¡Descarga pdf tesis erizo de mar y más Guías, Proyectos, Investigaciones en PDF de Biología solo en Docsity!
FEBRERO DEL 2010
No 04- 2010 - 101813383300 - 102 ®
LOS GAMETOS DE ERIZO DE MAR COMO MODELO DE ESTUDIO
DE LA FECUNDACIÓN
THE GAMETES OF SEA URCHIN AS A MODEL FOR THE STUDY
OF FERTILIZATION
Laura Edith Castellano-Torres^1 , Guadalupe Martínez-Cadena^2 , Juana López-Godínez^2 , Patricia Cuéllar-Mata^2 y J. de Jesús García-Soto^2
(^1) Departamento de Ciencias Aplicadas al Trabajo, División de Ciencias de la Salud, Campus
León, Universidad de Guanajuato, Av. Eugenio Garza Sada No. 572, Col. Lomas del Campestre sección 2, León, Gto. CP 37150. Correo electrónico [email protected].
(^2) Departamento de Biología, División de Ciencias Naturales y Exactas, Campus Guanajuato,
Universidad de Guanajuato, Noria Alta S/N, Col. Noria Alta, Guanajuato, Gto. CP 36050.
ABSTRACT
The sea urchin gametes are excellent models to study the molecular basis of fertilization and embryonic development. Studies by several research groups and ours have allowed understanding some of the molecular mechanisms that regulate both the physiolo gy of sperm and egg and early development of sea urchin.
KEYWORDS: Spe rm, egg, fertilization
RESUMEN
Los gametos de erizo de mar son excelentes modelos para estudiar las bases moleculares de la fecundación y del desarrollo embrionario. Estudios realizados por varios grupos de investigación así como el nuestro han permitido entender algunos de los mecanismos moleculares que regulan tanto la fisiología del espermatozoide y del óvulo como el desarrollo temprano del erizo de mar.
PALABRAS CLAVE: Espe rmatozoide, óvulo, fecundación
FEBRERO DEL 2010
No 04- 2010 - 101813383300 - 102 ®
INTRODUCCIÓN
La reproducción sexual involucra la recombinación del genoma de dos individuos de la misma especie para producir otro con características genéticas similares a sus progenitores. Para que la fecundación se lleve a cabo deben participar los dos gametos, el masculino o espermatozoide y el femenino u óvulo. Con la unión de ambos células, y la consiguiente formación del cigoto, comienza el desarrollo embrionario, sin lugar a dudas una de las áreas más fascinantes de la biología reproductiva, con significativas implicaciones médicas, sociales y económicas. Si bien los avances en este tema son extraordinarios, falta aun mucho por aprender y aplicar el conocimiento adquirido en las áreas que competen preferentemente a la salud.
Una vez ocurrida la fecundación, el cigoto recién formado ejecuta su programa de desarrollo mediante la realización de múltiples procesos celulares, de los cuales poco se conocen a nivel molecular (Leguia y Wessel, 2007). El desarrollo embrionario, comenzando con la activación del óvulo por el espermatozoide, es controlado por elaborados mecanismos de transducción de señales. En la célula, los sistemas de vías de señalización están constituidos por diversos elementos que de manera coordinada facilitan el flujo de información para la realización de un determinado proceso celular, propio de su estado de diferenciación. Tales elementos son, entre otros, receptores en la membrana plasmática, proteínas G o GTPasas, efectores como la adenilil ciclasa, fosfolipasa C, canales iónicos, fosfotransferasas así como segundos mensajeros, y cambios en el pH intracelular (pHi). Para abordar a un nivel básico la comprensión de los mecanismos de transducción de señales en el desarrollo embrionario, los gametos de invertebrados marinos han constituido modelos muy apropiados. Tal es el caso de los gametos de erizos de mar, cuyo empleo para el estudio de diversos temas de la biología básica y de la biología del desarrollo impulsó, hace algunos años, la secuenciación de su genoma, particularmente de la especie Strongylocentrotus purpuratus (Sodergen y col., 2006). Lo anterior, como era previsible, ha tenido un enorme impacto, contribuyendo a responder preguntas fundamentales sobre el desarrollo embrionario. Además, es significativo que los embriones
FEBRERO DEL 2010
No 04- 2010 - 101813383300 - 102 ®
tiene la maquinaria metabólica necesaria para el desarrollo de un nuevo individuo. El óvulo está rodeado de la membrana plasmática (MP) y de dos capas externas, la capa de gelatina (CG) y la capa vitelina (CV) formadas por glicoproteínas (SeGall y Lennarz, 1981). La más externa es la CG, que mide ~40 μm de grosor y está compuesta de un polímero de fucosa sulfatada (FSP, por sus siglas en inglés fucose sulfate polymer ) y por una sialoglicoproteína (Mourao y col., 2007). El FSP es el responsable de los cambios morfológicos y fisiológicos que ocurren en el espermatozoide (Vacquier y Moy, 1997). La CG también contiene pequeños péptidos que tienen efectos muy importantes en la fisiología del espermatozoide ya que lo preparan y lo guían hacia el óvulo (Beltrán y col., 2007; Darszon y col., 2006, 2008).
