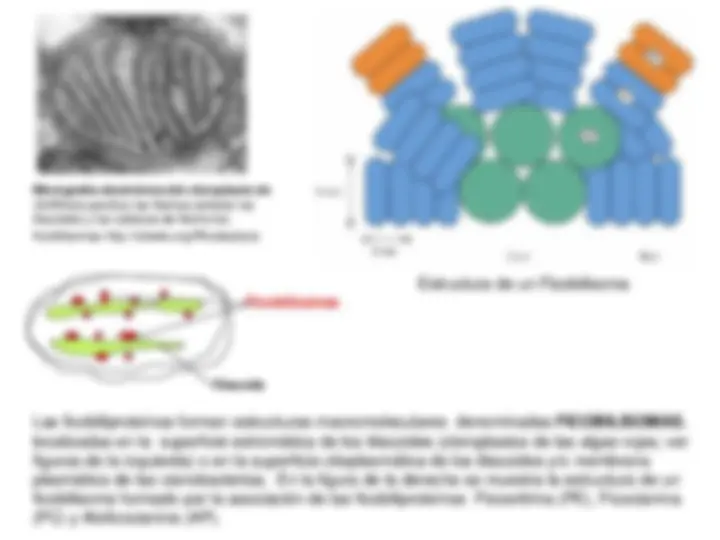
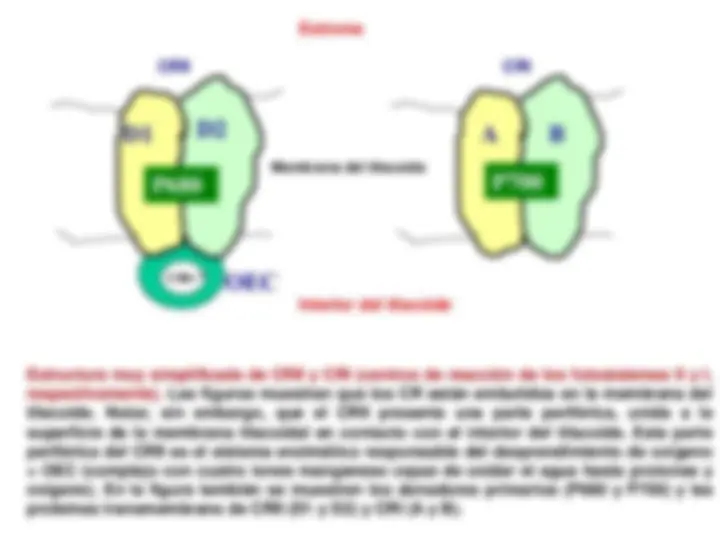
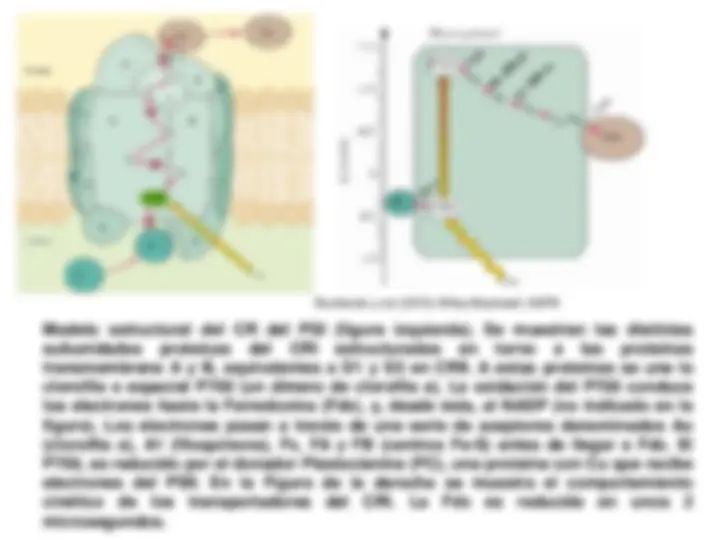
¡Descarga Fase luminosa de la fotosintesis y más Diapositivas en PDF de Física solo en Docsity!
