¡Descarga EL nucleo (Tema 4) resumen y más Apuntes en PDF de Biología Celular solo en Docsity!
Tema 4. El núcleo
Índice:
● Envoltura nuclear y poros
● Transporte de proteínas
● Lámina nuclear
● Cromatina
● Nucleolo
● Ribosomas
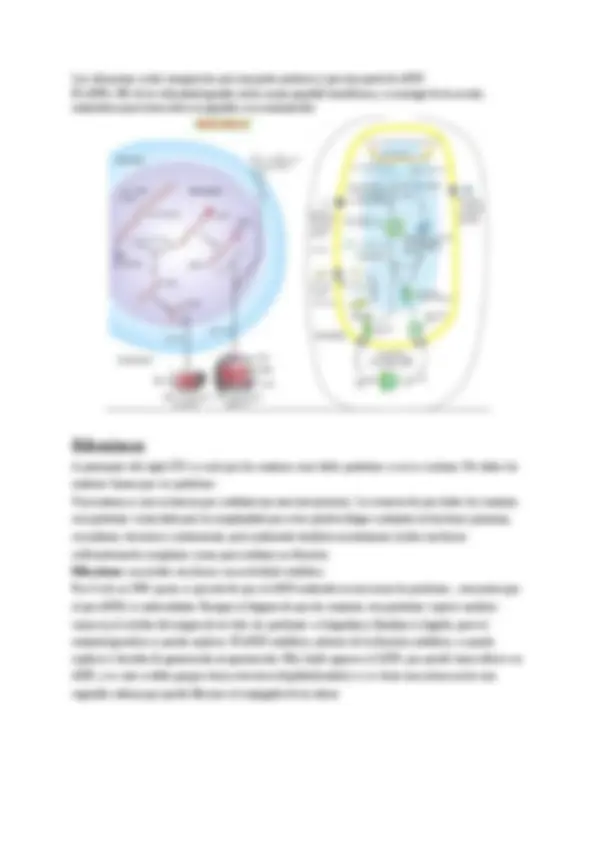
Envoltura nuclear y poros nucleares
La envoltura nuclear está formada por una doble membrana; membrana nuclear interna y
membrana nuclear externa, entre ambas la zona se denomina zona perinuclear. La membrana
nuclear externa se continúa con el RER (también podemos encontrar ribosomas en la
membrana externa). Esta continuidad permite que el espacio intermembranoso y el interior
del retículo endoplasmático se comuniquen directamente y que la envuelta nuclear funcione
también como almacén de calcio.
Ambas membranas a la vez se unen y están separadas por los poros nucleares.
Las criofracturas permiten ver bien las proteínas de membrana o asociadas al poro
nuclear.
Poros nucleares
Son grandes complejos de proteínas que atraviesan la membrana nuclear. En estos
poros, se fusionan ambas membranas.
Tienen una estructura octomérica, grande y una masa de 120000 Da (uma). Son estructuras
formadas por proteínas. Este tiene un diámetro de 120nm en cambio en su interior cuenta con
un diámetro de unos 50nm.
Alrededor del poro nuclear (en la foto) nos encontramos con la heterocromatina (forma de
condensación más alta de la cromatina).
Dónde está el poro hay pocas moléculas ya que el poro es un lugar de tránsito
por donde entran y salen sustancias al núcleo (gracias a los poros se puede
producir el intercambio de moléculas entre el citoplasma y el núcleo, impiden
diferencias osmóticas).
Transporte regulado a través de poros nucleares.
Si una célula necesita importar y exportar muchas sustancias (debido a su tamaño) necesitaremos más poros, por lo que podemos deducir la actividad de una célula por el número de poros que tengan.