La CV se encuentra unida a la superficie externa de la MP. Es una cápsula delgada, resistente y elástica que sirve como protección durante el desarrollo embrionario. Contiene los receptores glicoproteicos a los que se unen los espermatozoides homólogos. Después de la CV, se encuentra la MP, la cual se fusiona con la MP del espermatozoide durante el proceso de la fecundación. Por debajo de la MP del óvulo, están los gránulos corticales (GC) que se derivan del aparato de Golgi: contienen una mezcla de enzimas, proteínas estructurales y materiales coloidales, como mucopolisacáridos sulfatados (Carroll, 1976).
FEBRERO DEL 2010
No 04- 2010 - 101813383300 - 102 ®
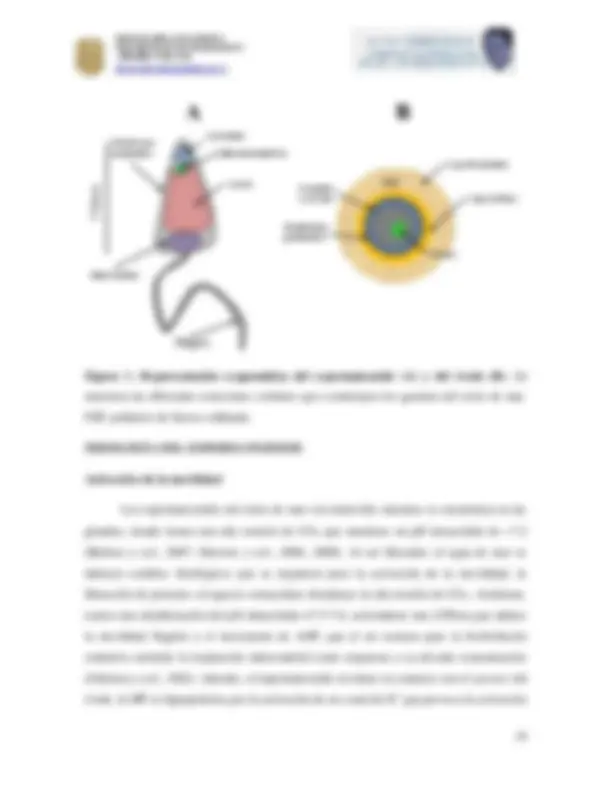
Figura 1. Representación esque mática del espermatozoide (A) y del óvulo (B). Se muestran las diferentes estructuras celulares que constituyen los gametos del erizo de mar. FSP, polímero de fucosa sulfatada.
FISIOLOGÍA DEL ESPERMATOZOIDE
Activación de la movilidad
Los espermatozoides del erizo de mar son inmóviles mientras se encuentran en las gónadas, donde tienen una alta tensión de CO 2 que mantiene un pH intracelular de 7. (Beltran y col., 2007; Darszon y col., 2006, 2008). Al ser liberados al agua de mar se inducen cambios fisiológicos que se requieren para la activación de la movilidad; la liberación de protones al espacio extracelular disminuye la alta tensión de CO 2. Asimismo, ocurre una alcalinización del pH intracelular 7.5-7.6, activándose una ATPasa que induce la movilidad flagelar y el incremento de ADP, que al ser sustrato para la fosforilación oxidativa estimula la respiración mitocondrial como respuesta a su elevada concentración (Christen y col., 1982). Además, el espermatozoide al entrar en contacto con el speract del óvulo, la MP se hiperpolariza por la activación de un canal de K+^ que provoca la activación
FEBRERO DEL 2010
No 04- 2010 - 101813383300 - 102 ®
espermatozoide (Trimmer y col., 1985), activa diversos transportadores en la MP. Estos transportadores facilitan flujos iónicos para que exista un aumento de Na+^ y Ca+2^ y una salida de H+^ y K+^ (Darszon y col., 2005, 2006, 2008), que producen cambios en el potencial de la membrana y la alcalinización del citosol. También se inducen otros cambios fisiológicos en el espermatozoide por el FSP como un aumento en la concentración de trifosfato de inositol (IP 3 ) y la elevación de la concentración de AMPc como consecuencia de la activación de la adenilil ciclasa dependiente de Ca2+^ y las transmembranales (Darszon y col., 2005, 2006, 2008; Beltrán y cols., 2007b). Asimismo, se estimula una PKA, una fosfolipasa D (Darszon y col., 2005, 2006, 2008) y una óxido nítrico sintasa (Kuo y col., 2000).