Introducción general a la Fotosíntesis. Aparato fotosintético.
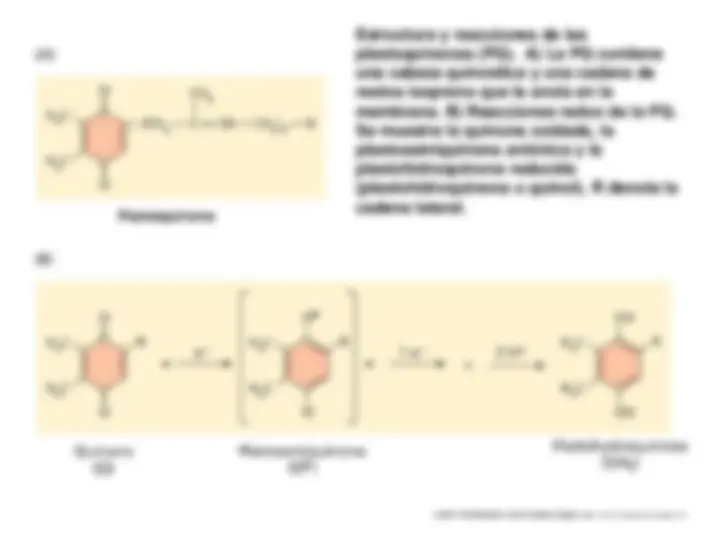
Pigmentos fotosintéticos: clorofilas, carotenoides y ficobilinas:
Organización de los pigmentos en los Tilacoides.
Utilización de la energía luminosa. Fotosistemas: Antenas y
Centros de Reacción. Absorción y Transferencia de Energía en los
fotosistemas.
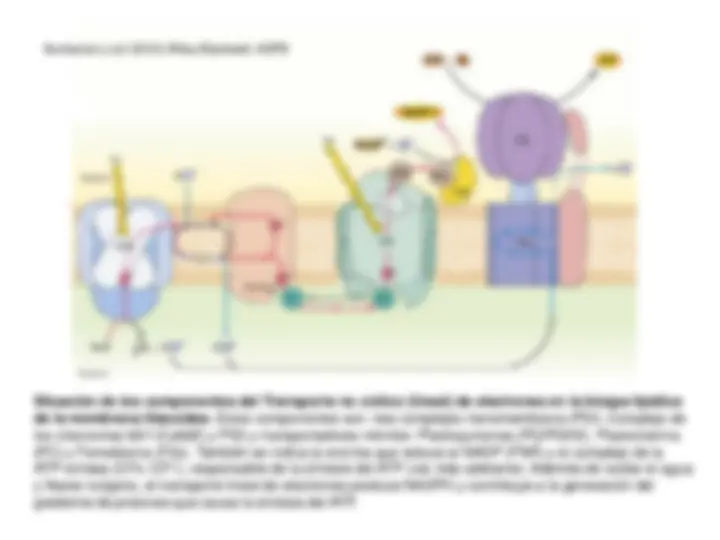
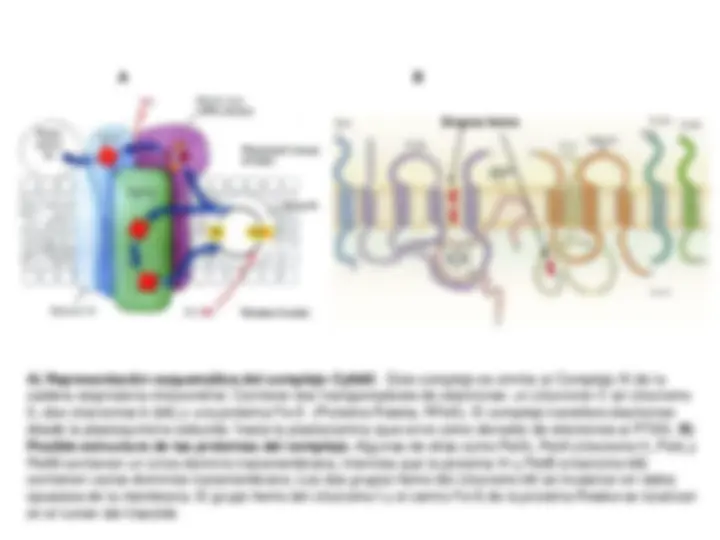
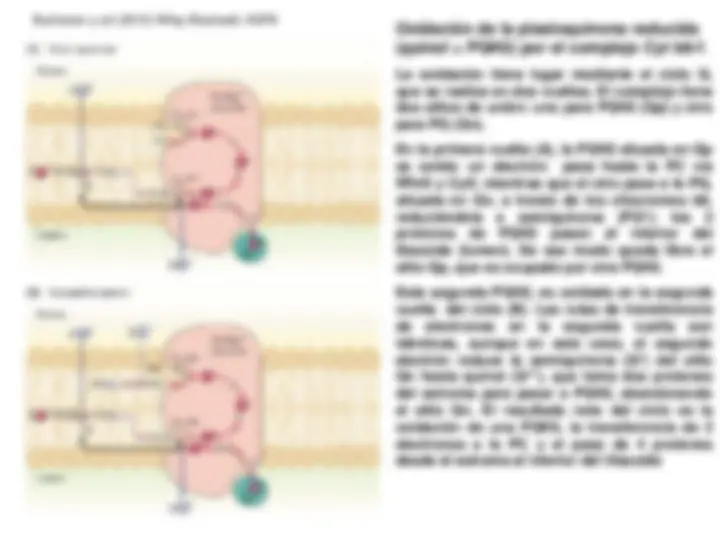
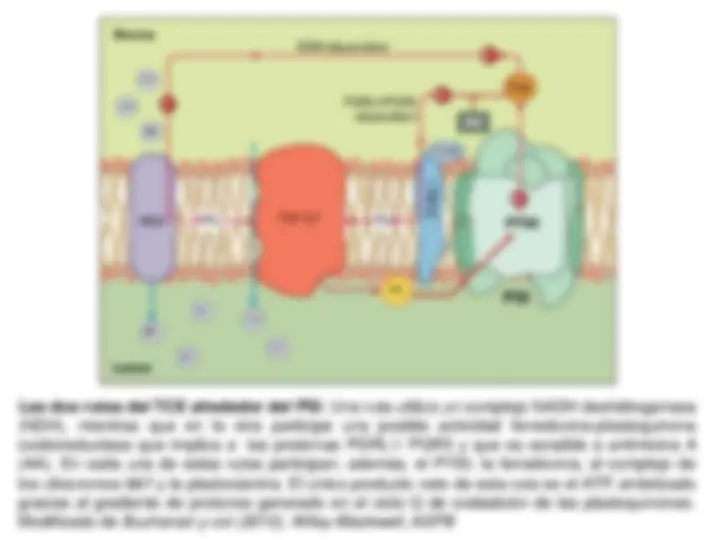
Sistema fotosintético de transporte de electrones y
Fotofosforilación. Esquema en Z. Transporte electrónico
No-Cíclico (Lineal), Cíclico y Pseudocíclico. Mecanismo de
fotofosforilación.
Protección del aparato fotosintético
FASE LUMINOSA DE LA FOTOSINTESIS
La FOTOSINTESIS
En una primera aproximación, puede definirse como el conjunto de procesos por los que las plantas, algas y algunas bacterias utilizan directamente la energía luminosa para sintetizar compuestos orgánicos. Además de suministrar alimentos, biomasa y combustibles fósiles, la fotosíntesis en las plantas, algas y cianobacterias es oxigénica (produce oxígeno), requerido para la actividad respiratoria de muchos organismos y para la formación de la capa de ozono, que protege al planeta Tierra de las radiaciones de longitud de onda corta, incompatibles con la vida. La fotosíntesis que realizan las bacterias fotosintéticas es anoxigénica (no produce oxígeno).
