¡Descarga Farmacologia y más Apuntes en PDF de Psicobiología solo en Docsity!
TEMA 1. LES CÈL·LULES DEL SISTEMA
NERVIÓS.
1. LA NEURONA.
1.1 ANTECEDENTS HISTÒRICS
Teoria reticular del SN: Teoria incorrecte que defensa que les neurones estan unides per fibres nervioses i formen una xarxa. (Golgi).
Teoria cel·lular del SN: Teoria que defensa que el SN està format per cèl·lules amb prolongacions que s’apropen entre elles però no contacten. (Ramón y Cajal).
1.2 MORFOLOGIA.
1.2.1 SOMA, DENDRITES I AXÓ.
La neurona està formada per tres parts característiques:
- Cos cel·lular (soma). És la regió que es trova al voltant del nucli. És on són els orgànuls cel·lulars encarregats de la síntesi de proteïnes, el metabolisme,…
- Dendrites. Prolongacions que surten del cos cel·lular. Es ramifiquen a mesura que s’allunyen del nucli. Capten la informació. -
Espines dendrítiques. Petites prolongacions de les dendrites que semblen espines, especialitzades en rebre informació.
- Axó. Format per: IMPULS NERVIÓS
- Cap axónic. Comença al soma.
- (^) Ramificació axónica. Botó terminal. Especialitzat en enviar informació.
*SINAPSIS: espai en el qual 2 neurones estan properes i intercanvien informació.
1.2.2 FIBRES MIELÍNIQUES I AMIELÍNIQUES.
FIBRES MIELÍNIQUES.
Embolcallen i aillen l’axó (mielina). Mielina Capes de membrana cel·lular.
Les fibres mielíniques són diferents al SNC i al SNP.
(SNC Processa la informació. Format per l’encèfal i la medul·la espinal. Protegit per ós.
SNP Permet la comunicación entre el SNC i el cos).
SNP : cèl·lules de Schwann. Embolcallen l’axó. Els axons mielínics són més ràpids en transmetre informació.
Entre les cèl·lules de Schwann es troben unes separacions anomenades nodes de Ranvier.
Node de Ranvier.
- SNC: oligodendròcits (poques prolongacions). Els braços dels oligodendròcits envolten un fragment d’axó i constitueixen la mielina.
FIBRES AMIELÍNIQUES.
Poden ser cèl·lules de Schwann que no cobreixen del tot l’axó. No han estat acabades de formar, per tant hi ha una part de l’axó que no està coberta per mielina.
1.3 ORGÀNULS I PARTÍCULES CITOPLASMÀTIQUES.
La neurona, com totes les cèl·lules, conté diversos orgànuls que li permeten duur a terme les seves funcions (nucli, mitocondris, reticle endoplasmàtic, filaments de proteïnes, aparella de Golgi, ribosomes).
Les neurones fan servir una gran quantitat de proteïnes, ja que d’elles depèn el seu funcionament.
1.4.2 CLASSIFICACIÓ SEGONS LA FUNCIÓ.
- NEURONES SENSORIALS. Transmeten la informació rebuda pels receptors de l’exterior al SNC. Moltes són neurones en T.
- INTERNEURONES. Reben informació d’una neurona i l’envien a altres neurones.
- Locals (multipolars Golgi II). Circuits en que la informació es processa a la mateixa zona.
- De projecció (multipolars Golgi I). Transmeten la informació a altres regions del SN. Contenen axons molt mielinitzats.
MOTONEURONES. Connecten el SNC amb els sistemes efectors (músculs, glàndules,…). Són neurons multipolars Golgi I.
AferènciaVa cap a …
(aferent al SNCva cap al SNC)
Eferència Surt
(eferent al SNC surt del SNC)
2. LA NEURÒGLIA DEL SNC.
Les cèl·lules glials mantenen les neurones en la seva posició, per tant tenen una funció estructural. També són vitals pel funcionament i la supervivència de les neurones.
Les cèl·lules glials del SNP són les cèl·lules de Schwann.
Hi ha 3 tipus de glia.
2.1 ASTRÒGLIA.
Conjunt d’astròcits. Són molt abundants i donen suport físic.
Envolten la sinapsis i permeten una certa direccionalitat entre les neurones. Allibera i regula substàncies neurotransmissores.
