte.
La actividad citotóxica y la capacidad de secretar citocinas de las células NK no requieren una exposición previa al patógeno o sensibilización. No presentan receptores específicos para los Ag, ni Igs de superficie. Las células NK pueden destruir a las células recubiertas por IgG. Este mecanismo citotóxico se denomina citotoxicidad celular dependiente de Ac o ADCC y es inducido a través de la molécula CD16 RFcIIIa expresada por las células NK. Constituyen un 5-10% de los linfocitos circulantes y tienen una vida media prolongada. Tienen mayor granularidad y tamaño que los linfocitos. La mayor granularidad de las células NK se debe a la presencia de gránulos cargados de enzimas que participan en la citotoxicidad como la Granzima A, B y la perforina.
2. Desarrollo y diferenciación de las NK
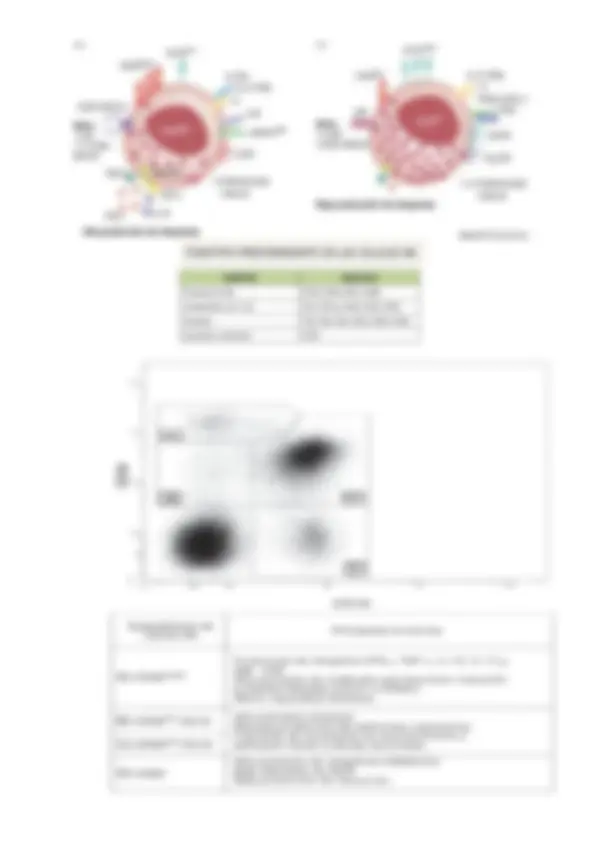
Fenotípicamente, las células NK humanas se identifican por la expresión en su membrana de las moléculas CD56 y CD16, y la ausencia de CD3 y CD4 (marcadores de los linfocitos T). Otros marcadores que caracterizan las células NK humanas son el CD57 y el CD94 pero son menos sensibles. De acuerdo con la densidad con la que expresan la molécula CD56 en la superficie celular se identifican dos subpoblaciones: CD56dim^ CD16bright CD56bright^ CD16dim Las células NK que llegan de la medula ósea a los órganos linfáticos secundarios ya maduros son los CD56bright^ (estadio inicial de diferenciación), allí estos se diferencian en los CD16bright^ (estadio final de diferenciación) que se hallan en la sangre. El 90% células NK tienen una expresión baja de CD56 y se las identifican como CD56dim. Esta subpoblación presenta una alta expresión de la molécula CD16 (CD16bright) y expresa diversos receptores para quimiocinas, lo que le permite migrar a los tejidos Inflamados. Las células CD56dim^ son más granulares que las CD56bright^ por su potencial citotoxico. Residen en la sangre y se extravasan a los tejidos durante la respuesta inflamatoria por un gradiente de quimiocinas inflamatorias entre ellas, la IL-8. El 10% restante de las células NK tienen un fenotipo caracterizado por una alta expresión de CD56 y una baja expresión de CD16 (CD56bright^ CD16dim). Las células NK CD56dim^ median: La actividad citotoxica natural, ya que presentan altos niveles de mediadores de citotoxicidad como las perforinas. La ADCC, porque expresan moléculas CD16. Pero producen bajos niveles de citocinas. Las células NK CD56bright^ tienen escasa capacidad citotóxica porque expresan bajos niveles de mediadores de citotoxicidad, como Granzimas y perforinas. Pero producen altos niveles citocinas inflamatorias e inmunorreguladoras (INF- , TNF- , IL-10, IL-13 y GM-CSF). Residen en los ganglios.