La Fotosíntesis es el proceso de síntesis mas importante que se realiza en la Tierra. En un año, los organismos fotosintéticos fijan aproximadamente 1.0 x 10^14 Kg (100* billones de toneladas) de carbono en materia orgánica (Productividad neta primaria). Esta enorme cantidad de materia orgánica es mas o menos equivalente a un "árbol de carbohidratos" de 10 m de altura y 100 Km de diámetro. También representa el 1% de las reservas mundiales conocidas de crudos fósiles (carbón, gas, petróleo) o 10 veces el consumo energético mundial. La fuente de carbono utilizada en la fotosíntesis es el CO 2 de la atmósfera (0.04% = 8 x 10^14 Kg de carbono) o el HCO 3 -^ disuelto en lagos y océanos (400 x 10^14 Kg de carbono).
*se refiere al billón americano (1000 millones)
H 2 O
Energía luminosa
CO 2
NO 3 -
SO 4 2-^ Compuestos orgánicos
O 2
Destino de la Energía solar (*) que llega a la superficie de la tierra
Balance global Cantidad en Terawatios a
Entrada de energía solar b^ 178.
Reflejada al espacio de forma inmediata 53.
Absorbida y reflejada como calor 82.
Utilizada para evaporar el agua 40.
Capturada por la fotosíntesis (productividad neta primaria)c^100
Energía total utilizada por la humanidad:
En 2005 13
Esperada en 2100 46
Total utilizada como alimento 0.
a 1 watio = 1 julio/unidad de tiempo. 1 Terawatio = 10 12 watios = 10 12 julios s -
b La entrada de energía solar por año es 5.62 x 10 12 terawatios (5.62 x 10 24 julios)
c Energía solar capturada por los organismos fotosintéticos en un año: 3.16 x 10 9
terawatios (3.16 x 10 21 julios)
(*) La energía solar es el producto de reacciones de fusión nuclear que convierten
los protones en núcleos de helio con una intensidad aproximadamente
equivalente a10 17 kg de TNT por segundo
Fuente: The molecular life of plants (2013) Wiley-Blackwell-American Society of Plant Biologists
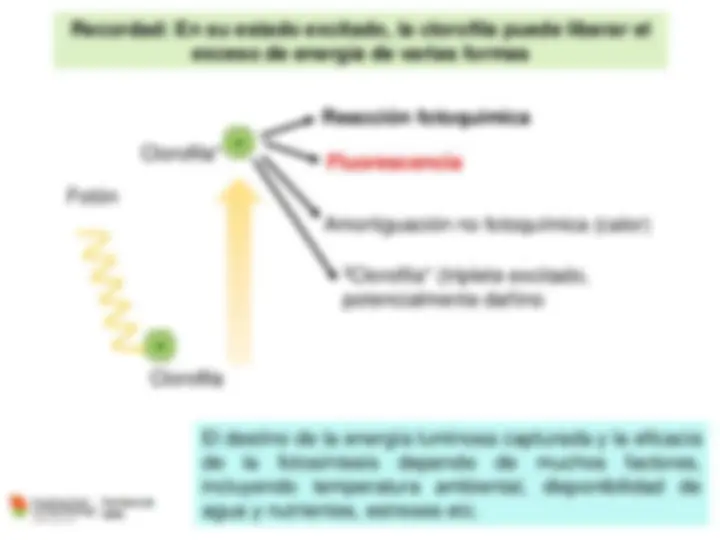
Cuando se propaga, la luz se comporta como una onda, mientras que cuando interactúa con la materia se comporta (o tienes propiedades) de partícula.
