



ATENCIÓN
Este documento está elaborado en
base a las clases dadas por Yolanda
León en 2015.
Se recomienda que además se consulte
otras fuentes para contrastar.





















Prepara tus exámenes y mejora tus resultados gracias a la gran cantidad de recursos disponibles en Docsity

Gana puntos ayudando a otros estudiantes o consíguelos activando un Plan Premium

Prepara tus exámenes
Prepara tus exámenes y mejora tus resultados gracias a la gran cantidad de recursos disponibles en Docsity
Prepara tus exámenes con los documentos que comparten otros estudiantes como tú en Docsity
Encuentra los documentos específicos para los exámenes de tu universidad
Estudia con lecciones y exámenes resueltos basados en los programas académicos de las mejores universidades
Responde a preguntas de exámenes reales y pon a prueba tu preparación
Consigue puntos base para descargar
Gana puntos ayudando a otros estudiantes o consíguelos activando un Plan Premium
Comunidad
Pide ayuda a la comunidad y resuelve tus dudas de estudio
Ebooks gratuitos
Descarga nuestras guías gratuitas sobre técnicas de estudio, métodos para controlar la ansiedad y consejos para la tesis preparadas por los tutores de Docsity
Asignatura: Fisiología Animal, Profesor: Yolanda León Álvarez, Carrera: Biología, Universidad: UAM
Tipo: Apuntes
1 / 32

Esta página no es visible en la vista previa
¡No te pierdas las partes importantes!
El medio aéreo presenta ventajas con respecto al acuático:
La concentración elevada y constante de oxígeno: o Permite unas tasas metabólicas elevadas. Evolutivamente, los animales con tasas metabólicas elevadas aparecen entre los que ventilan en el aire. o Respiración bidireccional, en general. Las velocidades de difusión que pueden alcanzar los gases (O2 y CO2) son muchísimo más rápidas que en el agua (La difusión de oxígeno a la misma presión parcial es superior a 2000. veces ene l aire que en el agua), lo que trae como consecuencia que se puedan alcanzar mayores distancias de difusión.
Pero también una desventaja: una gran evaporación de agua , por lo que las superficies de intercambio respiratorio se localizan en cavidades respiratorias especializadas (Tráqueas y Pulmones), por tanto, el acceso al aire está limitado.
Los insectos presentan un cuerpo recubierto por una serie de poros o invaginaciones ( espiráculos ), que comunican con un sistema de tubos ramificados ( tráqueas ) que van disminuyendo su diámetros a medida que se ramifican dando lugar a las traqueolas , que comunican directamente con las células para suministralas oxígeno en función de su demanda (luego el aparato circulatorio desempeña un papel mínimo o nulo en el transporte de este gas).
Los espiráculos están ubicados en la superficie corporal a lo largo de la pared corporal lateral. Son unos poros especializados, rodeados por una cutícula y con una serie de pelos filtradores en el exterior cuyas funciones fundamentales son:
Su número, así como su apertura, varía en los animales, pues podemos encontrar espiráculos con una ventilación unidereccional (el mismo espiráculo se abre para introducir O 2 y expulsar CO 2 ) o bidireccional (unos se abren para introducir oxígeno y otros para liberar dióxido de carbono).
Las tráqueas son unas invaginaciones que ingresan en el cuerpo a través del espiráculo, están forradas de cutícula (epicutícula) y presentan unos repliegues en forma de acordeón cuya función es la expansión y compresión de esta red de tubos que funciona a modo de fuelle (movido por los propios movimientos musculares) ventilando el sistema y evitando el colapso.
A medida que aumenta la distancia entre los espiráculos por la ramificación de las tráqueas, éstas reducen su diámetro, luego las tráqueas mayores poseen una cutícula de un grosor similar al del cuerpo del animal, mientras que las menores tienen cutículas más finas, hasta que originan túbulos terminales
insectos, no en todos. Aparece principalmente en animales que presentan un metabolismo más alto y en animales que son más grandes.
