¡Descarga Dominios del núcleo celular y más Apuntes en PDF de Biología Celular solo en Docsity!
BIOLOGÍA CELULAR TEMA 7: NÚCLEO CELULAR Lucía Sánchez S-M
El núcleo no tiene por qué ser esférico, posee envoltura y unos determinados componentes o zonas, tales como el nucléolo. En una tinción convencional se observa que en los núcleos se observan masas de cromatina más o menos compactada, áreas desprovistas de tinción y algo que suele ser muy evidente suelen ser los nucléolos. Esta visión es muy simplista.
1. DOMINIOS DEL NÚCLEO CELULAR
El núcleo aparece rodeado por la envoltura nuclear , formada por dos membranas, una interna y otra externa. A lo largo de la envoltura hay complejos de poro , lugares donde se produce un transporte núcleo-citoplasma. Por debajo de la membrana nuclear interna tenemos la lámina nuclear. Se encuentran masas de cromatina compactada y otras masas más laxas. Entre las masas de cromatina hay unos espacios con nucleoplasmas. El conjunto de esos espacios entre masas de cromatina se van a llamar dominio intercromosómico que suele estar a continuación de complejos de poro, para que pueda ocurrir un transporte. Entre otras, las funciones del núcleo son: maduración de RNAs, replicación de DNA, reparación de DNA y la transcripción.
A parte de esto, el nucléolo es una estructura prominente en el núcleo, hay un número de nucléolos determinado para cada célula. Normalmente en el dominio intercromosómico se observan cuerpos, algunos son exclusivos de células tumorales como los PML. Aunque hay otros, los cuerpos de Cajal (que pueden o no estar cerca del nucléolo) y también los Speckles.
En el núcleo su contenido es dinámico y está estructurado en diferentes dominios.
2. ENVOLTURA NUCLEAR
La envuelta nuclear está formada por una membrana externa y una membrana interna , entre las cuales se encuentra un espacio perinuclear o cisterna perinuclear (mejor denominado espacio). Interrumpiendo a estas dos membranas están los complejos de poro , y por debajo de la membrana interna habrá un entramado de filamentos formando la lámina nuclear. La membrana externa de la envuelta es continua con el RER, y sobre ella pueden aparecer asociados ribosomas.
a. Complejos de poro
El complejo de poro es una estructura ordenada formada por 8 pilares que se repiten dejando un hueco en medio por donde se produce el transporte. Las 8 estructuras se repiten, en composición también, son anillos proteicos que forman una canasta nuclear.
Las proteínas que forman parte del complejo de poro se llaman nucleoporinas , en mamíferos se han descrito alrededor de 30. Hay unas que están presentes en los filamentos citosólicos, otras forman parte de la canasta hacia el nucleoplasma. Estas algunas de estas estructuras que se repiten del lado interno al externo.
b. Lámina nuclear
Grosor variable entre los 10 y los 20 nm, compuesta por un material de densidad electrónica intermedia. Está formada por un entramado de filamentos intermedios , formados por unas proteínas laminas , que están compuestas por unos péptidos
lineales con cabezas globulares que se asocian dos a dos formando dímeros que se asocian cabeza con cola y se forman filamentos longitudinales. Estos filamentos se pueden asociar paralelamente entre sí formando unos filamentos más gruesos pasando a denominarse protofilamentos, y tras sucesivos ensamblajes serían filamentos. Estos filamentos comienzan a entrelazarse. Las laminas son filamentos intermedios de tipo V. En cuanto al ensamblaje tiene una forma muy parecida al resto de filamentos, pero se diferencia por función y por dinámica.
i. Tipos de laminas
En mamíferos existen 3 genes que codifican para laminas. Los mensajeros de estos genes sufren fenómenos de splicing alternativo y originan a distintos tipos de laminas. Estos genes son:
Gen LMNA : Laminas A, A(Δ)10, C1 Y C o C1 y C2 sólo están presentes en meiosis, en células germinales Gen LMNB1 : Lamina B Gen LMNB2 : Laminas B2 y B
El primer gen en células diferenciadas y los dos últimos genes en todas las células.
