¡Descarga resum y más Resúmenes en PDF de Psicobiología solo en Docsity!
MÒDUL 1 – LA PSICOBIOLOGIA Noemi Ferré Fdez. | 2012/
1. Què és la psicobiologia i quin mètode d’estudi utilitza?
La Psicobiologia és la disciplina que pretén estudiar la conducta i la cognició però atenent a les bases biològiques subjacents. Utilitzant el mètode científic, la Psicobiologia s’interessa en analitzar els diferents components del sistema neuroendocrí implicats, els factors genètics i epigenètics subjacents, els processos que posen en marxa la conducta i aquells que la controlen. Per tant, estudiar el paper del SN, en interacció amb la resta del cos, sobre el control del comportament.
2. Què és la cirurgia estereotàxica i què permet estudiar?
La cirurgia estereotàxica és un procediment quirúrgic que permet accedir a estructures profundes del cervell amb l’objectiu d’implantar dispositius que en permetran l’estudi. Així, es poden implantar elèctrodes, cànules, obtenir mostres de teixit, fer lesions o injectar fàrmacs al cervell. Es pot usar amb finalitats experimentals amb animals de laboratori o amb finalitat clínica en certes intervencions quirúrgiques que es practiquen en humans (biòpsies, implantació d’elèctrodes d’estimulació cerebral profunda, etc.).
3. En què consisteixen les tècniques de registre de l’activitat neuronal?
Les tècniques de registre de l’activitat neuronal permeten registrar de manera directa l’activitat, tant elèctrica com química, de les neurones. Amb aquestes tècniques es pot mesurar l’activitat d’una sola neurona i també la de moltes neurones alhora, de manera que es pot registrar l’activitat d’una regió cerebral. És a dir, amb aquestes tècniques es poden mesurar els potencials d’acció, els potencials locals, etc. i, per tant, permeten conèixer si una estructura del SN està activa o no en un determinat moment.
4. En què consisteixen les tècniques d’estimulació de l’activitat neuronal?
Les tècniques d’estimulació de l’activitat neuronal són aquelles que ens permeten estimular àrees concretes del SN amb l’objectiu d’esbrinar en quines funcions participa l’àrea estimulada i observar quins canvis conductuals es produeixen en el subjecte després de l’estimulació. L’estimulació pot ser elèctrica , mitjançant elèctrodes implantats en àrees concretes del cervell pels quals es fa passar un corrent elèctric de baixa intensitat, o química injectant determinades substàncies en àrees concretes del cervell amb una cànula.
5. En què consisteixen les tècniques de registre psicofisiològic? ()* - L’electroencefalograma (EEG): permet registrar l’activitat elèctrica cerebral d’una manera global a temps real, utilitzant elèctrodes situats en diferents zones del cuir cabellut. Recullen l’activitat i una màquina tradueix els senyals en ones. - L’electromiograma (EMG) i l’electrooculograma (EOG): registren l’activitat del SN somàtic. L’ EMG recull info sobre el grau de tensió muscular amb elèctrodes situats a la superfície del grup muscular que interessa. L’ EOG registra el moviment dels ulls, en situar els elèctrodes als músculs que els envolten. - L’electrocardiograma (ECG): com a mesurament del SN autònom, se sol registrar l’activitat del sistema cardiovascular, mesurant la freqüència cardíaca amb el ECG. Aquest registra l’activitat cardíaca mitjançant elèctrodes en zones del pit, i la tensió arterial mitjançant esfigmomanòmetre. 6. Explica en què consisteixen les tècniques d’estudi del SN mitjançant lesions cerebrals.
Són aquelles tècniques que ens permeten observar la conducta d’un subjecte que ha sofert una lesió a una àrea concreta del cervell i veure en quins processos està implicada aquesta àrea: La major part estudis de lesions s’elaboren induint lesions a animals d’experimentació però en el cas dels humans, s’utilitzen voluntaris que han sofert algun tipus de lesió cerebral, com per exemple, traumatismes cranioencefàlics o accidents vasculars.
7. Quin coneixement pot aportar l’estudi de lesions cerebrals? Com es realitzen i en quin tipus de subjectes?
Amb aquesta tècnica podem estudiar les relacions entre les regions cerebrals afectades i els dèficit conductuals mostrats pel subjecte. Aquesta tècnica es pot fer servir en animals experimentals, a qui se’ls causarà lesions controlades de regions cerebrals concretes per posteriorment avaluar la seva conducta. En humans es poden avaluar els dèficit cognitius, conductuals i emocionals mostrats per pacients amb lesions adquirides a diferents regions cerebrals.