Intervé en la homeostasi iónica Manté les concentracions iòniques a l’exterior de la cèl·lula (Reguladors).
Regula l’intercanvi de substàncies entre la sang i el teixit nerviós (oxigen i glucosa). Formen la barrera hematoencefàlica, que controla el pas de substàncies de la sang al SN.
Intervé en reaccions immunitàries (glia reactiva). Forma les cicatrius glials.
2.2 OLIGODENDRÒGLIA.
Formen la beina de mielina en el SNC. Els oligodendròcits estan relacionats amb l’esclerosi múltiple, que degenera la mielina de les neurones del SNC. Es tracta d’una malaltia autoimmunitària, les cèl·lules immunitàries deixen de reconèixer com a pròpies les proteïnes que formen part de la mielina i les ataquen, amb la qual cosa provoquen la degeneració d’aquesta, per tant, no es pot transmetre correctament els potencials d’acció. Els oligodendròcits també estan implicats en el manteniment neuronal.
2.3 MICRÒGLIA.
Les cèl·lules de micròglia tenen una forma molt variable. Són les cèl·lules de sistema immunitari que es troben al SNC. Aquestes cèl·lules es movilitzen a la zona afectada quan hi ha una infecció. Actuen eliminant les restes cel·lulars o a l’agent que ha produit la infecció, és a dir, actúen com a macròfags (fagocitosi).
TEMA 2. EXCITABILITAT I CONDUCTIVITAT
NEURONAL.
1.TRANSPORT A TRAVÉS DE LA MEMBRANA
El líquid extracel·lular i intracel·lular conté ions.
Amb més de dos ions permeables, el potencial de membrana será el valor intermig entre Ex+^ i Ez -^.
Dependrà de la diferencia de concentracions entre els dos costats de la membrana.
- Dependrà de la permeabilitat relativa dels diferents ions.
1.3 TIPUS DE TRANSPORT.
TRANSPORT PASSIU
- Difusió simple. Es travessa la membrana a favor del gradient.
- Difusió facilitada.
- Difusió mediada per canal. Proteïna que conté un canal aquós per on passen les partícules.
Difusió mediada per proteïna transportadora. Té dos configuracions, amb les quals transporta les substàncies del medi intracel·lular a l’extracel·lular i viceversa.
TRANSPORT ACTIU
- Proteïnes de transport que traslladen els ions en contra del gradient.
2. POTENCIAL DE REPÒS.
Hi ha un potencial de membrana en totes les cèl·lules del nostre organisme. Aquest és relativament constant. L’interior de la cèl·lula és negatiu respecte a l’exterior.
En les neurones el potencial pot variar en presència de determinats estímuls. Quan el potencial de membrana no rep estímuls s’anomena potencial de repòs (-70mV).
- Depèn de les concentracions iòniques:
- Proteïnes amb càrrega elèctrica a l’interior de la membrana.
- Potassi. Més concentrat a l’interior.
- Sodi. Més concentrar a l’exterior.
- Clor. Més concentrat a l’exterior.
- (^) Depèn de la permeabilitat de la membrana. Els canals iònics controlen el pas d’ions a través de la membrana. Són selectius i van a favor del gradient electroquímic. - Canals passius. Estan sempre oberts. - Canals actius. S’obren només davant de determinats estímuls ( per això varia tant el potencial de membrana).
En repòs només s’obren els canals passius per tant no hi ha canals per als anions, però si molts canals per al potassi.
El potencial en repòs es genera perquè hi ha ions que no poden travessar la membrana. Els anions intracel·lulars proteics obliguen als altres ions a distribuir-se asimètricament ja que no poden travessar la membrana. El principal determinant del valor d’aquest potencial és el potassi, encara que també contribueixen en el seu valor el clor i el sodi.
3. POTENCIAL D’ACCIÓ.
Durant un potencial d’acció (canvi intens en el potencial de membrana) es produeix una entrada massiva de sodi i sortida de potassi. Malgrat això , les seves respectives concentracions es mantenen constants gràcies a un mecanisme de transport actiu anomenat bomba de sodi-potassi. Aquest mecanisme extreu tres ions positius (sodi) i introdueix 2 (potassi), per tant contribueix al valor negatiu del potencial en repòs (-70mV).