Todos los núcleos tienen algún tipo de lamina B y en algunos también van acompañadas con algún tipo de laminas del gen LMNA. Vamos a pensar que al menos hay 2 tipos de laminas.
ii. LAPs (Lamin-associated proteins)
Las LAPs son una familia de proteínas que suelen aparecer como proteínas integrales de la membrana nuclear interna que se asocian a laminas (proteínas de asociación a laminas). Las encontramos en asociación de la membrana nuclear interna y también en el interior del núcleo, suelen aparecer formando pequeños grumos, tanto las laminas como las LAPs.
Las LAPs periféricas y nucleoplasmáticas se asocian además de con laminas con:
Histonas y fibras de cromatina organizadas Proteínas de silenciamiento Factores de transcripción o complejos de transcripción (DNA-polimerasa II) Actina, miosina y otras proteínas de asociación al citoesqueleto.
v. Patologías asociadas a la envoltura nuclear
Hay distintas patologías que responden a proteínas que forman parte de la envoltura nuclear. Hay mutaciones, por ejemplo en las laminas A o C (fundamentalmente en A). Estas mutaciones se corresponden con patologías muy concretas: laminopatías.
Algunas proteínas LAPs que se asocian a la lámina periférica desde la membrana nuclear interna se verán afectadas (LAP2, Emerina, Aladina, MAN1 y el receptor de la lamina B). La afectación parte de mutaciones en determinadas laminas o proteínas que se asocian que pueden llegar a desencadenar la patología
Las laminopatías son variables. Algunos tipos de mutaciones inducen patologías a nivel de sistema muscular estriado (corazón incluido), en otros casos a la distribución del tejido adiposo, en otros al sistema nervioso periférico. Hay un caso muy particular donde se ve afectado todo el organismo, el síndrome de Hutchinson-Gilford o progeria , el individuo afectado tiene una talla menor de la normal y problemas de alopecia y de envejecimiento prematuro. Suelen morir por problemas cardiacos. Se descubrió como laminopatía estudiando la distribución de una lamina periférica, los núcleos de las células estarán lobulados y tendrán muchos problemas, además se analiza la distribución de territorios cromosómicos ya que la posición estaba alterada.
Hoy en día se sabe que los individuos tienen deleciones en una determinada región del gen que va a codificar para laminas de tipo A, que no se termina de procesar (identificación de la causa genética). Si son mutaciones de otra forma de lamina A no produce patologías tan exageradas. Por el momento sólo hay tratamientos paliativos.
Al tener los dominios cromosómicos alterados, también está alterada la posición de la heterocromatina. Hay cambios en la morfología de los núcleos y surgen blebs. Si la lamina A no es muy funcional, hay condensación de cromatina en la propia envoltura, la estabilidad del núcleo se pierde y hay proteínas que alteran su disposición.
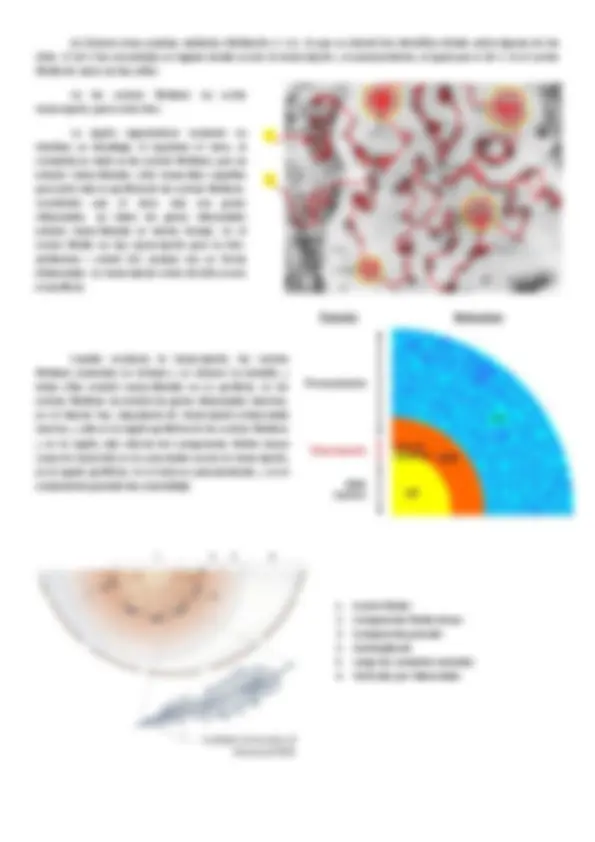
3. MATRIZ NUCLEAR (Nucleoesqueleto)
Después de extraer proteínas solubles y DNA en el núcleo queda una estructura muy prominente, el nucléolo , eminentemente proteico y también rico en RNA, aunque también aparecen cuerpos nucleares más o menos densos y estructuras filamentosas proteicas , que podrían ser RNA pero son filamentos proteicos que forman un entramado en el núcleo.