Como onda, se propaga a una velocidad ( c ), frecuencia ( ν ) y longitud de onda ( λ ) características. Las tres magnitudes se relacionan de acuerdo con la ecuación:
c = λ ν
Cuando interactúa con la materia, la luz se comporta como si estuviera dividida en unidades discretas (partículas) denominadas FOTONES, que contienen una cantidad discreta de energía (QUANTUM, plural QUANTA). La energía (E) de un fotón (el cuanto) se expresa por la ecuación:
E = h υ = hc / λ (1)
h = constante de Planck (6.626 x 10-34^ Julios. s); c = 3.0 x 10^8 m/s; ν (ciclos. s), λ (m)
En lugar de calcular la Energía/fotón, es mas aconsejable calcular la energía por mol de fotones, multiplicando el término hc de la ecuación (1) por el número de Avogadro N (6.022 x 1023 ). La energía de un mol de fotones se denomina, a veces, 1 einstein.
La luz es una onda electromagnética transversal con los campos eléctrico y magnético perpendiculares entre si y a la dirección de propagación
La curva A es la energía emitida por el sol en función de la longitud de onda, que incide sobre la atmósfera exterior. La curva B es la energía que incide sobre la superficie de la tierra. Las depresiones en la región del infra-rojo por encima de 700 nm indican la absorción de energía solar por moléculas de la atmósfera, principalmente vapor de agua y CO 2. La curva C, es el espectro de absorción () de la clorofila, que absorbe en las regiones del azul y rojo del espectro visible. Dado que la luz verde del espectro visible no es absorbida, la mayoría es reflejada y da a las plantas su característico color verde. () Los espectros de absorción nos dan información sobre la cantidad de energía absorbida por una molécula en función de la longitud de onda de la luz.
Distribución de las longitudes de onda de los fotones solares que inciden sobre la
atmósfera terrestre y su superficie. La curva de la irradiación solar es una idealización
basada en la fórmula de la distribución de la radiación de Planck. La distribución espectral y la
cantidad de irradiación que llega a la superficie terrestre depende de la nubosidad, otras
condiciones atmosféricas, altitud y ángulo solar en el cielo.
Efectos del vapor de agua y vapor dióxido de carbono
Efecto del ozono
Radiación solar que incide sobre la atmósfera externa
Radiación solar que alcanza la superficie terrestre
Espectro de absorción de la clorofila
Flujo fotónico por unidad de longitud de onda
Longitud de onda,
Espectro visible
Figura tomada de Taiz y col (2015) Plant Physiology and Development, Sinauer; modificada a partir de Nobel (1999) Physicochemical & Environmental Plant Physiology, 2nd ed. Academic press
Diagrama ilustrativo de las relaciones entre los tipos principales de plastos. Todos los plastos derivan de los proplastos, que son transmitidos de una generación a la siguiente por vía paterna o materna (depende de las especies; ver diapositiva siguiente) y son mantenidos en los meristemos. Las flechas con línea continua indican los pasos normales en el desarrollo de los plastos; las flechas con línea discontinua indican conversiones que sólo aparecen en circunstancias especiales. La gran plasticidad en el desarrollo de los plastos ha conducido a confusiones en su nomenclatura. Por ejemplo, el término leucoplasto se ha utilizado de tres formas diferentes: como genérico de todos los tipos de plastos sin pigmentos, como nombre de un precursor de un tipo de plasto que origina ciertos tipos de plastos sin pigmentos, o como nombre de un plasto sin pigmentos que produce aceites esenciales (plastos de las células secretoras de los tricomas glandulares). El etioplasto es el plasto de las plantas crecidas en oscuridad (etioladas); al iluminar, se transforma en cloroplasto (ver diapositiva siguiente)
Biochemistry and molecular biology of plants (2015) Wiley/Blackswell
Los organismos eucariotas fotosintéticos
(plantas y algas) realizan la fotosíntesis en los
cloroplastos, miembros de una familia de
orgánulos celulares exclusivos de las células
vegetales (los plastos).