8. Explica breument en què consisteixen les tècniques d’investigació farmacològiques.
Les tècniques farmacològiques permeten estudiar l’efecte de diverses substàncies sobre el sistema nerviós. Consisteixen a administrar una substància al subjecte experimental i observar com aquesta influeix en la seva conducta. Les substàncies poden ser fàrmacs, com un antidepressiu, o drogues, com la cocaïna.
9. Explica en què consisteixen les tècniques d’estudi del SN post mortem****.
Les tècniques post mortem són aquelles que requereixen el sacrifici dels subjectes experimentals, en cas que es tracti d’animals d’experimentació, o l’obtenció de mostres de teixit humà de persones que hagin donat els seus cossos a la ciència. Aquestes tècniques permeten estudiar amb més detall l’anatomia del sistema nerviós i identificar components que són impossibles d’estudiar in vivo. Algunes: tècniques histològiques, inmunocitoquímiques, autradiografia, hibiridació in situ.
10. En què consisteixen les tècniques genètiques? ()*
Estudiar la participació dels gens en la conducta, si algun tret té base genètica, quin tipus d’herència segueix o identificar els gens implicats en l’expressió d’un tret o malaltia psíquica. En humans es fan estudis de famílies, adopcions o bessons. És possible analitzar el genoma dels subjectes per buscar-hi alteracions gèniques. En animals es basen en el control dels aparellaments o s’utilitzen tècniques que permeten manipular directament el genoma.
11. Què són les tècniques de neuroimatge?
Les tècniques de neuroimatge constitueixen mètodes de visualització del cervell humà viu. Els avenços en les tècniques de rajos X i informàtiques han permès desenvolupar diferents mètodes per al estudi de la estructura i funcions del Cervell humà en viu. Alguns exemples d’aquestes tècniques són: o Ressonància magnètica (RM ): No utilitza rajos X, sinó ones electromagnètiques i permet obtenir imatges de l’interior del nostre organisme. o Tomografia axial computada (TAC): utilitza rajos X per a visualitzar la estructura del cervell (o d’altres estructures internes del cos humà) o Tomografía por emisión de positrones (TEP) : Aquesta tècnica de neuroimatge permet als investigadors determinar el nivell d’activitat metabòlica en diferents zones del cervell, i la presència de patologies a partir del subministrament d’un radiofàrmac. o Electroencefalografia (EEG) : permet registrar l’activitat elèctrica cerebral que subjeu a diferents processos cognitius, motors i sensorials. o Magnetoencefalografia (MEG) : permet registrar l’activitat cerebral. També s’utilitza per avaluar o estudiar processos cognitius bàsics com la memòria, el llenguatge, la percepció o funcions exectuives. o Estimulació magnètica transcranial (EMT) : No registra activitat cerebral sinó que indueix un corrent al cervell mitjançant un camp magnètica per a generar un benefici terapèutic o establir relacions causals entre activitat cerebral i comportament. o Estimulació elèctrica cortical (EEC) : Utilitzada en neurocirurgia, fent un mapatge cortical i veure quines àrees cerebrals poden quedar afectades.
17. Què és la estimulació magnètica transcranial (EMT)? ()*
No registra l’activitat cerebral sinó que consisteix a induir un corrent al cervell mitjançant un camp magnètic amb l’objectiu de generar un benefici terapèutic i/o establir relacions causals entre activitat cerebral i comportament.
S’utilitza per a efectuar mapatges corticals de diferents funcions cerebrals com la memòria, el llenguatge, etc. i fins i tot per a establir relacions causals entre excitabilitat i comportament.
18. Què és la estimulació elèctrica cortical (EEC)? ()*
Utilitzada bàsicament en neurocirurgia, fent un mapatge cortical i veure quines àrees cerebrals poden quedar afectades durant l’operació.
19. Imagina’t que treballes en un grup de recerca i vols portar a terme un treball en el qual vols estudiar quines estructures del sistema nerviós central estan implicades en el procés de codificació de la informació durant un aprenentatge. Quina tècnica d’estudi no invasiva creus que seria més adequada utilitzar? Per què?