**carloscar: es otra forma de expresarlo
Transporte de moléculas en los poros nucleares: El poro tiene una especie de laberinto formado por proteínas. Una proteína pequeña tiene unos 20kDa y una media tiene 40-60KDa. Por lo que si introducimos moléculas de 5000Da o menos pasarán por difusión pasiva. En el núcleo también hay proteínas grandes de 100-200KDa que caben en el poro pero requieren un transportador y se sintetizan en el citosol ( ADNpolimerasa, ARNpolimerasa). Un ribosoma tiene un unos 30-40nm de tamaño y cabría por el poro pero con el enrejado de proteínas no pasa. En los poros se produce una exportación y una importación. Si una célula necesita transportar muchas moléculas del núcleo hacia el citosol o del citosol hacia el núcleo es necesario que el núcleo tenga muchos poros. Por tanto , los poros nos dicen mucho sobre la función de una célula.**
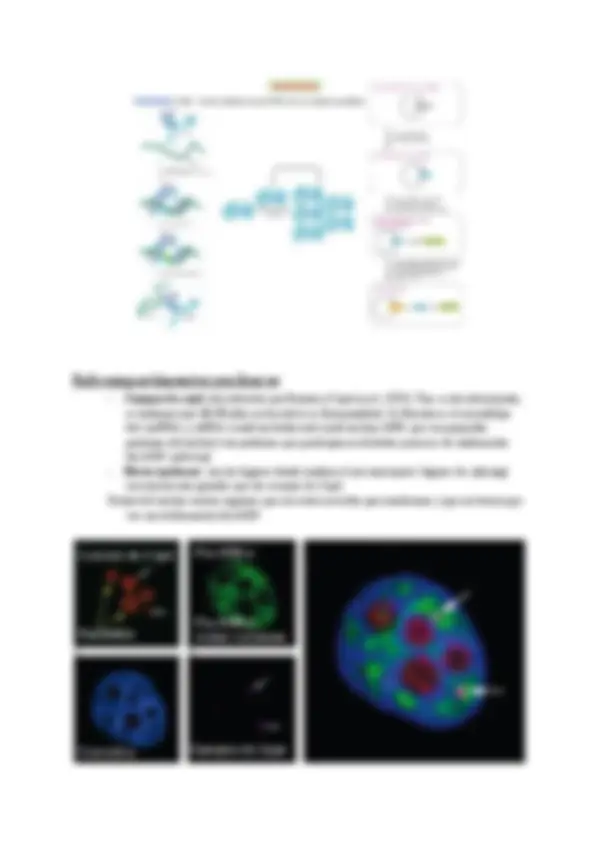
Mecanismo de transporte a través de los poros : Las proteínas que tienen GTP se les conoce como proteínas G(son grandes), también encontramos otro tipo de proteínas G que son proteínas pequeñas y son denominadas “proteínas pequeñas monoméricas (son interruptores moleculares ya que dependiendo de su estado, activan o inactivan procesos). Hay proteínas que cuando se fosforilan se activan o se inactivan. Las proteínas GAP(Proteína Activadora GTP-asa) activan la hidrólisis de las proteínas Ran-GTP(Nuclear que está relacionada con las proteínas Ras(sarcoma de rata)) en Ran-GDP. Cuando nos encontramos una proteína escrita en mayúsculas (ej:RAN, esta proteína cuando muta produce un sarcoma) significa que se estudia en humanos y cuando está en minúsculas en otro ser vivo. Las proteínas GEF(Factor de intercambio de Guanina) transforman el GDP en GTP. Las proteínas GEF están unidas a la cromatina y no pueden salir del núcleo.
En el citosol va a abundar la Ran-GDP y en el núcleo la Ran-GTP. Estas proteínas son un interruptor porque en el núcleo activa una función de una manera y en el citosol activa otra función de otra manera. Para que una proteína entre en el núcleo tiene una determinada secuencia de aminoácidos(secuencia de importación) con carga positiva que significan que esa secuencia pertenece al núcleo. Esa proteína será reconocida por una importina y si hubiese Ran-GTP ésta tendría afinidad por los dos pero como abunda el Ran-GDP se une a la proteína y no hay competencia. Cuando la proteína pasa por el poro a través del laberinto de proteínas y llega al interior del núcleo se libera la proteína porque el Ran-GTP que hay en el núcleo se une a la importina. Cuando la proteína ha sido liberada la importina sale hacia el citosol con la Ran-GTP y por acción de las proteínas GAP se vuelve a transformar en Ran-GDP y pierde la afinidad por la importina por tanto ésta queda libre de nuevo. Las proteínas y las importinas y exportinas pertenecen a las carioferinas, estas interactúan con las proteínas del complejo nuclear, que se denominan nucleoporinas.