ACTIVACIÓN DEL ÓVULO
La fecundación inicia con una serie de eventos que suceden de manera coordinada, iniciando con ella el programa de desarrollo del cigoto. Estos eventos, comúnmente conocidos como la activación del óvulo, ocurren de manera rápida y transitoria, comprometiendo al cigoto recién formado en procesos tales como el rearreglo de su citoesqueleto de actina, el reingreso al ciclo celular y en una formidable activación metabólica. La activación inmediata del óvulo se ejecuta principalme nte por proteínas ya existentes en esa célula, cuya localización y actividad se logra mediante recambio y modificaciones postraduccionales, particularmente la fosforilación de proteínas mediada por cinasas (Roux y col., 2008).
En el erizo de mar, y en el de todos los animales hasta ahora estudiados, un aumento del Ca2+^ citosólico es crucial en la activación del óvulo. La función del Ca2+^ es estimular la exocitosis de los gránulos corticales, reiniciar el ciclo celular y comenzar la síntesis de proteínas y DNA. El Ca2+^ se libera de depósitos internos, principalmente del retículo endoplásmico, en una acción mediada por el trifosfato de inositol (IP 3 ) (Townley y col., 2006). El IP 3 se une a un receptor en la membrana del retículo endoplásmico (RIP 3 ) provocando la liberación del Ca2+^ y su propagación como una onda en el citoplasma del óvulo; este aumento en el Ca2+^ retorna minutos después a sus niveles basales. Posteriormente se requieren señales de Ca2+, principalmente cuando el cigoto debe entrar a
FEBRERO DEL 2010
No 04- 2010 - 101813383300 - 102 ®
la mitosis y ejecutar eventos tardíos del desarrollo. Sin embargo, y si bien mucho se ha avanzado en este tema, se desconocen las bases de esta dinámica de Ca2+.
La onda inicial de Ca2+^ dispara la exocitosis de miles de vesículas (gránulos corticales) adheridas a la cara interna de la MP. Este proceso tiene como finalidad depositar en la superficie celular una batería de enzimas y materiales diversos para transformar la CV del óvulo – de naturaleza glicoproteica- en una estructura más resistente que contribuye a bloquear la polispermia. Estas vesículas secretoras migran a la región cortical durante la etapa final de la oogénesis, después de que se generan en el aparato de Golgi, confiriéndole al óvulo la capacidad para ser fecundado sin riesgo de polispermia; otros grupos y el nuestro han demostrado que la vía de transducción que regula este tráfico vesicular es dependiente del citoesqueleto de actina y proteínas G de la superfamilia de Ras (Covián- Nares y col., 2004). Además, la exocitosis de los gránulos sirve para depositar proteínas específicas en la membrana del cigoto que se ha formado (Rangel-Mata y col., 2007), ayudando al recambio de sus componentes de membrana. En la fusión de estas vesículas con la MP, el Ca2+^ es un mediador importante así como otras proteínas de la familia de las SNARE (Wong y col., 2007), aunque el mecanismo es mucho más complejo (López- Godínez y col., 2003). La función de los gránulos corticales cobra entonces especial relevancia en medicina, ya que alteraciones en su ubicación, composición y función es causa de infertilidad, incluso en la mujer.