La Ressonància Magnètica Funcional és una tècnica de neuroimatge no invasiva que permet registrar l’activitat cerebral in vivo en temps real. Això permetria veure quines estructures cerebrals s’activen mentre el subjecte codifica la informació mentre realitza una tasca d’aprenentatge.
MÒDUL 2 – LES CÈL·LULES DEL SN
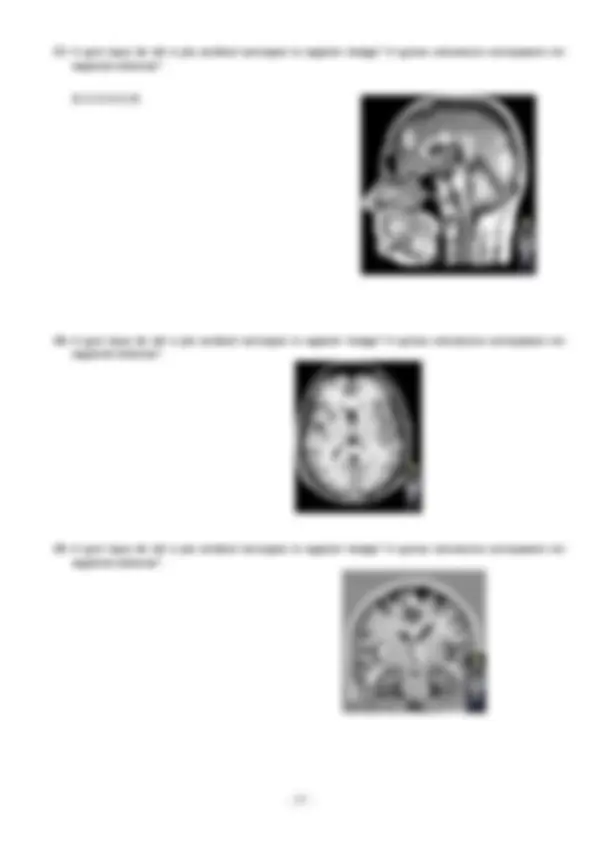
1. En la següent imatge, quin tipus cel·lular estan indicant les lletres A, B, C i D? Quines regions de la mateixa cèl·lula assenyalen els números 1, 2, 3, 4 i 5? Creus que es tracta d’un dibuix del sistema nerviós central o del sistema nerviós perifèric? Per què? Màxim 3 línies Quin tipus de conducció del potencial d’acció es produirà en la cèl·lula A? Per què?
A: Neurona multipolar, del tipus: Golgi I. B: Oligodendròcit C: Micròglia D: Astròcid
- Dendrites
- Soma (cos cel·lular)
- Nucli
- Axó / Nòdul de Ranvier
- Beina de Mielina (Internòdul)
Es tracta del Sistema nerviós central (SNC) ja que estan representades les cèl·lules glia com l’oligodendròcit són únicament presents en aquest sistema. A més es tracta d’una neurona motora, i aquestes són fibres eferents del SNC que transporten informació al SNP. Conducció Saltatòria. Perquè l’axó d’aquesta està milienitzat i només podrà haver despolarització quan els corrents de ions arribin als Nòduls de Ranvier, i per tant no podrà ser continua durant tota la membrana.
2. Descriu breument la classificació de les neurones segons la seva morfologia. Descriu les seves principals característiques.
Segons la seva la morfologia podem distingir entre neurones unipolars, bipolars i multipolars.
Les neurones unipolars són les més simples. Del soma surt una sola prolongació que es pot ramificar en moltes branques. Una d’aquestes serveix d’axó, i les altres funcionen com a estructures dendrítiques de recepció. En mamífers, les neurones unipolars són un tipus especial de cèl·lules unipolars anomenades neurones pseudomonopolars, pseudounipolars o neurones en T. En les neurones bipolars surten dues prolongacions del cos cel·lular. Es troben principalment en els sistemes sensorials, com és el cas de les cèl·lules bipolars de la retina. Les neurones multipolars són el tipus de neurones més comú en l’SN dels vertebrats. Del soma surt l’axó i diverses ramificacions dendrítiques. Segons la longitud de l’axó, les podem dividir en multipolars, tipus Golgi I (neurones d’axó llarg) i tipus Golgi II (neurones d’axó curt).