Por tanto, los principales factores influyentes en la difusión y ventilación en insectos son:
Cascada de Oxígeno: Requerimiento mitocondrial
El oxígeno incluido del exterior, sufre una caída de su presión parcial hasta que llega a las mitocondrias, en las cuales entrará por difusión a una tasa que depende del ΔPp entre los capilares sistémicos y la mitocondria. Esta reducción de la presión parcial en los extremos de las tráqueas aumenta la diferencia entre las presiones parciales en los dos extremos del sistema traqueal y de esta manera acelera la difusión del O 2 a las traqueolas (los espiráculos están cerrados, por lo que la velocidad de transporte de oxígeno es más rápido). La presión parcial en los extremos de las tráqueas debe permanecer bastante elevada para que el O 2 se difunda desde estos extremos hacia las mitocondrias intracelulares, es decir, debe asegurar siempre una mínima presión (1 mm Hg) para mantener el catabolismo aerobio. En consecuencia, nos enfrentamos a la cascada de oxígeno entre el aire ambiental y las mitocondrias.
Por tanto, la tasa de difusión en insectos depende de dos fuerzas motrices:
Los insectos modulan la apertura y cierre de los espiráculos en función de sus periodos de actividad/reposo. Cuando un insecto está en reposo, en determinadas situaciones, es capaz de cerrar sus espiráculos y no no se restringe la tasa de influjo de oxígeno, pues pasa a depender del oxígeno almacenado en su tronco traqueolar, ya que no interesa introducir oxigeno que no se destine a ATP, pues puede formar radicales libres. La constricción de los espiráculos dificulta la difusión del O 2 hacia el sistema traqueal y de esta manera tiende a disminuir la presión parcial de O 2 en los extremos internos de las tráqueas. Sin embargo, este efecto no impide el desarrollo de una presión parcial de O 2 en la parte interna de las tráqueas lo bastante elevada para que este gas se difunda hacia las mitocondrias a una velocidad adecuada para cubrir las necesidades de O 2 en reposo.
Cuando los insectos inician su actividad y requieren mayor cantidad de O 2 pueden requerir la apertura de sus espiráculos para mantener una presión parcial de O 2 elevada en la parte interna de la tráquea. Por tanto, los insectos en actividad aumentan la apertura de sus espiráculos, y pagan el precio del incremento de la evaporación. Por tanto, que el espiráculo esté cerrado impide la pérdida de agua.
Al aumentar el gradiente de presiones (Fick), se produce el aumento de la presión parcial de CO 2 y la disminución del pH de los fluidos corporales , éstos son los dos estímulos necesarios para que se produzca la apertura de los espiráculos. El CO 2 en forma de bicarbonato actúa como tampón, por tanto, si disminuye el pH aumentan los protones, por lo que el bicarbonato recoge los protones sobrantes para intentar mantener el pH corporal, dando lugar a CO2. De manera que, tanto el CO 2 como el aumento de protones estimulan la apertura de los espiráculos.
Este tipo de respiración se da en insectos que se encuentran en momentos de baja actividad ; quiescencia o diapausa y viven en un medio con una concentración de oxígeno relativamente alta, aunque sus células no precisen de tanto O 2 , lo cual puede ser peligroso para el animal. En este tipo de respiración, la apertura de los espiráculos dura menos tiempo y es diferente.
El control de los espiráculos en periodo de actividad permite mantener una determinada cantidad de O 2. En periodos de inactividad la apertura de los espiráculos tiene lugar durante menos tiempo y de forma diferente; poseen una fase abierta , una cerrada y otra en fase de aleteo (Flutening: impide la caída total de la presión parcial de oxígeno). En cualquiera de estas tres fases los espiráculos mantienen un consumo de O 2 constante , sin embargo, la expulsión de CO 2 no es constante , solo se expulsa con el espiráculo abierto. La caída de CO 2 no es tan brusca como la de O 2 ya que se encuentra en disolución
entra se consume rápidamente, siendo el N 2 el responsable de equilibrar esto.