Transporte de proteínas
Importación:
La importina es la proteína transportadora que se encarga de transportar ciertas proteínas desde el citosol hasta el núcleo. Es capaz de hacer esto gracias a las concentraciones de ran-GTP ; en el citosol no hay, pero en el núcleo sí. La importina tiene afinidad por la proteína que tiene que transportar, pero tiene más afinidad por el ran-GTP, por lo que al entrar en el núcleo, suelta la proteína y se une al ran-GTP (que abunda en el núcleo). Entonces sale de nuevo al citosol, donde el ran-GTP se hidroliza a ran-GDP por acción de el ran-GAP, porque en el citosol no puede haber ran-GTP.
Exportación:
La proteína que se va a transportar desde el núcleo hasta el citosol tiene que unirse a una exportina que ya tiene unida una molécula de RAN-GTP (que debe estar en el interior). Una vez en el citosol, ocurre la hidrólisis del GTP, formándose el ran-GDP, y la exportina pierde afinidad tanto por la proteína que se estaba transportando como el nuevamente formado ran-GDP. Como la proteína no tiene afinidad por el RAN-GDO, esta proteína que estaba unida a la exportina se suelta de ella queda libre en el citoplasma. La exportina vuelve al núcleo donde se repetirá el proceso.
Diferencias:
constituidos por las proteínas denominadas láminas nucleares. Es el único componente del citoesqueleto que hay en el núcleo.
Está formada por varios tipos de proteínas: A, B1, B2, y C. Originarias de un tipo de proteína B (las primeras, las más relevantes) que evolucionaron, cuya función puede ser unir la la cromatina (normalmente la heterocormatina) a la lámina nuclear.
Funciones:
La lámina nuclear une la heterocromatina a la envoltura nuclear.
Es la responsable de darle la forma al núcleo (diferentes tipos de células tienen un núcleo de una forma u otra).
Es la responsable de la rigidez del núcleo (gracias a las proteínas de tipo A); a algunas células les conviene tener el núcleo rígido y a otras no, por lo que presentarán o no proteínas A. Por ejemplo, los neutrófilos y las otras células (como las endoteliales, que son las que forman las “tuberías” de los
vasos sanguíneos) que pasan por los vasos sanguíneos (por lo que no tendrán proteínas A). Tienen que pasar por un sitio muy estrecho (atravesar las células epiteliales), por lo que adaptan su forma para estar más “delgado” y el núcleo también tiene que caber. Aún así se estropea un poco el ADN (la lámina celular también está involucrada en la reparación del ADN ).
La lámina celular participa en la organización del genoma (la forma que tienen los cromosomas) y en la repartición del ADN.
Las proteínas B también se encargan de la conexión del citoesqueleto con el citosol. En la membrana nuclear interna hay un receptor de lámina B, que ancla la cromatina a la membrana, la lámina celular al citoesqueleto. Esto se debe a que el núcleo puede necesitar moverse o deformarse, por lo que la lámina nuclear se encuentra unida al citoesqueleto para que así este sea capaz de moverlo y deformarlo.
Su última función tiene que ver con la mitosis. Justo después de la profase hay una fase, prometafase , donde desaparece la envoltura nuclear (para que los cromosomas puedan distribuirse y dividirse, por lo que no debe haber envoltura nuclear). Para ello, ocurre la fosforilación de la lámina nuclear; de la cual se encargan las proteínas CDK (son del tipo quinasas)que añaden fósforo a las proteínas de la lámina, haciendo que esta se desorganice y en consecuencia que los cromosomas queden libres.
En la telofase tiene que reorganizarse esta envoltura; por lo que ocurre la desfosforilación de las proteínas de la lámina nuclear. La cromatina está pegada a la lámina celular que comienza a reunirse. La lámina nuclear se reorganiza en torno a los cromosomas , que luego terminarán uniéndose, formando un núcleo.