3. Classifica les neurones segons la seva funció. Descriu les seves principals característiques.
Segons la seva la funció podem distingir entre neurones sensorials, motores i interneurones. Les neurones sensorials porten informació des de la perifèria fins a l’SNC, per tant, són fibres aferents a l’SNC
. Generalment són neurones pseudomonopolars. Les neurones motores porten informació des de l’SNC fins a la perifèria (músculs i glàndules), per tant, són fibres eferents de l’SNC. Generalment són neurones multipolars Golgi I. Les interneurones són el tipus de neurones més abundant; són totes les altres neurones que no són ni sensorials ni motores. Aquestes neurones processen informació localment i la transmeten d’un lloc a l’altre de l’SNC.
8. Quines són les cèl·lules que formen la beina de mielina a l’SNP i a l’SNC? Quina és la funció principal de la mielina? Què són els nodes de Ranvier? I els internodes? On es concentren més canals iònics, als nodes o als internodes? De quin tipus de canals iònics estaríem parlant? A quina part de la neurona es dóna la major concentració d’aquests canals iònics? Per què? Què hi tenen a veure els potencials locals amb aquesta part de la neurona rica en canals iònics?
Al SNP les cèl·lules de Schwann formen la beina de mielina mentre que al SNC els oligodendròcits són els que la formen. La mielina aïlla l’axó i aquest aïllament produeix l’acceleració de la propagació de l’impuls nerviós. Els nodes de Ranvier són el segment de l’axó on la membrana no està coberta per mielina mentre que els internodes són els segments de l’axó coberts per la mielina. Als nodes es concentren més canals iònics de sodi i de potassi (voltatge dependents) ja que quan es genera un potencial d’acció al conus axònic d’un axó mielinitzat, el senyal es transmet de forma passiva al llarg del primer segment de mielina fins al primer node de Ranvier. El senyal arriba al primer node, i és el suficientment intens com per obrir els canals de sodi activats per voltatge del node i generar un altre potencial d’acció. Aquest potencial d’acció es transmet novament al següent node de forma passiva i així successivament. Aquest fenomen es coneix com a conducció saltatòria. Al conus axònic es concentren la major part d’aquets canals iònics. És on s’integren els senyals elèctrics per produir el potencial d’acció. Que una neurona dispari o no un potencial d’acció depèn de l’equilibri entre els senyals excitadors i els senyals inhibidors que arriben al conus axònic. Els PEPs i PIPs (potencials locals), productes de l’acció dels neurotransmissors sobre els receptors postsinàptics, són conduïts i es van atenuant progressivament fins arribar al conus axònic. Si la suma de despolaritzacions i hiperpolaritzacions arriba al llindar de descàrrega es generarà un potencial d’acció.
9. En relació amb la pregunta anterior, és el mateix tipus de canals iònics el que intervé en el potencial de repòs? Per què es produeix el potencial de repòs? Quin és el principal ió responsable del valor del potencial de repòs? Quin paper juga la bomba de Na+/K+ en el manteniment del potencial de repòs? La bomba de Na+/K+, és un canal iònic?
Al potencial d’acció intervenen els canals dependents de voltatge. Al potencial de repòs els canals passius hi juguen un paper més important. L'existència de diferents quantitats d'aquests canals per a cada ió fa que la membrana sigui més o menys permeable a uns ions que a uns altres i això genera una distribució asimètrica dels ions que implica una diferència de potencial elèctric entre el dos costats de la membrana (potencial de membrana). El principal responsable del potencial de repòs és el K+. La bomba de Na+/K+ està integrada per molècules proteiques inserides a la membrana i impulsades per l’energia que proporcionen les molècules d’ATP produïdes pels mitocondris. La bomba extreu tres ions de Na+ cap a l’exterior per cada dos ions de K+ que introdueix a l’interior. Així restableix el potencial de repòs de la membrana després del potencial d’acció. Tot i que la bomba permet la circulació d’ions entre les dos costats de la membrana neuronal, no es considera pròpiament un canal iònic ja que els ions travessen la membrana en contra del seu gradient electroquímic, de manera dependent d’energia (ATP).