Aunado a la exocitosis de los gránulos corticales, la activación del óvulo se caracteriza por la reorganización del citoesqueleto de actina de su región cortical (Roux y col., 2008) y por un significativo aumento del pHi (Molich y col., 2005). Este último es fundamental para iniciar la traducción del RNA mensajero acumulado durante la oogénesis (Stitzel y col., 2007), aunque no se sabe cómo ocurre esto. Se ha postulado que el aumento en el pHi es debido a la actividad transportadora de un intercambiador Na+/H+^ (NHE). En células epiteliales de mamíferos, se han descrito al menos 9 isoformas del NHE (NHE1- NHE9) (Kiela y col., 2006). Estos intercambiadores se relacionan con cambios estructura les a nivel de citoesqueleto de actina, interactuando con las proteínas ezrin/radixin/moesin (ERM). La fosforilación de estas proteínas por ROCK cinasa (cinasa de la vía de
FEBRERO DEL 2010
No 04- 2010 - 101813383300 - 102 ®
Ridley, 2003). Ambas isoformas son activadas por Rho y segundos mensajeros tales como ácido araquidónico y esfingosilfosforilcolina. Las proteínas ROCK fosforilan diversos sustratos proteicos como los que regulan el ensamble de los filamentos de actina, la fosfatidilinositol-3 cinasa, la cinasa LIMK que fosforila cofilina, la cadena ligera de la miosina (CLM) (Uehara y col., 2008), la fosfatasa de la cadena ligera de la miosina (CLMP), las proteínas NHE y canales de Na+^ y K+^ e inhiben la actividad de canales de Ca2+ tipo T (Iftinca, 2007). Todos estos sustratos proteicos presentan secuencias conse nso R/KXS/T o R/KXXS/T (R, arginina; K, lisina; S, serina; T, treonina; X, cualquier aminoácido) en el sitio de fosforilación.
Se han sintetizado dos compuestos muy selectivos para bloquear competitivamente la actividad catalítica de ROCK: Y-27632 y H-1152 (Shum y col., 2003). Estos son permeables y tienen una Ki de 140 nM y 1.6 nM, respectivamente (Riento y Ridley, 2003). Nuestros resultados con estos inhibidores muestran que Y-27632 inhibe la alcalinización intracelular que se dispara minutos después de la fecundación (Rangel-Mata y col., 2007); hemos observado también que el H-1152 bloquea la exocitosis del acrosoma en el espermatozoide, proceso que es crucial para la fecundación (De la Sancha y col., 2007). Si bien los estudios con estos inhibidores permiten inferir la presencia de ROCK en los gametos del erizo de mar, esto se podrá confirmar con la identificación de una región codificante en el genoma de erizo de mar y su expresión en el óvulo y cigoto. Por otra parte, la identificación y localización de sustratos de ROCK permitirán establecer un mecanismo molecular de la vía de transducción de señales Rho/ROCK en eventos cruciales de la fecundación.
REFERENCIAS
Alves, A.P., Mulloy, B., Diniz J.A. and Mourao, P.A. (1997). Sulfated polysaccharides from the egg jelly layer are species-specific inducers of acrosomal reaction in sperms of sea urchins. J. Biol. Chem. 272: 6965-6971.
Beltrán, C., Galindo, B.E., Rodríguez-Miranda, E. and Sánchez, D. (2007a). Signal transduction mechanisms regulating ion fluxes in the sea urchin sperm. Signal Transduction. 7: 103-117.
FEBRERO DEL 2010
No 04- 2010 - 101813383300 - 102 ®
Beltrán, C., Vacquier, V.D., Moy, G., Chen, Y., Buck, J., Levin, L.R. and Darszon, A. (2007b). Particulate and soluble adenylyl cyclases participate in the sperm acrosome reaction. Biochem Biophys Res Commun. 13;358(4):1128-35.
Carrol, E.J. (1976). Cortical granule proteases from sea urchin eggs. Methods Enzymol. 54:343-
Christen, R., Schackmann, R.W. and Shapiro, B.M. (1982). Elevation of the intracellular pH activates respiration and motility of sperm of the sea urchin, Strongylocentrotus purpuratus. J. Biol. Chem. 257:14881-14890.
Covian-Nares, F., Martínez-Cadena, G., Lopez-Godínez, J., Voronina, E., Wessel, G.M. and Garcia-Soto, J. (2004). A Rho-signaling pathway mediates cortical granule translocation in the sea urchin oocyte. Mech. Dev. 121 (3): 225-35.
Cuéllar-Mata, P., Martínez-Cadena, G., López-Godínez, J., Obregón, A. and García-Soto, J. (2000). The GTP-binding protein RhoA localizes to the cortical granules of Strongylocentrotus purpuratus sea urchin egg and is secreted during fertilization. Eur. J. Cell Biol. 79:81-91.
De la Sancha, C.U., Martínez-Cadena, G., López-Godínez, J., Castellano, L.E., Nishigaki, T., Darszon, A. and García-Soto, J. (2007). Rho-kinase (ROCK) in sea urchin sperm: its role in regulating the intracellular pH during the acrosome reaction. Biochem. Biophys. Res. Commun. 364 (3): 470-5.