En microscopía electrónica y con inmunodetección sobre las matrices nucleares, se ve que las laminas forman parte de este entramado filamentoso, que seguramente se corresponde a un entramado que aparece en el nucleoplasma y a los agregados foci. Se obtuvieron anticuerpos contra actina, y fundamentalmente marcaba los núcleos en agregados grandes y amplios. Como para la formación de F-actina se necesita muchas proteínas ( ABPs , actin binding proteins ), se encontraron proteínas de asociación a la actina a la vez que está en la actina. Se localizaron filamentos de actina intranucleares.
Funciones de los filamentos de actina intranucleares: intervienen en la mayoría de procesos que suceden en el núcleo
Replicación del DNA: Si se retiran la célula se detiene en S Transcripción del DNA: Se detiene la transcripción en los 3 tipos de polimerasa que tenemos Reparación del DNA Tráfico núcleo-citoplasma
Viendo estos resultados, deberíamos pensar, que la actina intranuclear tiene funciones especiales respecto a las que tiene en el citoplasma.
Algunas de estas moléculas que forman parte del citoesqueleto intranuclear, o las proteínas asociadas, podrían formar estructuras filamentosas sobre las cuales se engarza la cromatina (actinas, laminas). Entre estas estructuras filamentosas y la cromatina podría haber proteínas que permitiesen la asociación de la cromatina y el citoesqueleto. En general se dirá que la cromatina aparece asociada a un núcleo-esqueleto. Es un modelo, está hablando de fibras de cromatinas de 30 nm de grosor, y por otro lado nos comenta que se podrían estar formando bucles de cromatina en el núcleo.
Se han identificado unas regiones de asociación a la matriz, MAR , son regiones ricas en pares de base AT, es el lugar donde se asocia al citoesqueleto formando bucles.
4. CROMATINA INTERFÁSICA
En el núcleo en interfase, los cromosomas estarán mayoritariamente descondensados, aunque puede haber regiones más o menos condensadas. Dependiendo de la actividad transcripcional, la cromatina estará más o menos compactada, a mayor transcripción, menor compactación. En general se puede decir que en núcleos intermedios, las masas de cromatina que está en la periferia suele estar silenciada, está más condensada, curiosamente asociadas a la envoltura nuclear.
En la formación de la fibra de cromatina de 30 nm se obtiene la estructura de solenoide. Se puede replegar a su vez y rendir fibras de cromatina con una anchura mayor. La fibra de 2 nm también puede encontrarse en el núcleo en los genes ribosomales (nucléolo) o en DNA en transcripción, que estará en fibra desnuda sin histonas. También encontraremos fibras de 10 nm. Así, dependiendo del área del núcleo y de la actividad transcripcional se puede encontrar distintos niveles de compactación de cromatina, aunque la fibra mayoritaria es la de 30 nm, forma en la que al DNA se le acoplan proteínas, histónicas y no histónicas, hablándose ahora de cromatina.
Se ha propuesto que la fibra de 30 nm esté formando bucles ( rosetas de cromatina ), estando en la base de todos los bucles las MARs (regiones de anclaje a la matriz). A lo largo de los bucles, que tendrán dimensiones variables, podemos encontrar varios genes, y sobre estas fibras que estarán formando el bucle, pueden estar asociadas distintas proteínas no histónicas y que no forman parte del núcleo-esqueleto ( scaffold , proteína no histónica). Las regiones MARs se asocian y están ancladas al nucleoesqueleto a través de proteínas que podrían servir de unión entre las regiones MARs (regiones sin histonas) y el núcleo-esqueleto. La cromatina en interfase puede tener diferentes dimensiones pero cuyas bases MARs están asociadas a la matriz nuclear.