Una característica llamativa de la burbuja de gas es que actúa como una branquia y no solo como depósito de O 2. Cuando un insecto que nada debajo del agua elimina O 2 proveniente de su burbuja, la presión parcial de O 2 en la burbuja disminuye. Cuando esta presión disminuye por debajo de la presión de O 2 del agua, el O 2 disuelto en el agua comienza a difundirse hacia la burbuja y renueva el oxígeno.
La velocidad de difusión del O 2 dentro de la burbuja depende de su superficie. Las burbujas descritas disminuyen de tamaño de forma gradual y su superficie se reduce, lo que determina el cese de su función efectiva como branquia.
3-En algunos insectos acuáticos ciertas partes de la superficie corporal están cubiertas por una capa muy densa de pelos hidrófobos delgados impermeables al agua. Estos pelos atrapan una película de aire casi invisible. Esta película de aire, se denomina plastrón , es incomprensible y permanente. Su superficie es constante y puede servir como indefinidamente como branquia gaseosa. Muchos insectos con este tipo de mecanismo pueden permanecer meses sumergidos en aguas bien aireadas. ( Ver Práctica en Aula 2 del Segundo Semestre )
Los pulmones son estructuras invaginadas con acceso al oxígeno del aire, forman una especie de sacos internos y complejos en la cavidad corporal, el hecho de que sean unas invaginaciones internas es para hacer frente al problema de la evaporación de agua.
En el mundo animal hay una gran variedad, desde los más sencillos de los anfibios, pasando por los relativamente complejos de los reptiles hasta terminar con los más complejos de los mamíferos y
aves. En la escala evolutiva, cada vez los sacos se van haciendo más grandes, con más ramificaciones, repliegues e invaginaciones, aumentando así la superficie de intercambio gaseoso, de manera que para el mismo volumen pulmonar todas esas invaginaciones lo que dan lugar es a que haya mayor superficie de intercambio asociada a su vez a mayores tasas metabólicas ( Rana : 1 ml ⇨ 20 cm2 de superficie de intercambio gaseoso; Ratón : 1 ml ⇨ 800 cm2 de superficie de intercambio
Equilibrio de Pp
gaseoso), además, para el caso de los mamíferos, es más patente la relación directa del tamaño corporal y el volumen pulmonar (a mayor tamaño, mayor volumen pulmonar).
Los anfibios lograron la mejor combinación entre la respiración en el agua y la respiración en el aire. Muchos de estos animales se mueven desde un ambiente acuático hacia uno terrestre durante su desarrollo y muchos respiran en ambos ambientes durante la adultez. Por tanto, además de pulmones, presentan otras superficies respiratorias como la piel o las branquias.
En todas las larvas de los anfibios jóvenes las branquias son externas. Las branquias suelen perderse durante la metamorfosis.
En la mayoría de los anfibios se desarrolla un par de pulmones a partir de la invaginación de la pared ventral de la faringe. Son sacos simples muy vascularizados. La superficie interna de los pulmones aumenta poco o nada debido a la presencia de algunos pliegues que están menos desarrollados que en los pulmones de los peces pulmonados modernos.
Para la ventilación del animal es fundamental la glotis (que favorece los periodos de apnea en la respiración discontinua), la cual se sitúa al final de la laringe en contacto con los pulmones. La cavidad central de cada pulmón permanece abierta y permite acceder a los diversos compartimentos laterales formados por los pliegues en las paredes.
Los anfibios pulmonados ventilan sus pulmones por compresión orofaríngea ; es un mecanismo de presión positiva , es decir, que el aire el introducido de forma forzada en los pulmones (algo parecido al de los peces). La bomba bucal es la encargada de implsar el aire hacia el interior de los pulmones gracias a la presión bucofaríngea que se ejerce en el suelo de la boca:
1- El aire fresco ingresa en la cavidad bucal a través de las fosas nasales o de la boca cuando la presión en la cavidad bucal disminuye y debido al descenso del suelo de la boca, manteniendo la glotis cerrada, y se acumula en el espacio que se ha formado. 2- Luego se eleva el piso de la cavidad bucal con la boca cerrada y las narinas selladas por lo menos en forma parcial gracias a la presencia de válvulas y el aumento de la presión obliga al aire a ingresar en los pulmones. La insuflación de los pulmones eleva la presión del aire dentro de ellos. Esto significa que si no fuera por la glotis los pulmones eliminarían el aire cuando se abre la boca o las fosas nasales, que se cierra después de la inhalación por medio de la contracción de los músculos. 3- Luego se produce un periodo de ausencia de respiración con los pulmones insuflados, es decir, la respiración es intermitente. Las fosas nasales se abren durante el periodo de apnea y en ese momento la rana suele bombear aire dentro y fuera de la cavidad bucal a través de las fosas nasales por medio del descenso y la elevación del piso de la cavidad bucal, proceso denominado bombeo bucofaríngeo.