3. Origen, desarrollo y patrón de circulación de las celulas NK
Se originan en la medula ósea a partir de un progenitor linfoide común. La medula ósea es el sitio de mayor relevancia en la maduración de las celulas NK, se originan a partir del precursor linfoide común que expresa el marcador CD34, no expresa ningún marcador de linaje (Lin-) y expresa los receptores tirosincinasas Flt3 y c-kit+.
A medida que estos precursores progresan en su maduración, adquieren la expresión de CD161 y la capacidad de responder a IL-7. La IL-7 es clave para la supervivencia de estas celulas y actúa en conjunto con los ligandos de Flt3 (Flt3L) y de c-kit (c-kit-L), induce la expresión de las cadenas (CD122) y (c) del receptor de IL-2 e IL-15. La cadena es compartida por los receptores de IL-2 e IL-15, mientras que la cadena c es compartida también por los receptores de IL-4 e IL-7. Estas células, bajo el efecto de la IL-15 secretada y transpresentada a través de la cadena del receptor de IL-15 expresado por las celulas estromales de la medula ósea, generan los precursores inmaduros de la celulas NK.
El estímulo ejercido por la IL-15 es un evento crítico en la adquisición del repertorio de receptores de las celulas NK durante su ontogenia, mientras que la expresión de CD122 es un evento crucial, ya que determina que la célula que lo expresa se comprometa al linaje de las celulas NK. El progenitor resultante de la estimulación por IL-15 transpresentada es el que diferencia las celulas CD56bright^ por efecto de la propia IL-15.
El acceso a la sangre periférica y la expresión de los receptores CCR7 y CD62L les permite a estas celulas anidar en los ganglios linfáticos, aun cuando se las encuentra en el bazo. Las celulas CD56bright^ maduran a CD56dim^ en los órganos linfoides secundarios a través de señales impartidas por las celulas dendríticas. Estas señales se producirán tanto por contacto célula-célula como por la acción de citocinas producidas por las celulas dendríticas. Todos estos cambios madurativos se acompañan de una expresión regulada de diferentes receptores presentes en las celulas NK. En la periferia, las celulas CD56dim expresan el receptor para la IL-2 de afinidad intermedia, compuesto por las cadenas y , pero que carece de la cadena (CD25). Las celulas NK CD56bright^ expresan en forma constitutiva el heterotrimero de alta afinidad. La presencia del receptor de alta afinidad permite que concentraciones picomolares de IL-2 logren expandir las celulas NK CD56bright. Mientras la IL-2 expande las celulas NK en los órganos linfáticos secundarios, una segunda citocina, la IL-15, media un poderoso efecto antiapoptotico. ¿Cómo acceden las celulas NK a los tejidos infectados? Las celulas NK CD56dim^ son rápidamente reclutadas hacia los tejidos inflamados o infectados en respuesta a un gradiente de quimiocinas captadas por los receptores CXCR1, CXCR2, CXCR3, CXCR4 y CX3CR1. Por el contrario, las celulas NK CD56bright^ tienen una tendencia mucho menor a ingresar en los tejidos inflamados y migran a los órganos linfoides secundarios, ya que expresan L-selectina y el receptor de quimiocinas CCR7.
4. Activación de las células NK
Pueden activarse mediante dos mecanismos de diferente naturaleza: Ligandos expresados sobre la superficie de las células infectadas o tumorales, que son reconocidos por receptores activadores de la NK. Ligandos expresados sobre la superficie del NK que interactúa con receptores de muerte en las células diana. Citocinas y quimiocinas inflamatorias secretadas principalmente por macrófagos y células dendríticas. Ligandos de receptores Toll y NOD (PAMPs). Las NK expresan TLR1, 2, 3, 4, 7, 8 y 9. Las células NK pueden ser activadas por varias citocinas que incluyen los interferones de tipo I (IFN- e IFN-β), IL-2, IL-15, IL-12, IL-18, IL-21 y C5a. En particular las citocinas IL-12, IL-15 e IL-18 liberadas por monocitos, macrófagos y células dendríticas durante las primeras etapas de los procesos infecciosos inducen la activación de las células NK estimulando la producción de citocinas, fundamentalmente, el IFN-γ, el TNF-α y las quimiocinas inflamatorias, como MIP-1y RANTES. Los interferones de tipo I son secretados principalmente por las células infectadas. El segundo de los mecanismos conducentes a la activación de las células NK involucra su contacto con la célula diana y es mediado por receptores que le permitirán distinguir las células infectadas o neoplásicas de las células normales.