5. TERRITORIOS CROMOSÓMICOS
Un territorio cromosómico será el volumen que ocupa un cromosoma en interfase. Mediante técnicas de pintado cromosómico o painting , se observó que había cierto orden en el núcleo.
Los cromosomas homólogos están separados en el núcleo, por norma, formando un ángulo mayor de 90º ocupando posiciones extremas.
a. Speckles
Un factor o proteína implicada en el procesamiento de mRNA no estará disperso de forma homogénea por todo el núcleo, sino concentrado en determinadas regiones, por lo que el procesamiento no ocurre en todo el núcleo. Estas regiones son los speckles , aparecen en áreas donde hay poca cromatina, en el dominio intercromosómico.
Cuando se emplean los mismos anticuerpos que identifican los speckles pero sobre cortes, se ve que dentro de los núcleos aparecen regiones donde hay una mayor densidad de granos, lo cual responde a lo que se ve en fluorescencia. Se había descrito en el pasado que en interfase en regiones con poca cromatina había granos denominados gránulos intercromatínicos. No podrían ser partículas pre-ribosómicas. Estos grupos de gránulos intercromatínicos se conocen hoy como speckles, y recogen factores implicados en el procesamiento. La dinámica de estos cuerpos en el núcleo es relativamente fija al núcleo-esqueleto.
Cuando se hace una inmunodetección de los núcleos que están transcribiendo activamente, después de bloquear la transcripción, siguen apareciendo en el núcleo que hay menos grumos y son más grandes. Se llega a la conclusión de que los complejos de procesamiento en general se almacenan en los speckles. Los complejos ribonucleoproteicos que intervienen en el procesamiento están relacionados con los speckles.
Los snRNA se obtienen en el núcleo, se les asocia unas proteínas para exportarlos al citosol, de forma que este complejo snRNA con proteína atraviesa el complejo de poro y la proteína transportadora se suelta dejando el snRNA en el citosol. En el citosol se sintetizan una serie de proteínas que se asocian al snRNA para formar un complejo ribonucleoproteico ( RNP ) que aún no estará maduro, que se importa al núcleo hasta que llega al nucleoplasma.
b. Cuerpos de Cajal
En los núcleos se observan unas estructuras muy densas, redondeadas, que aparecen muy frecuentemente. Inicialmente fueron denominados cuerpos accesorios. Son estructuras muy similares, mucho menos numerosas que los speckles y aparecían en todas las células animales y vegetales. A veces, según el tipo celular, estas estructuras aparecen cerca o aislados al nucléolo. Posteriormente se denominaron coiled bodies , y dentro de este cuerpo había zonas más gruesas, haciendo una reconstrucción, parecía un filamento retorcido. Ahora se sabe que no son así, se encontró una proteína coilina que aparece muy condensada en estos cuerpos. Por último se denominan cuerpos de Cajal pero la proteína sigue llamándose coilina.
Su dinámica en interfase es oscilar en el núcleo, pero no atravesándolo por completo, sino ocupando determinadas posiciones, estando en el dominio intercromosómico como los speckles.
i. Relación entre speckles y cuerpos de cajal
Los snRNA se sintetizan en el núcleo, son exportados al citoplasma, donde se les asocian proteínas para formar el RNP, aunque inmaduro. Estos complejos parcialmente ensamblados son importados de nuevo al núcleo y se van directamente a los cuerpos de Cajal, donde terminan de madurar. Para almacenarlos, una vez estén maduros, pasan del cuerpo de Cajal al speckle. Cuando la célula está transcribiendo será en el dominio intercromosómico, por lo que cuando surgen los mRNA se necesitará maquinaria para procesarlos y todos los RNP salen y van a lugares donde estará la transcripción y el procesamiento.
ii. Relación entre los speckles y las factorías de transcripción
Las factorías de transcripción se asocian con la RNA-polimerasa II , aparecen de forma que no hay una distribución homogénea, sino que la transcripción ocurre en lugares concretos denominados factorías. En general, estas factorías y los speckles no coinciden, aunque en algunas regiones haya solapamiento. En los speckles hay un almacenamiento que cuando la célula lo requiere viaja a las factorías de transcripción para hacer su función.