3.1 CANVIS EN EL POTENCIAL DE MEMBRANA.
COM ES PRODUEIX EL POTENCIAL D’ACCIÓ?
La membrana varia la seva permeabilitat mitjançant canals passius i canals actius controlats per voltatge, que s’obren quan la membrana arriba a una certa despolarització. Quan la intensitat de l’estímul és prou gran, aquests s’obren i varien les concentracions.
La permeabilitat relativa del sodi augmenta ja que els canals iònics que el regulen són nombrosos i s’obren ràpidament, per tant, el potencial de membrana en aquest moment s’assembla al del sodi (DESPOLARITZACIÓ). Aquests canals es tanquen automàticament.
Quan els canals del sodi es tanquen coincideix en que els canals iònics que regulen el potassi s’obren, per tant, el potencial de membrana en aquest moment s’assemblarà al potencial del potassi. Com que la permeabilitat del potassi és més gran i els canals iònics que el regulen es tanquen més lentament, el potencial es va estabilitzant fins arribar al del repòs (HIPERPOLARITZACIÓ i REPOLARITZACIÓ).
PERQUÈ ELS CANVIS EN LA PERMEABILITAT IMPLIQUEN CANVIS EN EL VALOR DE POTENCIAL DE MEMBRANA?
Si la membrana és permeable a un ió determinat, aquest la travessarà intentant arribar al seu potencial d’equilibri.
Com que els ions de sodi entren al medi intracel·lular, aquest es torna positiu. Quan els canals iònics que el regulen es tanquen, s’obren els de potassi i, com que el seu potencial d’equilibri és de -110mV, voldrà fer que el medi interior sigui negatiu, per tant sortirà a l’exterior ( com que els ions de potassi són positius, anant cap al medi extracel·lular contribuirà a la negativitat de l’interior de la cèl·lula).
Com que les concentracions de sodi i potassi han quedat alterades, s’activa la bomba de sodi-potassi, encarregada de regular aquestes concentracions.
3.4 PERÍODE REFRACTARI.
Si acostem massa dos estímuls llindars en el temps només es produirà un potencial d’acció.
Període refractari general: Temps en el qual un estímul llindar no genera un potencial d’acció. Aquest temps es redueix a mesura que augmentem la intensitat de l’estímul (estímul suprallindar), però té un límit.
Període refractari absolut. Cap estímul pot generar un potencial d’acció. Els canals actius del sodi no estan oberts.
- (^) Període refractari relatiu. Un estímul llindar no provoca un PA, pero un estímul de prou intensitat podrà provocar l’obertura dels canals de sodi.
3.5 CONDUCCIÓ DEL POTENCIAL D’ACCIÓ.
FIBRES AMIELÍNIQUES
El potencial d’acció es produeix a cada punt de la membrana (la zona on comença l’estímul sol ser el segment inicial axònic, i es condueix en sentit ortodròmic, és a dir, del soma als botons axònics). Conducció contínua. Aquest PA serveix d’estímul per la zona veïna de la membrana.
FIBRES MIELÍNIQUES
funcionar en les dues direccions. Són extremadament rígides.
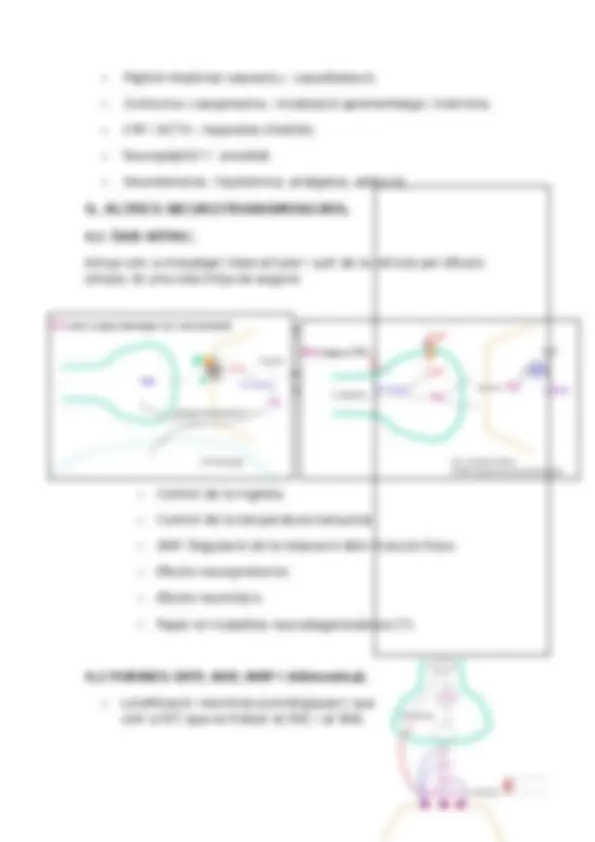
- Sinapsis químiques. L’espai entre les neurones és gran. La neurona presinàptica allibera un missatger químic (neurotransmissor) que interacciona amb el receptor de la neurona postsinàptica, això produeix un canvi en la permeabilitat de la membrana postsinàptica i allibera un potencial d’acció.