TNF-
5. Receptores expresados por células NK
5.1 Receptores activadores e inhibidores
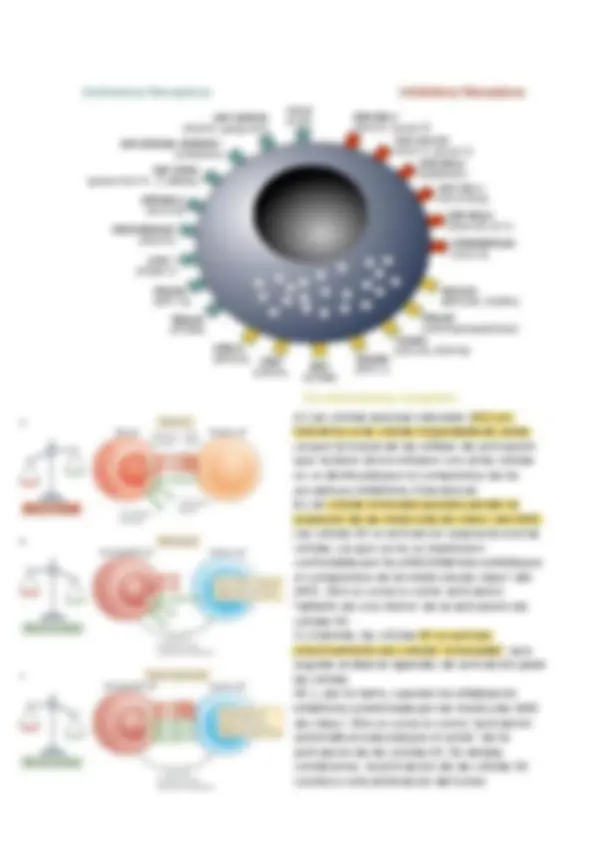
Las células NK expresan una diversidad de receptores. Alguno de ellos son receptores inhibitorios y disparan señales intracelulares que previenen la activación de las células NK. Otros, en cambio, los receptores activadores , disparan señales intracelulares que conducen a la activación de las células NK. Cuando una célula NK "sensa" a una potencial célula diaria, se establece un equilibrio entre señales inhibitorias y activadoras que determinara, si la célula NK se activara, secretara citocinas y desarrollara su capacidad citotoxica antimicrobiana y antineoplásica (desgranulación), o no. En ausencia de procesos infecciosos o neoplasicos, las células NK reciben mayoritariamente las señales inhibitorias de parte de las células con las que establecen contacto. Ello previene su activación. Por el contrario, ante la presencia de células
infectadas o células neotransformadas (tumorales), el equilibrio establecido entre las señales inhibitorias y estimuladoras se inclina a favor de estas últimas, promoviéndose la activación de la célula NK. Esto puede ocurrir mediante: Una expresión aumentada, en la célula diana, de ligandos para los receptores activadores de las células NK. Una expresión disminuida, en lo célula diana, de ligandos para los receptores inhibitorios de las células NK. Los ligandos de los receptores inhibitorios mejor conocidos son las moléculas de clase 1 del CMH. Su reconocimiento por parte de los receptores inhibitorios evita que las células NK destruyan las células normales. En cambio, las células infectadas o neoplásicas, que suelen expresar menores niveles de moléculas de clase I del CMH (como consecuencia de la infección o la neotransformación) desencadenan una señalización intracelular de menor intensidad, lo que permite que prevalezcan las señales de activación y en consecuencia se active el NK. Las células NK realizan un control permanente de los niveles de expresión de las moléculas de clase 1 del CMH en las células del organismo. La activación de la célula NK se promueve cuando ésta se enfrenta a una célula que expresa bajos niveles de moléculas de clase 1.
Los receptores inhibitorios presentan una secuencia común en su cola citoplasmática, denominada secuencia ITIM ( inmunoreceptor inhibidor basado en tirosina ). Después que el receptor que porta esta secuencia reconoce a su ligando específico expresado en la superficie de las células diana, el motivo ITIM se fosforila en el residuo de tirosina, y se dispara una cascada de desfosforilaciones mediadas por fosfatasas, que bloquea la cascada de señalización que media la activación de las células NK.