La captación de O 2 por la piel es prácticamente constante, pero en los meses más cálidos donde aumenta la temperatura y la tasa metabólica , los pulmones toman un papel más relevante.
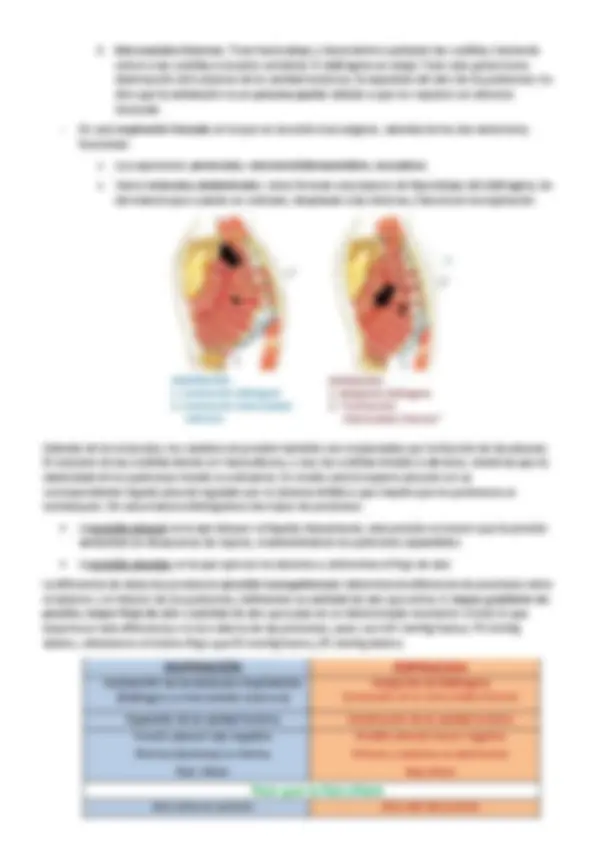
En la mayoría de los reptiles los pulmones absorben casi todo el O 2 y eliminan casi todo el CO 2. Su piel suele ser menos permeable que la de los anfibios, lo que significa que protege mucho más contra la deshidratación por evaporación, pero que no permite el pasaje de los gases respiratorios de forma sencilla.
Hay dos tipos de pulmones:
gases entre la cavidad central y las profundidades de las celdillas en las paredes se realice por difusión. Este tipo de tabicación aumenta la superficie respiratoria en animales menos activos.
Los pulmones se expanden y, por tanto, tienen menos presión que el exterior. Presentan respiración discontinua , para lo que es necesario la existencia de la glotis, además, las costillas tienen un papel muy importante en los cambios de volumen de la cavidad. Estas diferencias entre las cavidades se generan por la acción de los músculos intercostales (lagartos y serpientes) y diafragmiticus (cocodrilos). En lagartos y serpientes, para que se produzca la entrada de aire se contraen los músculos intercostales (hacia atrás y hacia abajo) y el movimiento de las costillas (hacia abajo y hacia afuera), aumentando el tamaño de la cavidad torácica. Esta expansión genera el estiramiento pulmonar y una presión negativa, que permite la entrada de aire siempre y cuando la glotis permanezca abierta, fenómeno conocido como inspiración. En cocodrilos hay un precursor del diafragma que es lo que denomina mos diafragmitus o músculo diafragmático, el cual tira del hígado hacia la pelvis, provocando un aumento de la cavidad, y por tanto, el llenado pulmonar. Evidencias actuales sugieren que el flujo del aire en los pulmones de los cocodrilos y algunos lagartos es unidireccional , pues hay una serie de comunicaciones entre las cámaras que definen el pulmón. Estos animales cierran la glotis y el aire inhalado queda en los pulmones durante varios segundos o minutos de apnea , mientras la cavidad bucal suele ventilarse a través de una bomba bucofaríngea. Esta fase de apnea dura más o menos en función de la tasa metabólica, lo que les permite estar mucho tiempo sin respirar, sin embargo, los niveles de CO 2 aumentan. La expiración ocurre invirtiendo la presión. La relajación de estos músculos da lugar a un retroceso elástico pasivo de los pulmones, y por tanto, se genera un aumento de presión en el interior de los pulmones, lo que produce la expulsión del aire.