(otra forma de expresarlo para que quede de dos formas por si de una no se entiende); Una vez llegamos a la telofase de la mitosis desfosforilamos la proteína y por lo tanto estas se vuelven a unir y organizarse de nuevo. Se organizan pegándose a la cromatina, lo que evita que entren otras sustancias. Es decir, la envoltura y lámina nuclear al reorganizarse deben excluir a los orgánulos del citoplasma para que estos no se queden en el interior del núcleo)
Formas de los núcleos:
Algunos ejemplos:
Núcleo de la célula epitelial del epidídimo: tiene una forma muy clara (mucha eucromatina), y muchas invaginaciones nucleares. Esto quiere decir que se transcriben muchas proteínas en este núcleo.
Eritoblasto (célula precursora del eritrocito): tiene mucha heterocromatina, porque necesita mayoritariamente la proteína hemoglobina y poco más, es decir, no necesita transcribir muchos genes para formar a los eritrocitos(al madurarse, el eritoblasto expulsará el núcleo). La heterocromatina está pegada a la lámina nuclear
Neutrófilo: su núcleo es lobulado; cuantos menos lóbulos tienen, más jóvenes son. Esto es debido a lo que se mencionó antes, para atravesar las células epiteliales de los vasos sanguíneos.
Espermatozoide: la cromatina está en su máximo grado de compactación (no tiene que expresar genes); el núcleo sólo está para transportarse hasta el óvulo. Es todo heterocromatina. En lugar de haber histonas para compactar el ADN, hay protaminas (proteínas básicas que se unen al ADN ácido, al igual que las histonas), que se encargan de tal grado de condensación.
Plegamientos de la cromatina:
Las proteínas son las responsables de que el ADN se condense más o menos.
- Doble hélice del ADN (2nm). Nunca se encuentra sola en el núcleo, siempre se encuentran unidas formando nucleosomas.
- Nucleosomas : octámero de ADN, unidos con las histonas: H2A, H2B, H3 y H4. (11nm).
- Nucleosomas compactados en forma de fibra de cromatina de 30 nm, solenoides, al unirse los H1 (30nm). La eucromatina estaría así de condensada, entre los 11 y 30 nm (sobre todo 30 nm). La heterocromatina es una organización de condensación superior que no se sabe exactamente cómo es (se cree que es una sección del cromosoma en forma extendida de 300nm, dominios en bucle).
EN LA MITOSIS se condensan los cromosomas hasta 1400nm.
Side note: siempre tenemos cromosomas, solo están a diferentes niveles de condensación.
Condensinas
Condesinas ≠ cohesinas
Las condesinas forman el “esqueleto”, el eje de los cromosomas. Tienen una estructura/ forma de horquilla y van formando bucles dentro de la cromátida, uniendo, condensando el ADN de una cromátida entre sí. Forman como flores a lo largo de la cromátida. Son del tipo intra.
Las cohesinas unen las cromátidas hermanas entre sí y aparecen durante la mitosis. También tienen forma de horquilla. Son del tipo inter ya que se da entre dos cadenas diferentes.
Territorios cromosómicos
El cariotipo: el conjunto de genes de una especie.
La organización de los cromosomas en el núcleo no ocurre de manera aleatoria y tienen un significado funcional. Se colocan de una forma en función de la necesidad de transcripción de la célula. Los cromosomas más grandes están pegados a la envoltura nuclear. Las regiones que se transcriben están en el centro del núcleo, y las que no, pegadas a la envoltura nuclear, esto muestra la diferencia entre hetero y eucromatina. Los cromosomas interactúan unos con otros, más o menos dependiendo de la célula.
( En color verde están las regiones del cromosoma que se están transcribiendo y las que tienen color naranja es la que si se está transcribiendo. Observando la imagen podemos ver que la heterocormatina (de color verde() se encuentra pegada a la envoltura nuclear))
Enfermedades relacionadas con el núcleo:
Existen enfermedades que afectan a las láminas nucleares (algunas afectan al tejido muscular): laminopatías. Uno de los tipos de estas enfermedades son las progerias , enfermedades que producen un envejecimiento prematuro relacionado con las proteínas de las láminas nucleares.