- Segons els efectes postsinàptics:
- (^) Sinapsis excitadores. La resposta postsinàptica és una despolarització ( més possibilitats de potencial d’acció en la neurona postsinàptica).
- Sinapsis inhibidores. La resposta postsinàptica és una hiperpolarització (menys possibilitats de potencial d’acció en la neurona postsinàptica).
- Segons el lloc de contacte:
- Sinapsis axo – dendrítiques. El botó terminal acaba sobre una dendrita. Sovint són excitadores.
- Sinapsis axo – somàtiques. El botó terminal fa sinapsi sobre el cos cel·lular. Sovint són inhibidores.
- Sinapsis axo – axòniques. El botó terminal termina sobre un axó. Serveixen per regular la potència del senyal elèctric.
- Sinapsis de pas. Al llarg de la ramificació axònica hi ha varicositats que funcionen com a botons terminals que afecten a les neurones del voltant. S’alliberen neurotransmissors que es difonen pel medi i actuen sobre grans regions alhora, generant sinapsis en les neurones properes.
1. ALLIBERAMENT DEL NEUROTRANSMISSOR.
Els neurotransmissors es troben en la vesícula sinàptica. Quan es produeix la despolarització, els canals controlats per voltatge s’obren i el calci entra a l’interior cel·lular a favor del gradient electroquímic.
Quan augmenta la quantitat de calci intracel·lular les vesícules sinàptiques s’acosten a la membrana cel·lular i es fusionen amb aquesta, i allò que hi ha a l’interior de la vesícula és alliberat a l’exterior cel·lular (exocitosi). L’exocitosi només es dona en llocs determinats de la membrana on hi ha proteïnes específiques (zones actives).
Una vegada utilitzades, les vesícules es desencaixen de la membrana per tornar a ser utilitzades (reciclatge de vesícules).
2.RECEPTORS POSTSINÀPTICS.
Els neurotransmissors es difonen en el medi i arriben als receptors postinàptics per una interacció clau – pany. Aquestes proteïnes (receptors), quan se’ls uneixen els neurotransmissors activen l’obertura dels canals iònics de la membrana postsinàptica, el canvi de permeabilitat de la qual provoca un potencial d’acció. Aquests canals que s’obren pel fet de que una molècula s’uneixi a una proteïna s’anomenen canals controlats per lligand.
TIPUS DE RECEPTORS
Ionotròpics: complex receptor – canal. Proteïnes amb forma de canal que quan se’ls uneix el neurotransmissor s’obren.
Metabotròpics: el receptor i el canal són proteïnes diferents. Un cop s’ha unit el neurotransmissor al receptor, aquest activa la proteïna G, la qual té una subunitat que quan s’activa s’uneix al canal iònic i l’obre.
En un altre cas, el neurotransmissor s’uneix, el receptor activa la proteïna G, la subunitat d’aquesta activa un enzim al·lostèric el producte del qual és una molècula amb capacitat per moure’s pel citoplasma (2n missatger). Aquest 2n missatger pot actuar sobre les proteïnes quinases que trenquen l’ATP en ADP + Pi, aquest Pi s’uneix
Els efectes postsinàptics depenen del canal que s’obri, no del neurotransmissor. Un mateix neurotransmissor pot tenir efectes postsinàptics diferents si actua sobre receptors diferents.