10. Explica la membrana de la neurona. ()*
És l’estructura que defineix els límits de la neurona; separa el fluid intracel·lular del extracel·lular. És una doble capa de molècules lipídiques en que floten diferents tipus de molècules proteiques. Entre aquestes molècules s’hi troben els canals que controlen l’accés. Aquests canals s’anomenen canals iònics i són proteïnes en forma de canal. N’hi ha de dos tipus:
- Passiu � sempre estan oberts
- Actius � s’obren de manera transitòria i poden ser dependents de lligands/molècules, o dependents de voltatge (varia en resposta als canvis en el potencial de membrana).
11. Què és el potencial de membrana i perquè es dóna? ()*
El potencial de membrana fa referència a la diferència de càrrega elèctrica entre l’interior i l’exterior cel·lular. Aquesta diferència és causada perquè hi ha molècules (ions) amb diferents càrregues (postives = cations / negatives = anions) i es troben en quantitats diferents a l’interior i a l’exterior de la cèl·lula. En aquesta diferència hi intervenen dos forces oposades:
- Força de difusió (química) � Referent al moviment de les molècules al desplaçar-se de regions on es troben en concentracions altes a regions de concentració baixa.
- Força electroestàtica � Referent a l’atracció o repulsió de les partícules entre elles en funció de la càrrega elèctrica (Ions amb càrregues oposades s’atreuen, i amb càrregues iguals es refuten). Quan els ions estiguin separats per una membrana permeable la travessaran per assolir l’equilibri. Quan la membrana sigui semipermeable, els ions que la travessin es disposaran de manera asimètrica per compensar els que no, creant la diferència de potencial elèctric i químic = Potencial de membrana. 12. Què és el potencial de repòs i per què es produeix?
El potencial de membrana fa referència a la diferència de càrrega elèctrica que es produeix entre l’interior i l’exterior cel·lular. Quan les neurones no estan actives, és a dir, no reben ni condueixen informació, aquest potencial de membrana se l’anomena potencial de repòs. El potencial de repòs té un valor d’entre –60 i – mV (l’interior cel·lular és negatiu respecte a l’exterior cel·lular, ja que conté un nombre més gran de càrregues negatives). El potencial de repòs es produeix per la naturalesa semipermeable de la membrana i per l’existència de la bomba Na+-K+. En condicions de repòs, la membrana és molt permeable al K+, és a dir, la membrana té molts canals passius per a aquest ió. Per tant, el K+^ pot travessar la membrana a favor del seu gradient electroquímic (segons les forces químiques i les elèctriques) i distribuir-se asimètricament contrarestant els ions que no la poden travessar o que la travessen amb menys facilitat. Per altra banda, la membrana no és totalment impermeable al Na+. Encara que la permeabilitat de la membrana al Na+^ és baixa mentre la neurona està en repòs, hi ha una filtració de Na+^ cap a l’interior cel·lular. Aquesta situació, que augmenta la positivitat del fluid intracel·lular, contribueix al fet que el K+^ tingui tendència a sortir. En teoria, la diferència de concentració entre els dos ions tendiria a desaparèixer si no existís la bomba de Na+-K+^ que expulsa ions de Na+^ cap a l’exterior i impulsa ions de K+^ a l’interior. La bomba de Na+-K+^ és un sistema de transport actiu que manté estable el valor del potencial de repòs. La bomba de Na+-K+^ expulsa tres ions de Na+^ a l’exterior per cada dos de K+^ que entra a l’interior. Afavorint així la negativitat interior de la cèl·lula i mantenint el valor del potencial de repòs.
13. En què consisteix el potencial de repòs? Com es distribueixen els ions en aquestes condicions?
El potencial de repòs és la diferència de voltatge que s’observa entre els dos costats de la membrana de les neurones quan aquestes no estan actives (no reben ni transmeten cap mena d’estimulació). En aquesta condició, a l’interior de la neurona se situen els anions orgànics (A-), de càrrega negativa, i el K+ (potassi), de càrrega positiva, mentre que a l’exterior trobem el Na+^ (Sodi / positiu) i el Cl- (clor / negatiu). En aquesta condició de repòs la membrana és molt més permeable al K+^ que al Na+, sent la permeabilitat al Cl- intermèdia entre aquests valors. La membrana és totalment impermeable als anions orgànics (A-). Això fa que l’interior de la neurona sigui negatiu respecte l’exterior (uns -70mV), i que el K+, bàsicament pel seu gradient de concentració (químic), es mogui de l’interior a l’exterior de la membrana buscant el seu potencial d’equilibri, que és d’uns -90mV. De tota manera, no aconsegueix mai aquest valor ja que hi ha una petita entrada contínua de Na+^ (que té un gran gradient electroquímic per entrar a la neurona) que compensa la sortida de contínua de K+. A més, la bomba de Na+/K+^ s’encarrega de mantenir aquesta distribució d’ions entre els dos costats de la membrana durant el potencial de repòs.