Contracción de los músculos → Cavidad más grande (Volumen mayor) Aumento de presión Succión/inspiración (entrada del aire)
Relajación de los músculos→ Cavidad más pequeña (Volumen menor) Aumento de presión Espiración (se expulsa el aire)
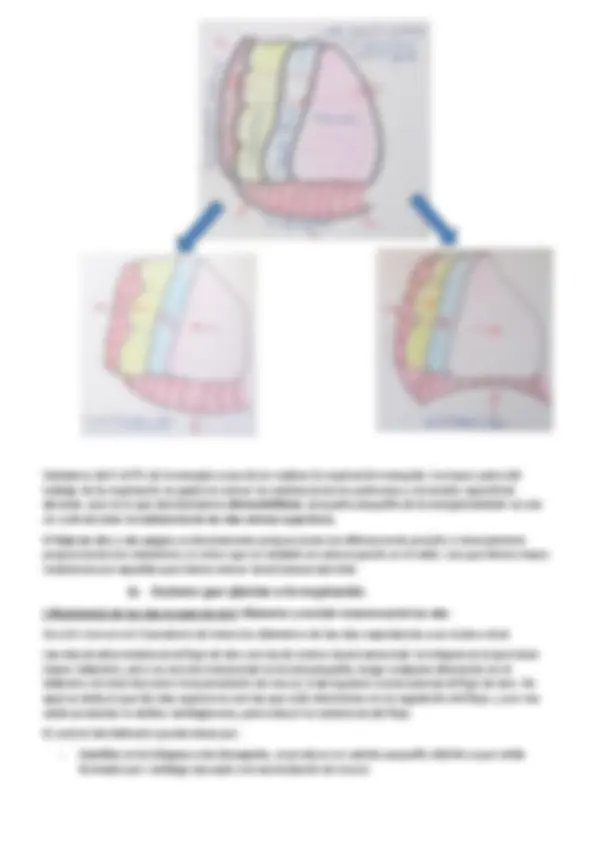
El pulmón (paleopulmón) es muy rígido, tanto que no cambia de volumen, sin embargo si se produce una succión de aire, pero es creada por los sacos aéreos, ejerciendo un presión negativa en el cuerpo de estos animales.
Los sacos aéreos forman parte del aparato respiratorio, se localizan fuera de los pulmones y ocupan una porción considerable de las cavidades corporales torácica y abdominal (todas las zonas del cuerpo que carezcan de vísceras), incluso algunos penetran en los huesos. Son estructuras de paredes delgadas y vascularización escasa que son esenciales en la ventilación , pero no participan en el intercambio gaseoso, simplemente permiten acumular aire sin aumentar mucho el peso del animal. Por lo general hay 9 sacos aéreos que se dividen en dos grupos:
Los sacos aéreos posteriores tienen menor proporción de CO 2 y más concentración de O 2 , mientras que en los sacos anteriores hay una mayor proporción de CO 2 que de O 2. Esto llevó a la determinación de que en la ventilación el flujo de aire es unidireccional y siempre postero-anterior (de atrás hacia delante).