Darszon, A., Nishigaki, T., Wood, C., Trevino, C.L., Felix, R. and Beltran, C. (2005). Calcium channels and Ca2+^ fluctuations in sperm physiology. Int. Rev. Cytol. 243:79-172.
Darszon, A., Guerrero, A., Galindo, B.E., Nishigaki, T., and Wood, C.D. (2008). Sperm- activating peptides in the regulation of ion fluxes, signal transduction and motility. Nt. J. Dev. Biol. 52: 595 – 606.
Darszon, A., Acevedo, J.J., Galindo, B.E., Hernández-González, E.O., Nishigaki, T., Treviño, C.L., Wood, C. and Beltrán, C. (2006). Sperm channel diversity and functional multiplicity. Reproduction. 131(6):977-88.
Hirohashi, N., and Vacquier, V.D. (2002). Egg fucose sulfate polymer, sialoglycan and speract all trigger the sea urchin sperm acrosome reaction. Biochem. Biophys. Res. Commun.. 296: 833-839.
FEBRERO DEL 2010
No 04- 2010 - 101813383300 - 102 ®
Riento, K. and Ridley, A.J. (2003). Rocks: multifunctional kinases in cell behaviour. Nat. Rev. Mol. Cell Biol. 4(6): 446-56.
Roux, M.M., Radeke, M.J., Goel, M., Mushegian, A. and Foltz, K.R. (2008). 2DE identification of proteins exhibiting turnover and phosphorylation dynamics during sea urchin egg activation. Dev. Biol. 313(2): 630-47.
Sardet, D. and Tilney, L. (1970). Origin of acrosomal precess is actin complexed with membrane precursos? Cell Biol. Rep. 1:193-200.
SeGall, G.K. and Lennarz, W.J. (1981). Jelly coat and induction of the acrosome reaction in echinoid sperm. Dev. Biol. 86(1): 87-93.
Schmandke, A. and Strittmatter, S.M. (2007). ROCK and Rho: biochemistry and neuronal functions of Rho-associated protein kinases. Neuroscientist. 13(5): 454-69.
Shum, W.W., Le, G.Y., Jones, R.L., Gurney, A.M. and Sasaki ,Y. (2003). Involvement of Rho- kinase in contraction of guinea-pig aorta induced by prostanoid EP3 receptor agonists. Br. J. Pharmacol. 139(8): 1449-61.
Sodergren, E., G. M. Weinstock, et al. (2006). The genome of the sea urchin Strongylocentrotus purpuratus. Science 314(5801): 941-52.
Stitzel, M. L. and Seydoux, G. (2007). Regulation of the oocyte-to-zygote transition. Science 316(5823): 407-8.
Townley, I.K., Roux, M.M. and Foltz, K.R. ( 2006). Signal transduction at fertilization: the Ca2+ release pathway in echinoderms and other invertebrate deuterostomes. Semin. Cell Dev. Biol. 17(2): 293-302.
Trimmer, J.S., Trowbridge, I.S. and Vacquier, V.D. (1985). Monoclonal antibody to a membrane glycoprotein inhibits the acrosome reaction and associated Ca2+^ and H+^ fluxes of sea urchin sperm. Cell. 40: 697–703.
Uehara, R., Hosoya, H. and Mabuchi, I. (2008). In vivo phosphorylation of regulatory light chain of myosin II in sea urchin eggs and its role in controlling myosin localization and function during cytokinesis. Cell Motil. Cytosk. 65(2): 100-15.
FEBRERO DEL 2010
No 04- 2010 - 101813383300 - 102 ®
Vacquier, V.D., Swanson, W.J. and Hellberg, M.E. (1995). What have we learned about sea urchin sperm bindin? Dev. Growth Different. 37: 1–10.
Vacquier, V.D. and Moy, G.W. (1997). The fucose sulfate polymer of egg jelly binds to sperm REJ and is the inducer of the sea urchin sperm acrosome reaction. Dev. Biol. 192(1): 125-135.
Vilela-Silva, A.C., M.O. Castro, A.P. Valente, C.H. Biermann, and P.A. Mourao. (2002). Sulfated fucans from the egg jellies of the closely related sea urchins Strongylocentrotus droebachiensis and Strongylocentrotus pallidus ensure species-specific fertilization. J. Biol. Chem. 277:379-87.
Wong, J.L., Koppel, D.E., Cowan, A.E. and Wessel, G.M. (2007). Membrane hemifusion is a stable intermediate of exocytosis. Dev. Cell. 12(4): 653-9.