Las factorías de transcripción están unidas al núcleo-esqueleto y los speckles estarán cerca de las factorías. La idea es que esa factoría al estar fijada a la matriz nuclear, lo que se desplaza es el DNA sobre la maquinaria.
En las factorías se encuentran los factores necesarios para transcripción, las RNA-polimerasas I y II y sobre esas polimerasas que aparentemente están unidas al núcleo-esqueleto son los genes los que van atravesando los complejos, es la cromatina la que se desplaza, y no al revés. También se ha visto que en la replicación y la reparación del DNA también ocurre en lugares fijos del núcleo.
Realizando inmunodetecciones de periodo S para ver la replicación, dependiendo de si el periodo S es temprano, medio o tardío, también se ve que la replicación también ocurre en determinados sitios concretos, lo mismo para la reparación, estos
b. Composición
Los centros fibrilares pueden tener un tamaño mayor o no, y a veces lo que se ve son áreas que no tienen ni fibras ni gránulos dentro del nucléolo, hablamos de intersticios nucleolares. En su momento, estas áreas inicialmente se denominaron vacuolas nucleolares , pero ese nombre no duró porque no hay ninguna membrana. Lo que hay es nucleoplasma.
Según el tipo de nucléolos, aparece también masas de cromatina condensada sobre la superficie del nucléolo, al conjunto de esa cromatina densa asociada a la superficie del nucléolo, la denominamos cromatina perinucleolar. En ocasiones también encontramos masas de cromatina en el nucléolo, estas masas serán de cromatina intranucleolar.
Componentes del nucléolo:
Componente fibrilar Componente fibrilar denso Componente granular
c. Genes ribosomales
La principal función del nucléolo es como lugar de síntesis de los componentes ribosomales:
Subunidad Pequeña: 40 S, hay un rRNA 18S y 33 proteínas Subunidad Grande: 60 S, hay rRNA 28S, rRNA 5,8S y un rRNA 5S y 49 proteínas
Los genes mayores (18, 28 y 5.8 S)
o En humanos hay unas 400 unidades repetitivas o Ordenadas en tándem o Distribuidas en las constricciones secundarias (NORs) en los cromosomas 13, 14, 15, 21 y 22 o Transcritos por la RNA-polimerasa I en el nucléolo
Mientras que los genes menores :
o En humanos hay unas 800 unidades repetitivas o Ordenadas en tándem o Distribuidas en los cromosomas 1, 9 y 19 o Transcritos por la RNA-polimerasa III fuera del nucleolo
En el nucléolo no se sintetizan los ribosomas, sino pre-unidades ribosomales. Sólo cuando estas subunidades salen al núcleo y sólo cuando encuentren un mRNA se unirán para conformar un ribosoma.
Los genes mayores que codifican para los rRNA 18, 28 y 5.8 S , están inmersos en una serie de unidades que se denominan repetitivas (a partir de un pre-rRNA se obtendrán los 3 rRNA). En humanos, se estima que estos genes mayores están estructurados en unidades repetitivas y que existen unas 400 copias. Estas unidades repetitivas están ordenadas en tándem y localizadas en una región cromosómica que son las que corresponden a regiones organizadoras del nucléolo. En nuestro cariotipo, sólo aparecen en constricciones secundarias o NORs secundarias en los cromosomas 13, 14, 15, 21 y 22. Estos genes serán transcritos por la RNA-polimerasa I Los genes menores codifican para el rRNA 5 S y se estima que hay 800 copias por genoma, estructuradas por unidad que se repite, también ordenados en tándem y en este caso estarán en otros cromosomas, mayoritariamente en el 1, aunque también en el 9 y el 19. No están localizados en constricciones secundarias, sino en regiones de cromátidas que son similares al resto, además de que son transcritos por la RNA-polimerasa III.
La organización de los genes ribosomales mayores son los que van a originar las formaciones que nos recuerdan a árboles de navidad. Habrá una región NTS que no se transcribe (región espaciadora) y se va a encontrar siempre entre los bloques donde se encuentran los genes ribosomales mayores (unidad transcripcional). Tenemos un bloque que corresponde a la región que origina a los rRNA 18 S, otro bloque que transcribe para el 5,8 S y otro para el 28 S. Nos quedan unas regiones en la unidad transcripcional intermedias, que se transcriben pero posteriormente serán eliminadas en el procesamiento de los rRNA.
