GABA
Gaba como neurotransmisor: más allá de las aminas biógenas
–NT muy distinto de las monoaminas, neuronas gabaérgicas hay en todas las zonas del SNC
(apenas se encuentra en otros tejidos). Forman parte de circuitos locales dentro de cada zona
del SN, son interneuronas inhibidoras (junto con las de glicina en algunos casos).
–Se encuentra en muchos sitios, como bacterias, plantas...
–La mayoría de los estudios del GABA eran electrofisiológicos, es muy complicado
estudiarlo porque está dentro de las rutas intermediarias de las rutas de los carbohidratos y
es difícil diferenciarlo (el pool metabólico es mucho mayor que el de NT). Una de las
razones por las que el GABA adquirió interés fue por el descubrimiento de que los
ansiolíticos clásicos (benzodiacepinas) funcionaban en los receptores del GABA.
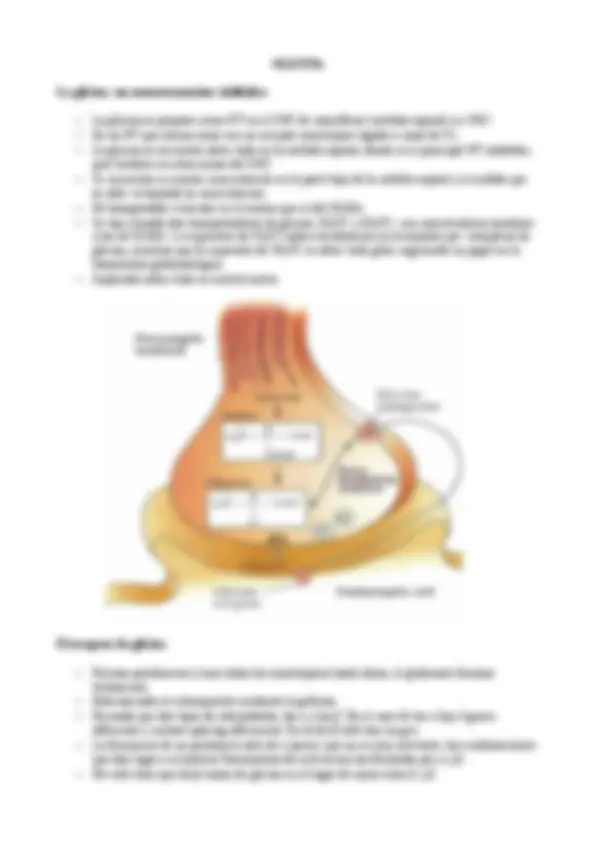
Síntesis de GABA a partir de glutamato
–El GABA se forma fundamentalmente a partir del glutamato por el enzima GAD. El
glutamato y el GABA están relacionados con el ciclo de Krebs por la GABA transaminasa.
–El GAD puede ser el GAD-65 y GAD-67, son genes distintos pero con un 90% de
homología. Cuando se habla de neurotransmisión se considera que el más importante es el
GAD-65 (es más sensible al piridoxal-fosfato). Cuando uno analiza dónde está el GAD
situado en las neuronas, el 65 se encuentra en las terminales sinápticas en mayor
concentración mientras que el 67 se encuentra más repartido, también hay en el soma y
dendritas (también en terminales). Para marcar neuronas GABAérgicas se utiliza el GAD-67
porque es el que se puede encontrar en el soma. También se puede hacer hibridación in situ
contra el GAD-65 porque el mRNA sí que está en el soma.
–Los animales KO para el GAD-65 tienen problemas neurológicos, pero los KO para el
GAD-67 son letales por lo que debe tener un papel que aún se desconoce.
–La fosforilación de la GAD-65 activa la enzima mientras que inhibe la 67.
–La GABA-T es una enzima mitocondrial que también requiere piridoxal-fosfato.
Transportador vesicular de GABA
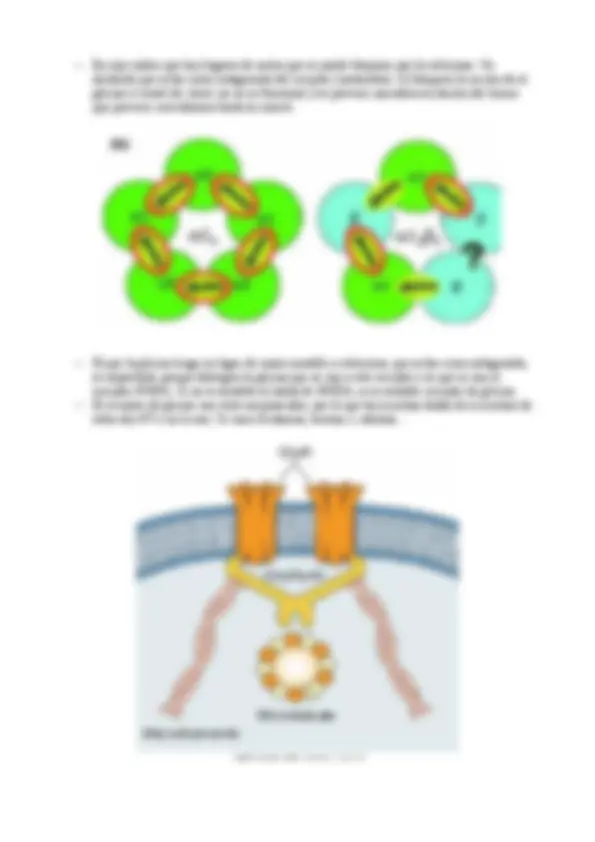
–El transportador vesicular de GABA es diferente de los de aminas.
–Parece tener 10 en lugar de 12 dominios intramembranales.
–Algunas neuronas GABAérgicas no expresan el transportador, alomejor existe otro
–Es el mismo transportador que el de la glicina
–Los receptores presinapticos son del tipo GABA-B (autorreceptores), los postsinápticos son
los GABA-A (y también GABA-B en menor concentración). El GABA-B es un receptor