Centrifugación
Muchas partes de la célula se han estudiado mediante el método bioquímico de la centrifugación. Esta permite estudiar componentes celulares y moleculares según su velocidad de sedimentación, que viene en unidades de Svedberg. La centrífuga permite fraccionar la célula. Para estudiar una parte de la célula primero hay que romper las membranas (ya sea mediante procedimientos mecánicos, ultrasonido, etc.) y se obtiene el homogenado o extracto. Una vez rota la célula y filtrado el homogenado, este se mete en un tubo especial para volverlo a meter en la centrífuga.Cada fuerza de centrifugación separa ciertos componentes de la célula, y cada tubo resultante se denomina de una forma. Primero se pone a 600 g durante 10 min, a esto se llama pellet y se depositan los orgánulos más pesados (el núcleo) al fondo del tubo debido a la gravedad. Si seguimos centrifugando se van quedando abajo los orgánulos más pesados (mitocondrias, cloroplastos…) y se puede ir así discriminando las partes de la célula que queremos estudiar. La unidad de la velocidad de sedimentación es la unidad Svedberg=10-13s
El nucleolo es una estructura del núcleo donde se sintetiza/transcribe la mayor parte del ARN ribosómico que en mamíferos tiene 45s, y que se agrupa con proteínas para formar ribosomas y donde se ensamblan las subunidades ribosómicas. El nucleolo se encuentra en el núcleo y carece de membrana, tienen las proteínas y demás moléculas concentradas de una zona. Además el nucleolo es lo más prominente de la célula. Las células pueden tener más de un nucleolo. Su función es la producción de ribosomas y la síntesis de SRP y telomerasa (enzima que se encarga de alargar los telómeros, que se acortan tras cada división celular).
*El nucleolo desaparece durante la profase mitótica, permitiendo a la cromatina que lo forma reorganizarse para construir los cromosomas. Empiezan a activarse de nuevo al final de la telofase.
Al observar el núcleo de una célula mediante un microscopio electrónico de transmisión distinguimos zonas con distintas densidades (ej: por fuera hay eucromatina), zonas con aglomeraciones de moléculas
Dentro del nucleolo se distinguen 3 regiones, momentos de la fabricación de ribosomas:
● Centro fibrilar claro: La densidad es inferior a la parte fibrilar y granular (más claro). Contiene ADN con histonas y ribonucleótidos, fibras de cromatina y moléculas filamentosas (ARNr). En esta región es donde se encuentra el NOR (y donde se transcribe el ARN 45 pre-ribosomal) y contiene el ADN con las numerosas copias del gen para el pre-ARNr-45S (transcrito primario a partir del cual se obtienen 3 de los 4 ARNr que forman los ribosomas).
● Porción fibrilar densa: es la parte más densa que rodea el centro fibrilar claro y está Está constituida por por ribonucleoproteínas. Aquí ocurren los procesos, son las áreas de transcripción activa u organizadores nucleares. Tiene lugar el procesamiento del transcrito primario pre-ARNr-45s, el corte del transcrito primario en trozos más pequeños. (Parece ser el lugar donde se procesa el ARNr)
● Porción granular: Agrupaciones de gránulos con ribonucleoproteínas. Ocurre el procesamiento tardío del ARNr y se ensamblan las subunidades ribosómicas. Corresponde al ensamblaje del ARNr con proteínas formando ribosomas inmaduros.
El nucleolo tiene procesos característicos de las eucariotas. Por ejemplo. la replicación, la transcripción, la maduración del ARN mensajero y la traducción (tras las salida esepcial por los poros)
SINTESIS DE UN RIBOSOMA ( esto es del peña y está bastante bien explicado, lo pongo por si hay dudas y alguien lo ve mejor de una manera que de la otra)
En el nucleolo es donde se fabrican los ribosomas, que son unidades ribonucleicas que se componen de ribonucleoproteínas, por lo que contienen proteínas y RNAr.