Los distintos tipos de plastos derivan del proplasto (proplastidio), también conocido como eoplasto en las plantas terrestres. Los etioplastos son los progenitores de los cloroplastos, mientras que los gerontoplatos son los cloroplastos senescentes. Los desicoplastos son plastos que pueden sufrir inter-conversiones entre cloroplastos y proplastos en plantas tolerantes a la desecación. Los cromoplastos sufren interconversiones con cloroplastos y proplastos, y sintetizan y almacenan carotenoides en frutos, flores, y algunos tejidos de raíces. Varios tipos de plastos se han clasificado como plastos sin color, y genéricamente se conocen como leucoplastos, aunque se subdividen en distintos tipos de acuerdo con las moléculas que sintetizan y/o almacenan: Elaioplastos, que almacenan lípidos en plastoglubulos, o aleuroplastos, que almacenan proteínas; Amiloplastos, que almacenan gránulos de almidón, predominantemente en las raíces. Además de estos tipos, relativamente bien descritos, recientemente se han añadido los taninosomas, derivados de los cloroplastos, los feniloplastos, que sintetizan/almacenan fenil-glucósidos, y los xiloplastos, derivados de proplastos o amiloplastos, presentes en los tejidos vasculares secundarios, especializados en la síntesis de precursores de la producción de monolignoles. Pinard and Mizrachi (2018) Unsung and understudied: plastids involved in secondary growth. Current Opinion in Plant Biology 42:30–
Actualización de los tipos de plastos
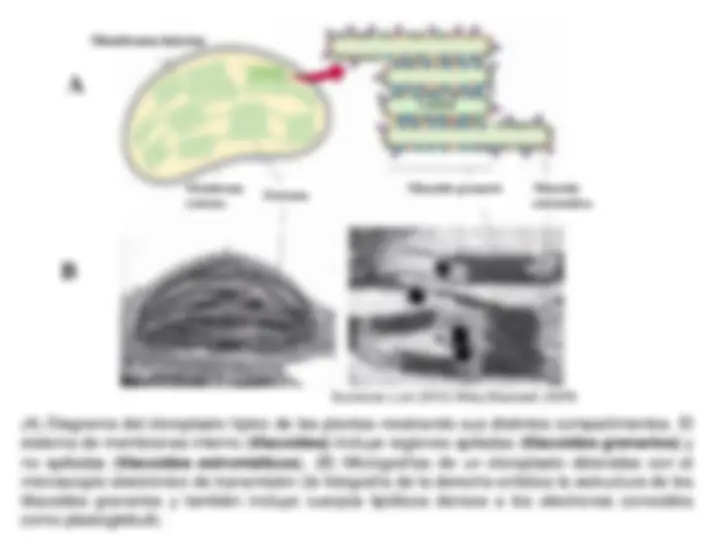
Estructura simplificada de un cloroplasto. Los cloroplastos están rodeados por una doble
membrana (cubierta plastidal); la membrana interna limita una matriz (el estroma) en la
que se sitúa el sistema membranoso interno (los tilacoides).Los tilacoides son sáculas que
delimitan un espacio interno o lumen. En las membranas de los tilacoides se localizan los
pigmentos fotosintéticos, transportadores de electrones y ATP-sintasa. En las plantas, se
distinguen dos tipos de tilacoides, los granarios (apilados, formando granum) y los
estromáticos (no apilados). En el estroma también se sitúan granos de almidón,
plastoglóbuli (acumulaciones lipídicas), el material genético (ADN, ARN) y los
ribosomas.
ADN
Ribosomas y ARNm
Grano de almidón Plastoglobuli
Granum Tilacoide granario
Tilacoide estromático
Estroma
Membrana externa
Membrana interna
Cubierta plastidal
Lumen de los tilacoides
( A) Diagrama del cloroplasto típico de las plantas mostrando sus distintos compartimentos. El
sistema de membranas interno (tilacoides) incluye regiones apiladas (tilacoides granarios) y
no apiladas (tilacoides estromáticos). (B) Micrografías de un cloroplasto obtenidas con el
microscopio electrónico de transmisión (la fotografía de la derecha enfatiza la estructura de los
tilacoides granarios y también incluye cuerpos lipídicos densos a los electrones conocidos
como plastoglobuli).