4.MECANISMES D’INTEGRACIÓ SINÀPTICA.
El segment inicial axònic està realitzant en cada moment una sumació algebraica de PEPs i PIPs. Si el resultat arriba al llindar de descàrrega es produeix un potencial d’acció.
Sumació temporal. Es sumen els efectes dels potencials que arriben als diferents punts de la membrana en un temps reduït.
Sumació espacial. Es sumen els efectes dels potencials en un mateix moment.
5.MECANISMES D’INACTIVACIÓ DEL
NEUROTRANSMISSOR.
Procés pel qual impedim que el neurotransmissor s’uneixi al receptor.
- Degradació enzimàtica. Mitjançant un enzim alterem l’estructura del neurotransmissor amb l’unió d’una molècula. Aquest no es pot unir al receptor.
- (^) Recaptació. Hi ha un mecanisme de transport actiu d’alta afinitat pel neurotransmissor que el capta i el transporta cap a l’interior de la cèl·lula presinàptica. No és un receptor, és un transportador. Si aquest procés té lloc des d’una neurona
s’anomena recaptació neuronal, si en té lloc des d’una cèl·lula glia s’anomena recaptació glial.
Cada neurotransmissor té un mecanisme d’inactivació diferent.
6. MODULACIÓ SINÀPTICA.
La modulació sinàptica es duu a terme mitjançant els receptors presinàptics.
- Autoreceptor. Reconeix i capta neurotransmissors procedents de la pròpia cèl·lula en la que es troba. Controlen la síntesi i l’alliberament de neurotransmissors, sovint de forma inhibitòria. Si n’hi ha molts en el medi, els capten.
- Heteroreceptor. Reconeix i capta neurotransmissors procedents d’altres cèl·lules. Receptor per terceres neurones (sinapsi axo-axònica), és a dir, aquelles neurones que no intervenen en la sinapsi bàsica entre les dos neurones.
INHIBICIÓ PRESINÀPTICA.
Quan l’heteroreceptor rep neurotransmissors de la tercera cèl·lula, aquest s’activa i modula els canals de calci, produint una disminució de calci a la cèl·lula, per tant, s’alliberaran menys neurotransmissors (sinapsi axo-axònica inhibitòria).
FACILITACIÓ PRESINÀPTICA.
Quan l’heteroreceptor rep neurotransmissors de la tercera cèl·lula, aquest activa i modula l’entrada de calci, produint un augment de la quantitat de calci a la cèl·lula, alliberant així més neurotransmissors (sinapsi axo – axònica facilitadora).
7.MECANISMES DE PLASTICITAT SINÀPTICA.
Plasticitat sinàptica: canvis bioquímics i/o estructurals en les connexions entre les neurones que proporcionen a l’organisme la capacitat d’adaptació a l’entorn. Hi ha dos tipus de canvis:
- Potenciem l’inactivació de NT: - alteració de l’efecte postsinàptic.
- Potenciem els autoreceptors: - alteració de l’efecte postsinàptic.
- Si inhibim la inactivació: major efecte postsinàptic.
3. RECEPTORS.
Hi ha dos tipus de receptors:
- AGONISTA: molècules amb el mateix efecte que el NT. Lligands del receptor que reprodueixen l’acció del NT.
- ANTAGONISTA: lligands que en unir-se al receptor bloquegen l’acció del NT. - Antagonista competitiu (directe). Com una clau que no és la que obre el pany però entra fent que el NT no es pugui unir, és a dir, s’uneix al mateix lloc que el NT. - Antagonista no competitiu (indirecte). S’uneixen a un lloc diferent que el NT, però bloqueja el seu efecte. - Antagonista invers. Substància que quan s’uneix produeix l’efecte contrari que el NT.
4. AMINOÀCIDS EXCITADORS.
- Localització: SNC (principals components de les sinapsis excitadores).
- Síntesi: glutamina (mitjançant la glutaminasa, en el botó terminal)glutamat (mitjançant l’aspartat aminotransferasa) aspartat.
Segons quin sigui el NT, la sinapsi serà glutamatèrgica o aspartatèrgica.
- Inactivació i reciclatge: recaptació.