17. Quines són les principals diferències i similituds entre els potencials locals i els potencials d’acció?
Els potencials locals consisteixen en canvis en el potencial de membrana que es produeixen en un punt de la membrana neuronal deguts a l’arribada d’un estímul dèbil. Els potencials locals es propaguen, de manera passiva, al llarg de la membrana neuronal amb pèrdua d’intensitat a mesura que s’allunyen del punt d’estimulació (conducció electrotònica). Els potencials locals poden consistir en petites despolaritzacions o bé petites hiperpolaritzacions en el potencial de membrana.
Els potencials d’acció són també canvis en el potencial de membrana, però a diferència dels potencials locals, aquests canvis són més intensos. Una altra diferència consisteix en què el potencials locals es propaguen de manera activa al llarg de la membrana axonal, és a dir, es condueixen sense pèrdua d’intensitat, regenerant- se al llarg de l’axó. Cada potencial d’acció, a diferència dels potencials locals, consta de tres fases: despolarització, repolarització i hiperpolarització.
18. Defineix què és un potencial d’acció i explica les seves fases i els seus fonaments iònics.
El potencial d’acció (PA) és el senyal elèctric bàsic que representa la transmissió d’informació en el sistema nerviós. El PA consta de tres fases:
El potencial d'acció comença amb la fase despolarització en la que el potencial de membrana s'inverteix fins a arribar a un valor proper al valor del potencial d'equilibri del sodi (aproximadament al voltant dels +50 mV). Aquesta fase es deu a l'obertura dels canals de sodi controlats per voltatge i la consegüent entrada de sodi que positivitza l'interior cel·lular. El tancament d'aquests canals unit a l'obertura dels canals de potassi controlats per voltatge, fa que s'aturi el flux de sodi i es generi un important flux de sortida d'ions de potassi. La conseqüència d'aquest canvi és que l'interior recupera la seva negativitat (fase de repolarització ). Aquest augment de la negativitat interna sobrepassa el valor del potencial de repòs (fase d'hiperpolarització) degut a què la permeabilitat per al potassi és major que en condicions de repòs. Quan els canals actius de potassi es tanquin i la permeabilitat torni als valors de repòs, s'assolirà novament el valor del potencial de repòs.
El potencial d’acció és un canvi ràpid en el potencial de membrana que es condueix de manera activa, sense pèrdua d’intensitat, i que es regenera al llarg de l’axó.
Fases del potencial d’acció:
- Fase de despolarització. L’interior perd negativitat fins que arriba un moment en què no hi ha diferència de potencial. De fet, fins i tot el potencial s’inverteix, l’interior es fa positiu respecte a l’exterior.
- Fase de repolarització. L’interior recupera la seva negativitat respecte a l’exterior.
- Fase d’hiperpolarització , en la qual l’interior presenta un valor més negatiu que el del potencial de repòs.
- Per acabar, el potencial de membrana recupera el valor del potencial de repòs.
Fonaments iònics:
- Fase de despolarització : es correspon amb un canvi (augment) en la permeabilitat de la membrana al Na+, per a l’obertura de canals de Na+^ dependents de voltatge. Aquesta obertura fa que el Na+^ es precipiti cap a l’interior de la cèl·lula i despolaritzi la membrana
- Fase de repolarització : es produeix quan quan augmenta la conductància del K+, i disminueix la del Na+. La sortida d’ions de K+^ produeix la repolarització de la membrana, la qual va recuperant la seva negativitat interna
- Fase d’hiperpolarització : quan la membrana recupera el valor del potencial de repòs, la conductància del K+^ continua essent elevada, i això fa que el K+^ continuï sortint de la neurona i fent que la membrana s’hiperpolaritzi.
