¡Descarga HEMATOPOYESIS, INMUNOLOGÍA y más Apuntes en PDF de Inmunología solo en Docsity!
HEMATOPOYESIS:
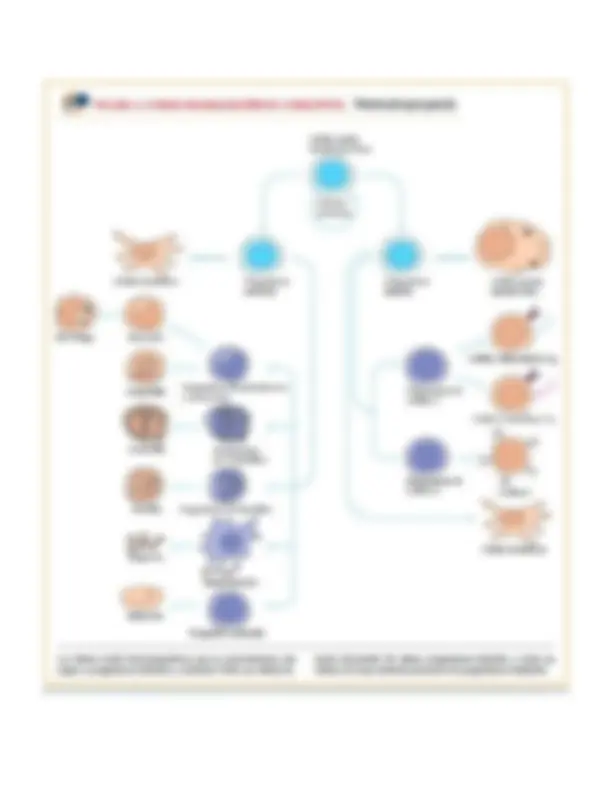
La hematopoyesis durante los primeros tiempos de la embriogénesis ocurre en el saco vitelino embrionario y posteriormente en el hígado. Durante el 3ro a 7mo mes de la gestación ocurre sobre todo en el bazo y momentos antes de nacimiento cambio a la cavidad de tuétano y de nacimiento adelante ocurre sobre todo en la médula Todas las células sanguíneas provienen de un tipo de célula llamada célula madre hematopoyética (HSC, del inglés hematopoietic stem cell). Las células madre pueden diferenciarse en otros tipos celulares; se renuevan por sí mismas mantienen su población por división celular. En el ser humano la hematopoyesis, o sea la formación y desarrollo de glóbulos rojos y blancos, se inicia en el saco vitelino embrionario durante las primeras semanas del desarrollo. Las células madre del saco vitelino se diferencian en células eritroides primitivas que contienen hemoglobina embrionaria. Hacia el tercer mes de la gestación las células madre hematopoyéticas migran del saco vitelino al hígado fetal y a continuación al bazo; estos dos órganos tienen funciones mayores en la hematopoyesis desde el tercero al séptimo meses del embarazo. Después de este período, la diferenciación de la HSC en la médula ósea se constituye en el principal factor de la hematopoyesis, y hacia el nacimiento ésta es escasa o nula en hígado y bazo. En la fi gura 2-1 se muestra la migración de la hematopoyesis durante el desarrollo desde una serie de sitios primitivos hasta la médula ósea
Es notable que toda célula sanguínea madura funcional-mente especializada deriva de una HSC. Debido a su capacidad de renovarse por sí mismas (autorrenovarse), las células madre hematopoyéticas se mantienen en concentraciones estables durante toda la vida adulta; empero, cuando aumenta la demanda de hematopoyesis, las HSC muestran una enorme capacidad de proliferación. En un momento temprano de la hematopoyesis, una célula madre multipotente se diferencia a lo largo de una de dos vías y da lugar a una célula progenitora linfoide o una célula progenitora mieloide (fi g. 2-2). Las células progenitoras han perdido la capacidad de renovarse por sí mismas y están comprometidas respecto de un linaje celular particular. Las células progenitoras linfoides dan lugar a células B, T y NK (asesinas naturales). Las células madre mieloides generan pro-genitoras de glóbulos rojos (eritrocitos), muchos de los diversos glóbulos blancos (neutrófilos, eosinófilos, basófilos, monocitos, células cebadas, células dendríticas) y células generadoras de plaquetas llamadas megacariocitos. En la médula ósea las células hematopoyéticas y sus descendientes crecen, se diferencian y maduran en un andamiaje parecido a red de células del estroma o estromales, entre las que se incluyen células adiposas (adipocitos), células endoteliales, fibroblastos y macrófagos. Las células estromales influyen en la diferenciación de células madre hematopoyéticas al proporcionar un microambiente inductor hematopoyético (HIM, del inglés hematopoietic inducing microenvironment) que consiste en una matriz celular y factores que promueven el crecimiento y la diferenciación. Muchos de estos factores de crecimiento hematopoyéticos son agentes solubles que llegan a sus células blanco por difusión; otros son moléculas unidas a membrana en la superficie de células estromales que requieren contacto célula a célula entre las células que responden y las estromales Durante la hematopoyesis, los eritrocitos y muchos tipos distintos de glóbulos blancos descienden de las pocas células madre hematopoyéticas en un proceso complicado que incluye muchos pasos los cuales atraviesan por una jerarquía de poblaciones precursoras. ¿Por qué surgió por evolución tal proceso complejo para generar células sanguíneas? En el curso de su vida, una persona producirá unas 1016 de tales células. Esto hace necesario una enorme cantidad de divisiones celulares. John Dick ha señalado que la división celular es propensa a errores y da la oportunidad para que el genoma sufra mutaciones, algunas de las cuales pueden producir cáncer. Él ha sugerido que para hacer menos probable este suceso potencialmente catastrófico, el sistema formador de sangre está organizado en una ingeniosa jerarquía en la cual la mayor parte de la proliferación ocurre dentro de precursores más diferenciados y no en la población misma de células madre hematopoyéticas. Estos precursores en diferenciación progresiva no se autorrenuevan, y las células sanguíneas maduras que forman son incapaces de dividirse o sólo lo hacen en circunstancias especiales. En consecuencia, la posibilidad de generar cáncer en las HSC y sus descendientes inmediatos se reduce, si bien no a cero.