Membranaexterna Estroma Tilacoide granario
Membrana interna
Tilacoide estromático
A
B
Lumen
Buchanan y col (2015) Wiley-Blackwell, ASPB
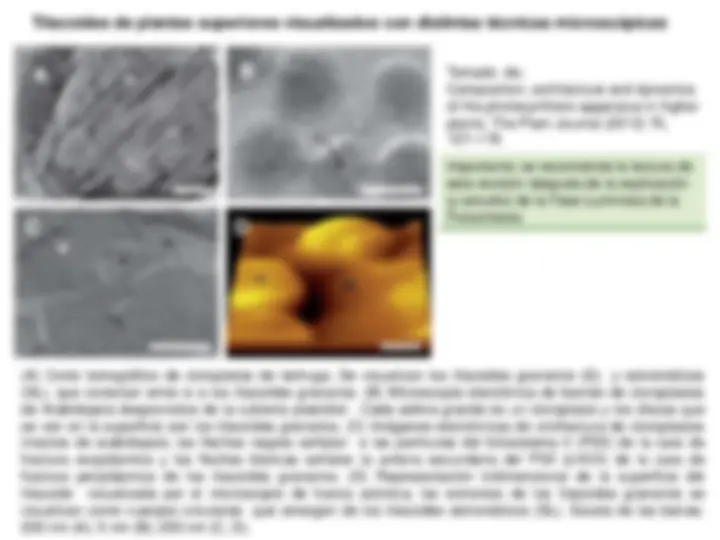
Tilacoides de plantas superiores visualizados con distintas técnicas microscópicas
(A) Corte tomográfico de cloroplatos de lechuga. Se visualizan los tilacoides granarios (G) y estromáticos (SL), que conectan entre si a los tilacoides granarios. (B) Microscopía electrónica de barrido de cloroplastos de Arabidopsis desprovistos de la cubierta plastidial. Cada esfera grande es un cloroplasto y los discos que se ven en la superficie son los tilacoides granarios. (C) Imágenes electrónicas de criofractura de cloroplastos intactos de arabidopsis; las flechas negras señalan a las partículas del fotosistema II (PSII) de la cara de fractura exoplásmica y las flechas blancas señalan la antena secundaria del PSII (LHCII) de la cara de fractura periplásmica de los tilacoides granarios. (D) Representación tridimensional de la superficie del tilacoide visualizada por el microscopio de fuerza atómica; los extremos de los tilacoides granarios se visualizan como cuerpos circulares que emergen de los tilacoides estromáticos (SL). Escala de las barras: 200 nm (A); 5 nm (B); 250 nm (C, D).
A B
C D
Tomado de: Composition, architecture and dynamics of the photosynthetic apparatus in higher plants. The Plant Journal (2012) 70, 157–
Importante: se recomienda la lectura de esta revisión después de la explicación (y estudio) de la Fase Luminosa de la Fotosíntesis
I) (A) Visión general. Los tilacoides granarios (G) están interconectados por múltiples tilacoides estromáticos (SL). Estos últimos también establecen algunas conexiones con la membrana interna (EM) de la cubierta plastidal (ver recuadro). Los ribosomas aparecen como partículas muy teñidas; PG denota los plastoglobuli. (B) Ampliación del ensamblaje granum- estroma. Los tilacoides granarios, mostrados en varios colores, se entrecruzan con el cuerpo del granum. Notar que las superficies apicales y basales de los granum son lisas, mientras que las superficies de las zonas apresadas aparecen rugosas (granuladas).
II) Organización tridimensional del ensamblaje de los tilacoides granarios y estromáticos. La estructura se generó a partir de los datos tomográficos de la figura anterior. (A) Los tilacoides granarios son contiguos con los estromáticos que bifurcan en la interfase granum- estroma. Las conexiones internas entre capas adyacentes se indican mediante una flecha y una cabeza de flecha. G, granum; SL, tilacoides estromáticos.