NK
A) Las células asesinas naturales (NK) son tolerantes a las células hospedadoras sanas, ya que la fuerza de las señales de activación que reciben al encontrarse con estas células se ve disminuida por el compromiso de los receptores inhibitorios (tolerancia). B) Las células tumorales pueden perder la expresión de las moléculas de clase I del MHC. Las células NK se activan en respuesta a estas células, ya que ya no se mantienen controladas por la señal inhibitoria emitida por el compromiso de la molécula de clase I del MHC. Esto se conoce como activación 'faltante de uno mismo' de la activación de células NK. C) Además, las células NK se activan selectivamente por células "estresadas", que regulan al alza los ligandos de activación para las células NK y, por lo tanto, superan la señalización inhibitoria suministrada por las moléculas MHC de clase I. Esto se conoce como "activación automática inducida por el estrés" de la activación de las células NK. En ambas condiciones, la activación de las células NK conduce a la eliminación del tumor
directamente (a través de la citotoxicidad mediada por las células NK) o indirectamente (a través de la producción de citoquinas proinflamatorias, como el interferón-).
5.2 Receptores de muerte
Estos receptores se expresan en las células cuando están infectadas o son tumorales. La activación de estos receptores por la interacción con sus ligandos, lleva a la célula a morir por apoptosis. Los receptores de muertes son FAS y TRAIL. Los ligandos para FAS y TRAILR se hallan en las células NK y son el FASL y el TRAIL respectivamente. Por otro lado los receptores para IFN-γ y TNF-α expresados en las células infectadas o neoplásicas también estimulan la muerte de la célula diana. Sus ligandos, el IFN- y el TNF-, son liberados por las células NK activadas.
5.3 Otros receptores
Los NK también expresan receptores para citocinas, quimiocinas, factor de crecimiento, TLR, NOD, NLR y RIG-
6. Mecanismos que provocan la citotoxicidad de las NK
Las células NK median su actividad citotoxica a través de dos mecanismos:
Vía secretoria, exocitosis del contenido de sus gránulos, en el cual interviene la acción de los receptores activadores, o el contacto con IgG unida a un Ag de la superficie de la célula diana. Las células NK desgranulan produciendo apoptosis o necroptosis. Vía independiente de caspasas. Activación de los receptores de muerte en la célula diana en la cual se activa la apoptosis independiente de gránulos. Un mecanismo similar ocurre cuando el IFN- y el TNF- se unen a sus receptores en las células afectadas. Vía dependiente de caspasas.
Necroptosis
celulas diana. Este ligando es la molécula CD95 (Fas). La microagregacion de Fas inducida por FasL induce la apoptosis de la célula diana por un mecanismo que consta de los siguientes pasos:
**- Trimerizacion de la molécula Fas sobre la membrana de la célula diana.
- Reclutamiento de proteínas adaptadoras al dominio de muerte de Fas.
- Activación de caspasas 8 (vía extrínseca de inducción de la apoptosis).**
Las células NK pueden también inducir la apoptosis de las células diana a través de TRAIL empleando un mecanismo similar al descrito para FasL. A diferencia del FasL, que presenta un solo receptor en la célula diana (la molecula Fas), TRAIL tiene cuatro receptores diferentes denominados DR4 (también conocido como TRAIL-R1 o APO-2), DR5 (o TRAIL- R2), DcR1 (TRAIL-R3) y DcR2 (TRAIL-R4). Sólo DR4 y DR5 son capaces de inducir apoptosis como consecuencia de su trimerización inducida por TRAIL. En cambio, DcR1 y DcR inhiben la inducción de apoptosis gatillada a través de DR4 y DR5, ya que reconocen a TRAIL pero carecen de dominios intracelulares de activación, por lo tanto, secuestran a TRAIL, previniendo su reconocimiento por DR4 y DR5.
7. Interacción entre las celulas NK, las celulas dendríticas y los
macrófagos
Las celulas NK son capaces de modular el curso global de la respuesta inmunitaria innata y adaptativa, es relevante la producción de IFN- por las celulas NK. De hecho, las celulas NK es la primera fuente de IFN- en el transcurso de los procesos infecciosos. Están presentes en la sangre y en los órganos linfáticos secundarios, pero no infiltran los tejidos periféricos sanos. Frente a un proceso infeccioso, acceden al tejido infectado en respuesta a un gradiente de quimiocinas inflamatorias, la IL-8 (CXCR1). En los tejidos, las celulas NK reciben señales de activación impartidas por tres tipos diferentes de estímulos:
- Ligandos expresados sobre la superficie de celulas infectadas, celulas tumorales, macrófagos o celulas dendríticas, que son reconocidos por receptores activadores.