La hematopoyesis se regula a nivel genético El desarrollo de células madre hematopoyéticas pluripotentes en distintos tipos de células requiere la expresión de distintos grupos de genes determinantes y específicos de linaje en los tiempos apropiados y en el orden correcto. Las proteínas especificadas por estos genes son componentes críticos de las redes reguladoras que dirigen la diferenciación de la célula madre y su descendencia. Gran parte de lo que se conoce en la actualidad sobre la dependencia de la hematopoyesis respecto de un gen particular proviene de estudios en ratones en los que se desactiva o altera (en inglés knock out, “apagar” o “noquear”) un gen por alteración dirigida, lo que bloquea la producción de la proteína que dicho gen codifica (cap. 22). Si los ratones no producen glóbulos rojos o blancos específicos cuando se desactiva un gen, se concluye que es necesaria la proteína especificada por el gen para el desarrollo de dichas células. La tecnología de desactivación de genes (knockout) es uno de los instrumentos más potentes disponibles para determinar las funciones de genes particulares en una amplia gama de procesos y ha hecho contribuciones importantes a la identificación de muchos genes que regulan la hematopoyesis. Aunque aún queda mucho por hacer, la desactivación dirigida y otros métodos han identificado varios factores de transcripción, que tienen actividades relevantes en la hematopoyesis (cuadro 2-1). Algunos de estos factores de transcripción afectan muchos diferentes linajes hematopoyéticos y otros sólo un linaje aislado, como la vía de desarrollo que conduce a linfocitos. Un factor de transcripción que afecta múltiples linajes es el GATA-2, un miembro de una familia de factores de transcripción que reconoce la secuencia tetranucleótido GATA, un motivo presente en genes blanco. Para el desarrollo de los linajes linfoide, eritroide y mieloide es esencial un gen GATA-2 funcional, que especifica este factor de transcripción. Como cabría esperar, los animales en los que se altera este gen mueren durante el desarrollo embrionario. En contraste con el GATA-2, otro factor de transcripción, Ikaros (Ícaro), sólo es necesario para el desarrollo de células de linaje linfoide. Aunque los ratones alterados Ikaros no producen cifras considerables de cé-lulas B, T y NK, su producción de eritrocitos, granulocitos y otras células del linaje mieloide no se modifica. Los ratones alterados Ikaros sobreviven al desarrollo embrionario, pero son muy deficientes en sentido inmunitario y mueren por infecciones a una edad temprana. Otro regulador de la transcripción más, Bmi-1, es un represor transcripcional que constituye un determinante clave de la capacidad de las HSC de autorrenovarse. Cuando se desactiva el gen que codifica Bmi-1 (el cual es altamente expresado en las HSC de ser humano y ratón), los ratones deficientes en Bmi-1 mueren en un plazo de dos meses después de nacer. La causa de la muerte es la incapacidad final de la médula ósea de generar glóbulos rojos y blancos. Esta incapacidad de la médula ósea se rastreó hasta la falta de autorrenovación de las HSC.
BIBLIOGRAFIA:
- Richard A. Goldsby, Barbara A. Osborne y Th omas J. Kindt. INMUNOLOGIA DE Kuby. 6ta Edición. Mexico.
- Abul K. Abbas, Andrew H. k Lichtman, Shiv Pillai. INMUNOOGIA CELULAR Y MOLECULAR. 7ta edicion. España. Gea Consultoria Editorial, S.L. 2012