Como ya hemos visto en el dibujo, las aves poseen un pulmón principal denominado paleopulmón, compuesto por el mesobronquio (bronquio primario), bronquio medioventral y mediodorsal (bronquios secundarios) y los parabronquios (bronquios terciarios). Además, la mayoría de las aves tienen un sistema de tubos parabranquiales más o menos desarrollados que transcurren entre los sacos aéreos posteriores y las partes posteriores de los mesobronquios y los dorsonronquios. Este sistema de tubos se denomina neopulmón (20-25% del volumen pulmonar total).
Recordemos que la tráquea, los bronquios, el neopulmón y los sacos aéreos, son estructuras en las que NO se produce intercambio gaseoso
Otra característica de las aves a tener en cuenta, es que como la mayoría de los reptiles no presentan diafragma
Bronquios 2º
Las vías aéreas superiores suponen un espacio fisiológico muerto, sin embargo, son necesarias para la conducción de oxígeno. La parte superior de estas vías calienta y humedece el aire así como elimina los patógenos y partículas extrañas que se hayan podido incorporar mediante la fabricación de un moco (el cual se eliminará posteriormente por el tracto digestivo o se exala por un mecanismo parecido a la tos) y gracias a la presencia de unos cilios dotados de inmunoglobulinas. Es destacable que en estas vías habrá tanto oxígeno como dióxido de carbono, pues el aire tiene O 2 , CO 2 y 78% de otros gases que no se incorporan, mientras que el oxígeno se absorbe, el dióxido de carbono se mantiene, y cuando se produce la expiración, en la tráquea se encontrará el CO 2 inspirado y el creado por la respiración, luego se reduce la presión parcial de oxígeno.
Los pulmones de las aves son estructuras compactas y rígidas, se diferencian de los pulmones de los mamíferos porque sufren cambios mínimos en el volumen durante el ciclo respiratorio. En cambio, los sacos aéreos se expanden y contraen en forma notable durante la ventilación. Al igual que fuelles, los sacos aéreos succionan y expulsas gases a través de las vías aéreas rígidas de los pulmones.
¿Cómo se hinchan y deshinchan los sacos aéreos?
Durante la inhalación la contracción de los músculos intercostales internos y de otros músculos torácicos (en concreto el apendicocotalis ) expande otras partes de la caja torácica (cuyas costillas están articuladas y unidas al esternón) y el esternón se mueve hacia abajo y delante. Cualquier movimiento que se haga de la cavidad toracoabdominal va a estar sujeto a un cambio de volumen, y en este caso se desencadena el aumento de los sacos aéreos.
Durante la exhalación algunos músculos intercostales externos y abdominales comprimen esta cavidad y los sacos aéreos, y el esternón se eleva.
El ritmo de respiración es controlado por una zona marcapasos del bulbo raquídeo, que manda señales a los musculos de las costillas que se contraen:
Unos musculos abren el esternón: appendicocostalis inspiraciónabren angulación de las costillastiran del esternónhinchan sacos aéreos abriéndolos
anterior, combinado con el gas que se elimina de los sacos aéreos anteriores, se dirige hacia el mesobronquio a través de los bronquios secundarios anteriores y se exhala.
Se produce intercambio tanto en la inspiración como en la espiración; lo que es una gran ventaja, por ejemplo, respecto al propio ser humano. Siempre pasa aire por las superficies respiratorias.
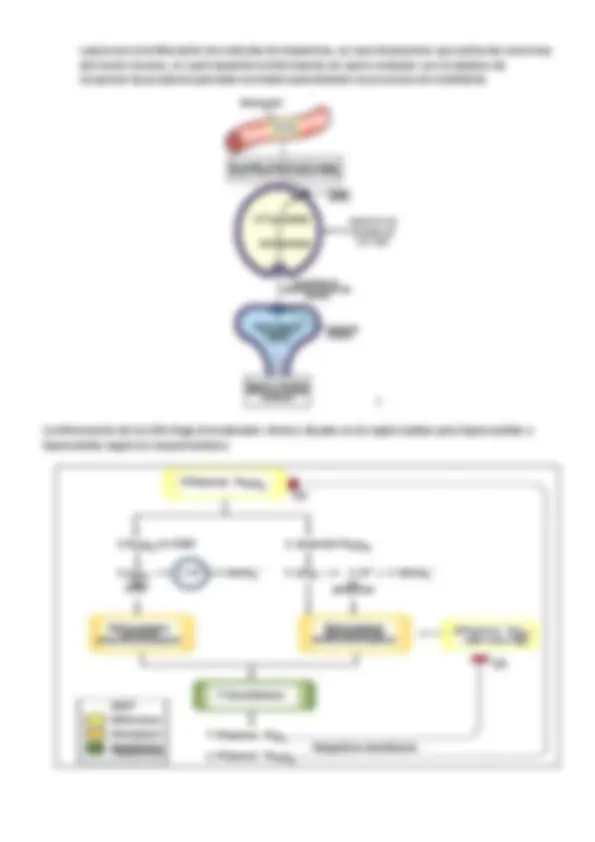
El flujo de sangre en los vasos que desarrollan el intercambio respiratorio revela un patrón de corrientes cruzadas en relación con el flujo de aire que atraviesa los parabronquios.