Habrá un primer bloque después del espaciador (secuencia transcrita externa 1) y luego habrá otras 2 (las secuencias transcritas internas 1 y 2) y la última, que también será secuencia transcrita externa 2. Después de esta última sección habrá otro espaciador NTS que no se transcribe. Esta unidad transcripcional junto con el espaciador en 5’ es lo que forma la unidad repetitiva, en tándem. El rRNA es el 80% de los RNA celulares. No hay núcleolo-esqueleto.
Una región cromosómica extendida contiene genes ribosomales, rDNA, en los extremos estarán las NTS y sobre estos ejes de DNA (genes ribosomales serán la fibra desnuda, no se habla de cromatina) hay complejos de transcripción de RNA- polimerasa I. Para la transcripción también puede ser necesaria una serie de factores de transcripción. Se transcribirán fibras a ambos lados del DNA, desde la más pequeñas hasta la más grande, que no será tan grande como la secuencia de DNA porque hay cierto plegamiento conforme los pre-rRNA se van transcribiendo.
En el extremo de cada una de estas fibras hay complejos de procesamiento, según se van transcribiendo se irán modificando y procesando, serán partículas ribonucleoproteícas, RNPs, pero como están en el nucléolo se denominan snoRNPs y sólo funcionarán aquí.
Se hicieron otras pruebas mediante hibridación in situ , lo que se intentó fue identificar dónde están algunos de los rRNA. El 18 S fue encontrado en lugares donde ocurre la transcripción y el procesamiento, al igual que el 28 S. En el centro fibrilar de nuevo no hay señal.
En los centros fibrilares no existe transcripción, pero existe DNA.
La región organizadora nucleolar en interfase se despliega. Si seguimos el trazo, la cromatina se mete en los centros fibrilares, que no estarán transcribiendo, sólo transcriben aquellos que estén más en periferia de los centros fibrilares. Asumiendo que el trazo rojo son genes ribosomales, no todos los genes ribosomales estarán transcribiendo al mismo tiempo. En el centro fibrilar no hay transcripción pero la RNA- polimerasa I estará ahí, aunque sea en forma almacenada. La transcripción como tal sólo ocurre en periferia.
Cuando comienza la transcripción, los centros fibrilares aumentan en número y se reducen en tamaño, y todos ellos estarán transcribiendo en su periferia. En los centros fibrilares no estarán los genes ribosomales inactivos, en el interior hay maquinaria de transcripción almacenada inactiva, y sólo en la región periférica de los centros fibrilares y en la región más interna del componente fibrilar denso (zona de transición) es la zona donde ocurre la transcripción, en la región periférica. En el resto es procesamiento, y en el componente granular hay ensamblaje.
- Centro fibrilar
- Componente fibrilar denso
- Componente granular
- Nucleoplasma
- Loops de cromatina nucleolar
- Partículas pre ribosomales
e. Funciones
Su principal función será sintetizar ribosomas, los genes ribosomales mayores se transcriben y originan un pre-rRNA (precursor), por lo que se dará un procesamiento posterior. Para ello se necesitan snoRNA y proteínas que formarán complejos RNP, además de otras proteínas endonucleasas. Todas ellas serán importadas y comenzarán con el procesamiento.
Esto ocurre en la periferia de los centros fibrilares y en el componente fibrilar denso. Las proteínas ribosómicas llegan desde el citosol y se empiezan a formar la partícula pre-ribosomal. El rRNA 5 S es transcrito por la RNA-polimerasa III fuera del nucléolo, se importa y va a formar parte de la subunidad mayor.
Las proteínas de las telomerasas se sintetizan en el citosol, se importan al nucléolo, donde con el RNA de la telomerasa (también sintetizado fuera del nucléolo) forman un complejo maduro que es lo que se conoce como telomerasa.
Al final, en la síntesis de partículas ribosomales, lo que se exportan son las partículas, no el ribosoma asociado.
Se ensamblan las subunidades ribosómicas Se ensamblan parcialmente las partículas SRP Se ensambla parcialmente la telomerasa Se procesan algunos pre-tRNAs Se procesa el rRNA 5 S. Se procesa el RNA de la RNAasa P Se secuestran algunas fosfatasas Se almacenan complejos de condensina