I
II
Información suplementaria
Información adicional sobre los pastoglobuli (PG)
Los PG están rodeados de una monocapa de lípidos polares contigua con los lípidos de la
superficie externa de los tilacoides (ver diapositiva siguiente). La zona central del PG contiene
lípidos neutros, como prenilquinonas (tocoferol, filoquinona, plastoquinona, carotenoids y
triacilglicéridos). La superficie de los PG contiene proteínas, como las fibrilinas (FBN),
conocidas previamente como plastoglobulinas (PGL). Las FBNs son proteínas estructurales
que también pueden estar implicadas en varias rutas metabólicas de los lípidos.
El número, tamaño y contenido lipídico de los PGs cambia con las condiciones ambientales y
el estado de desarrollo de la planta. A nivel ultraestructural, estos cambios se correlacionan
con la remodelación del contenido lipídico de la membrana de los tilacoides. Los PG participan
en la remodelación metabolizando y almacenado moléculas. Su función en el tráfico desde y
hacia los tilacoides se infiere de la existencia de sitios de contacto entre tilacoides y PGs, que
pueden servir como conductos para las moléculas lipídicas.
Recientemente, se ha demostrado que los PGs están implicados en la biosíntesis y
metabolismo de las vitaminas E (Tocoferol) y K (Filoquinona). Ambos tipos de compuestos
pertenecen a la familia las prenilquinonas, que también incluye a las plastoquinonas. Son
moléculas solubles en lípidos que actúan como antioxidantes impidiendo la peroxidación de
los lípidos en la membrana de los tilacoides y amortiguando especies reactivas de oxígeno
(ROS). La filoquinona y plastoquinona también actúan como transportadores de electrones en
el PSI y PSII, respectivamente.
Mas información en: Unexpected roles of plastoglobulis (plastid lipid droplets) in vitamin K
and E metabolism. Current Opinion in Plant Biology 2015, 25:123–
Biochemistry and molecular biology of plants (2015) Wiley/Blackswell
Los plastoglobuli se asocian con las membranas de los tilacoides
A) Modelo tomográfico tridimensional de las membranas de los tilacoides asociadas a plastoglóbuli. Se muestran tilacoides granarios (gt), tilacoides estromáticos (st) y plastoglóbuli (pg).
B) Diagrama de un plastoglóbuli unido a la membrana del tilacoide. La monocapa lipídica que rodea al plastoglóbuli es continua con la capa externa de la membrana del tilacoide y contiene proteínas estructurales y enzimáticas. El dominio funcional de estas enzimas se localiza, mayoritariamente, hacia el interior del plastoglóbuli.
Información sobre los estrómulos
Los estrómulos son extensiones tubulares de los plastos cuya función esta comenzando a conocerse.
Se asume que los estrómulos intercambian señales y metabolitos entre los plastos y otros orgánulos y que son inducidos durante el estrés. Utilizando microscopia confocal y de super-resolución tridimensional se ha demostrado que los estrómulos se forman dentro de los cloroplastos en respuesta a señales redox sensibles a la luz. Su frecuencia incrementa durante el día o después de tratamientos con productos químicos que producen ROS en los cloroplastos. Los cloroplastos aislados pueden hacer estrómulos lo que sugiere que los factores asociados con los cloroplastos son suficientes para generar estrómulos. En la figura de la derecha (PNAS 112: 10044–10049, 2015) se muestra como la frecuencia de formación de estrómulos en cloroplastos de Nicotiana benthamiana incrementa durante el día (barras amarillas en A) y disminuye por la noche (barras azules en A). En B y C se muestran imágenes representativas de cloroplastos marcados con GFP (verde) durante el día (B) y la noche (C). Barra: 10 μ m.
Estrómulos tubulares de dos cloroplastos que emiten fluorescencia roja y verde. Los dos plastos contienen la misma proteína verde fluorescente (GFP), pero el de la izquierda fue irradiado para que la GFP emitiera fluorescencia roja. Biochemistry and molecular biology of plants (2015) Wiley/Blackswell