El aire entra por la luz del vaso aferente del parabronquio ( arteria pulmonar ) y sale por la vía eferente ( vena pulmonar ). Cada capilar establece contacto con una zona del parabronquio, de manera que los capilares en contacto con la zona del principio del parabronquio captarán más O 2 que los más alejados. En la arteria pulmonar, la sangre de salida será una mezcla de todos esos capilares con 0% de oxigeno diferentes.
El mecanismo contracorriente se diferencia de las corrientes cruzadas en que no se va enriqueciendo la sangre. En contracorriente si inviertes el flujo no consigues el enriquecimiento de la sangre, mientras que en el mecanismo de corrientes cruzadas sí; a cualquier nivel del parabronquio siempre va a haber
Esternón Esternón
1. Vaso aferente 2.Dorsobronquio 3.Parabronquio 4.Vaso eferente 5.Ventrobronquio
una presión positiva , porque el vaso con el que establece contacto es un vaso que siempre tiene la menor presión parcial de O 2.
Otra explicación:
El aire sale del saco aéreo posterior y viaja por el dorsobronquio hasta llegar a los parabronquios. En el parabronquio los gases difunden por sus membranas respiratorias hasta los capilares sanguíneos, éstos provienen de un vaso aferente que se ramifica en el parabronquio, como su presión parcial de oxígeno es muy baja (casi cero) se oxigena y comunica con un vaso eferente que se destina a los tejidos. La cantidad de oxígenos que los capilares adquieren, se va reduciendo a medida que se ramifica el
vaso, sin embargo, como su presión parcial de oxígeno es mucho más baja que la de los parabronquios, siempre se van a oxigenar. Con este sistema, el animal tiene un 60-65% de eficiencia a la hora de obtener el oxígeno.
Al ser un modelo de corrientes cruzadas, da igual que la sangre valla en la misma dirección que el aire, ya que lo esencial es que el vaso aferente se ramifique con una presión parcial de oxigeno cercana a 0 y entre en contacto con el aire con una presión aferente mayor
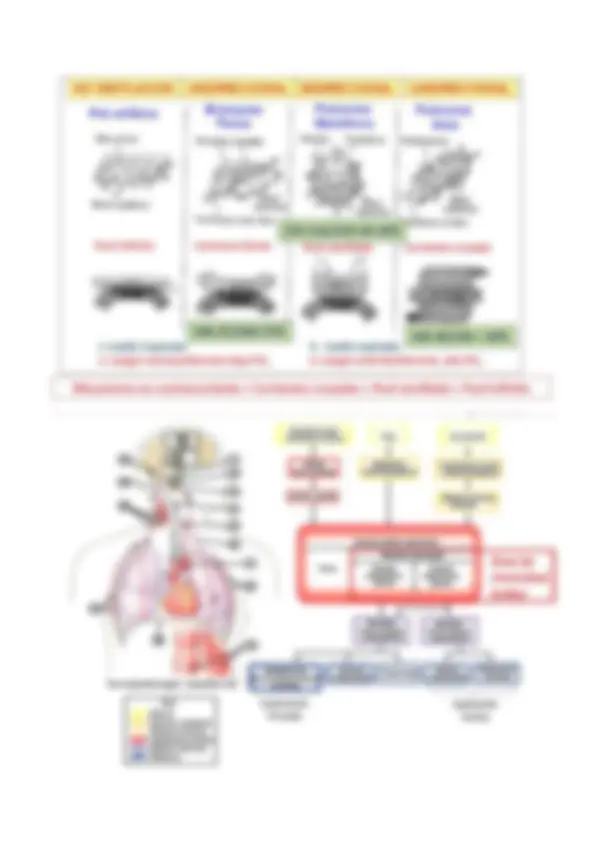
Los mamíferos ya gozan de un verdadero diafragma, cuya contracción es vital para la ventilación del animal, y de esta forma, podemos diferenciar una caja torácica separada de la abdominal.