En el nucléolo también se sintetizan otras ribonucleoproteínas (contienen RNA y proteína), como la telomerasa y la partícula SRP (partícula de reconocimiento de la señal).
Un ribosoma se crea a partir de un RNA 45S que es muy grande al principio (45S en mamíferos, 35S en levaduras) y es transcrito en el centro fibrilar claro del nucleolo. La región del cromosoma que codifica ese RNAr preribosomal se denomina región del organizador nucleolar (NOR). En una célula, varias cromosomas pueden contener un RON/NOR. Cada RON/NOR puede contener muchas copias de la región que se transcribe a RNAr pre-ribosomal mediante la ARN polimerasa tipo I, la que las hace más eficientes. Para que madure el ARNr y se ensamble con las proteínas ribsosomales se requieren determinadas proteínas y moléculas de snoARN (small nuclealar ARN).
Este ARN 45S pre-ribosomal madura en el procesamiento del ARN, eliminando algunas zonas que no se empalman (zonas negras de la imagen) y quedando algunas zonas activas, estas son: el RNA 28S y el RNA 5,8S, que forman parte de la subunidad grande, a estos se le une un RNA5s (extranuclear, se codifica fuera del NOR por la ARN polimerasa tipo III) que contribuye a que se forme la subunidad grande del ribosoma pero se crea fuera del nucleolo (Subunidad 60S); y el RNA 18S, que forma la subunidad pequeña (subunidad 40S).
Zonas del nucleolo:
El gen que codifica para la síntesis de ARN 45s está en una parte del cromosoma denominada Región del Organizador nucleolar (NOR o RON), y es en el centro fibrilar claro donde se transcribe el ARN 45s pre-ribosomal. Mientras que es en el centro fibrilar denso donde se ensambla (corta) el ARNr y en el componente/porción granular se ensambla el ARNr con las proteínas procedentes de la importación para formar los ribosomas, inmaduros de momento.
● RON:
En el nucleolo se encuentra una gran cantidad de genes para producir ribosomas, porque una célula necesita una gran cantidad de ribosomas (estas células además de tener muchos genes para producir ribosomas tienen nucleolos muy desarrollados). En el cromosoma humano se encuentra, entre el centrómero y el telómero, la zona del RON. En los cromosomas humanos (acrocéntricos, cromosomas con el centrómero cerca del extremo, pero no el telómero) en fase mitótica encontramos que en los pares 13,14,15,21 y 22 encontramos una región con muchas copias del RON.
Las zonas del RON de cromosomas en interfase se agrupan para formar un nucleolo; por esto, se denomina región organizadora del nucleolo, esto es porque contienen la información necesaria para sintetizar ARNr. Entonces, cuando una célula humana tiene un nucleolo es porque los RON de los diferentes cromosomas lo están formando; en el cromosoma mitótico la zona del RON está condensada y en el nucleolo está descondensado de forma que se pueda sintetizar ARNr. El nucleolo no está formado por membranas, pero tiene compartimentos en los que en cada parte ocurre algún proceso diferente.
Existen hasta 200 copias del gen RON. Las proteínas se sintetizan en el citosol, se exportan al interior y se unen con el ARN obtenido del RON para formar ribosomas que deben ser exportados. El PRE-45sARN usado para formar el ARN que forma parte de los ribosomas se denomina PRE-RNARibosomal (el de las levaduras es 35s).
Para que madure el ARNr y se ensamble con las proteínas ribsosomales se requieren determinadas proteínas y moléculas de snoARN (small nuclealar ARN) que se importan junto con las proteínas AF (factores de ensamblaje), también necesarias. Al principio se forman las dos subunidades ribosómicas juntas y luego se dividen en dos, la grande y la pequeña (las proteínas ribosomales proceden del citosol), también son importantes los factores no ribosomales AF. EL 45s pre-ARN está unido a proteínas (AF).
La gran partícula ribonucleoproteica (ribosoma inmaduro) tiene una velocidad de sedimentación 90s, una vez maduro es 80s