- Citocinas inflamatorias secretadas localmente, sobre todo por macrófagos y celulas dendríticas: TNF-, IL-12, IL-15, IL-18 e IL-21. Frente a las infecciones virales, las celulas NK son estimuladas por los interferones de tipo I (IFN- e IFN-) secretadas por las celulas infectadas y las celulas dendríticas plasmocitoides.
- Ligandos de receptores Toll (PAMP). Las celulas NK expresan TLR1, TLR2, TLR3, TLR4, TLR7, TLR8 y TLR9, lo que le permite responder a diferentes PAMP.
Las células NK no solo interactúan con las células dendríticas a fin de promover su activación; ejercen “un control de calidad” sobre las propias células dendríticas. Aquellas células dendríticas que no logran madurar son destruidas por las células NK a través de un proceso que involucra el reconocimiento, por parte de los receptores Nkp30 y CD (DNAM-1) , de ligandos expresados en las células dendríticas. Este proceso se denomina “edición” de células dendríticas por las células NK. Así, las células NK eliminan células
El IFN- secretada en el foco inflamatorio por la célula NK media dos funciones importantes: activa a los macrófagos y promueve la maduración de las celulas dendrítica. Al activarse y madurar, las celulas dendríticas producen IL-12, IL-15, IL-18 e IL-21. Estas citocinas estimulan la activación de las celulas NK, estableciéndose así un circuito bidireccional inflamatorio. La estimulación de las celulas NK por los macrófagos y las celulas dendríticas desempeña un papel importante en la inmunidad contra diferentes virus, contra bacterias y contra parásitos.
dendríticas que podrían mediar respuestas tolerogenicas frente a los agentes infecciosos. Las células dendrítica maduras son resistentes al ataque citotóxico de las NK debido a que incrementan la expresión de la molécula HLA-E, la cual desencadena señales inhibitorias en las celulas NK a través de CD94/NKG2A. En el propio foco inflamatorio, se establece un circuito de activación reciproca que promueve el desarrollo de células NK con funciones efectoras eficaces y una adecuada maduración y control de calidad (edición) de las célula dendríticas, que adquieren la capacidad de migrar a los ganglios linfáticos para dar inicio a la respuesta inmunitaria adaptativa. ¿Qué pasa mientras tanto en los ganglios linfáticos drenantes del sitio de infección? Se establece una comunicación entre las células NK y las células dendríticas. Las células NK son activadas por acción de las citocinas inflamatorias producidas por las células dendríticas. Asimismo, durante las infecciones virales existe una contribución a la activación de las células NK por parte de los IFN de tipo I secretados por las celulas dendríticas plasmocitoides. Esto provoca la activación de las NK y el desarrollo de una alta capacidad citotóxica y secretoria de IFN-. El IFN- desempeña un papel al promover la producción de IL-12 por las celulas dendríticas y el desarrollo de respuestas Th1 y respuestas mediadoras por los linfocitos T CD8+ citotóxicos. En el mismo ganglio linfático, aquellas células dendríticas que no hayan madurado adecuadamente serán objeto de la actividad citotóxica de las células NK (edición). La participación de NKp30 y CD226 (DNAM-1) no se restringe al proceso de edición de las células dendríticas. Al reconocer los ligandos presentes en la superficie de las células dendríticas, NKp30 y CD226 promueven la producción de IFN- por las células NK. En relación con la acción antitumoral de las celulas NK, estaría mediada tanto por la producción de IFN- como por la acción citotóxica ejercida sobre las propias celulas tumorales. Esta acción citotóxica media dos acciones importantes: a) reduce el número de celulas tumorales viables y b) libera Ag tumorales que son capturados por las celulas dendríticas y promueve el desarrollo de respuestas T CD8+ citotóxicas. Al igual que los macrófagos, las células NK son sensibles a los efectos inmunosupresores del TGF- y la IL-10. La secreción de estas citocinas contribuye al silenciamiento de las células NK y a la restauración de la homeostasis en esta población celular.