El sistema pulmonar de un ser humano adulto está compuesto por 23 niveles de ramificaciones en las vías aéreas. El aire entra por las vías nasales (mucho más beneficioso que por la boca), cruza la faringe y atraviesa la laringe hasta que llega a la tráquea , ésta se ramifica para formar una vía aérea principal, el bronquio fuente , que ingresa en cada pulmón. Cada bronquio fuente se ramifica dentro del pulmón para originar
¿Por qué es importante calentar y humedecer el aire? Si nos fijamos, no hay ningún órgano que esté en contacto con el aire, ya que este impide la funcionalidad de las células (es verdad que la piel sí lo está, pero por que cuenta con un tejido estratificado muerto), de esta manera, siempre se necesita una fina capa de agua para mantener los gradientes de las células. Con esto, con la espiración, los mamíferos estarían perdiendo grandes cantidades de agua, por eso los animales disponen de un sistema en contracorriente para humedecer y calentar el aire, optimizando así la pérdida de agua: o INSIRACIÓN: Cuando entra aire seco por la nariz (22ºC y 155 mmHg O 2 ) se calienta gracias a un gradiente de temperatura que le cede calor y agua a medida que avanza por la zona nasal (↑Tº↑PpH2O), de manera que cuando llega a la faringe ya tiene la presión de O final, menor que la inicial debido a su hidratación y su temperatura 37º. o ESPIRACIÓN: Cuando sale el aire en la espiración, al estar en contacto con unos capilares con una temperatura inferior, se enfría y cede el agua que se condensa, de manera que sale más seco. Esto es muy importante debido a que uno de los grandes problemas es la evaporación, por lo que mantener el gradiente nasal es imprescindible para mantener la conservación del agua, de ahí a que sea más eficiente, en cuanto a la conservación del agua, respirar por la nariz en vez de por la boca.
Los alveolos son sacos interconectados entre sí que se agrupan al final de los bronquios. Están formados por una capa de tejido epitelial con dos tipos de células:
Los alveolos se encuentran en íntima asociación con los capilares, de modo que hay una capa casi continua de sangre que cubre éstos, donde tiene lugar el intercambio gaseoso. De manera que la capa de intercambio está formada por:
Capa surfactante. Membrana epitelial del epitelio. Células del alveolo. Capilares de pequeño diámetro. A esta capa de intercambio hay que sumarle la membrana de los eritrocitos, pues es dentro de éstos donde se encuentra la hemoglobina, aun así, estas capas son muy finas, por lo que la distancia de difusión no se ve muy perjudicada.
Cuando los bronquios terminales se quedan sin cartílago a lo largo de su ramificación, disponen de músculo liso, sin embargo, los alveolos carecen de músculo liso (porque la membrana de intercambio tiene que ser lo más fina posible); pero poseen gran cantidad de fibras de elastina y colágeno que le confieren gran elasticidad al pulmón, permitiendo sus movimientos de expansión y compresión.
El cambio de volumen de los pulmones produce cambios en las presiones del medio externo y el interno, dando lugar a la ventilación.
Cuando la presión alveolar es menor que la presión atmosférica el aire entra inspiración. Cuando la presión alveolar el mayor que la atmosférica el aire sale espiración.
En la respiración normal o tranquila, estos cambios son provocados por el juego de los músculos intercostales y el diafragma :