19. Què és el llindar de descàrrega? Quina relació té amb la intensitat de l’estímul? En què es diferencia la codificació neuronal d’un estímul que arriba al llindar de descàrrega i la d’un altre estímul que supera aquest llindar?
El llindar de descàrrega és el valor del potencial de membrana a partir del qual s'obren els canals de sodi controlats per voltatge i per tant es dispara el potencial d'acció. Aquest valor suposa una despolarització respecte al potencial de repòs. Estímuls de baixa intensitat produiran una despolarització dèbil (proporcional a la intensitat de l'estímul) que no arribarà al llindar de descàrrega i, per tant, no generaran un potencial d'acció sinó un potencial local, només aquells estímuls que tinguin una intensitat suficient per fer arribar el valor del potencial de membrana fins al llindar de descàrrega generaran un potencial d'acció. Estímuls amb intensitat encara més gran no generaran potencials d'acció de més amplitud, sinó major freqüència de potencials d'acció.
20. En què consisteix la fase de despolarització del potencial d’acció? Explica les bases iòniques.
La despolarización se corresponde con la primera fase del potencial de acción y se caracteriza porque el interior celular se vuelve positivo respeto al exterior. Para que se de esta fase antes que nada necesitamos que se dé una estimulación intensa que produzca una despolarización de la membrana (aproximadamente 15 mV). A continuación, se abren los canales de Na+ dependientes de voltaje: el Na+ se precipita hacia el interior impulsado tanto por fuerzas químicas como eléctricas, y produce una despolarización rápida del potencial de membrana. Los canales de Na+ dependientes de voltaje se abren muy rápidamente y se mantienen abiertos durante un periodo corto de tiempo. Por lo tanto, la conductancia del Na+ aumenta muy rápidamente, pero también disminuye muy rápidamente.
El potencial d’acció consta de tres fases: despolarització, repolarització i hiperpolarització. La fase de despolarització es caracteritza pel fet que l’interior de la membrana perd negativitat fins que arriba un moment en què no hi ha diferència de potencial i, posteriorment, el potencial s’inverteix, és a dir, l’interior es fa positiu respecte a l’exterior. El potencial d’acció es desencadena quan s’arriba al llindar de descàrrega, és a dir, a un determinat valor de despolarització. En aquest moment, s’obren els canals de Na+ dependents de voltatge: el Na+ es precipita cap a l’interior impulsat tant per forces químiques com elèctriques, i produeix una despolarització ràpida del potencial de membrana. Els ions de Na+ tendeixen a entrar fins que aconsegueixen que el potencial de membrana coincideixi amb el valor del seu potencial d’equilibri. El valor del potencial d’equilibri del Na+ és de +55 mV, però el potencial d’acció té un pic (punt de més gran despolarització) de +40 mV. Per tant, el Na+ mai no arriba a estar en equilibri, ja que la conductància del K+ comença a augmentar gairebé al mateix temps que comença a disminuir la conductància del Na+.
21. Descriu la fase de repolarització del potencial d’acció i els seus fonaments iònics.
La repolarización del potencial de acción se produce por la apertura de los canales de K+ dependientes de voltaje. Esta apertura hace que el K+ salga de la neurona y repolarice el potencial de membrana. Cuando aumenta la conductancia del K+ y disminuye la del Na+, la salida de iones de K+ produce la repolarización de la membrana, la cual va recuperando su negatividad interna. Los iones de K+ salen impulsados por fuerzas eléctricas y químicas. Fuerzas químicas : La concentración de K+ es mayor en el interior y, por lo tanto, estas fuerzas lo empujan a salir. Fuerzas eléctricas : La entrada masiva de Na+ ha hecho que el interior adquiera valores positivos. Como el K+ es un catión, las fuerzas eléctricas también lo empujan a salir.
26. Què són els períodes refractaris? Quins tipus hi ha i en què consisteixen?
És el temps que ha de transcórrer després d’un potencial d’acció perquè un estímul llindar sigui capaç de produir un nou potencial d’acció en el mateix punt. Els períodes refractaris es deuen a que els canals de sodi dependents de voltatge es mantenen inactius durant un cert període de temps. Hi ha dos tipus de períodes refractaris:
- Període refractari absolut: és la part del període refractari en la qual per molt alta que sigui la intensitat de l’estímul no es produeix un potencial d’acció.
- Període refractari relatiu: és la part del període refractari en la qual, si l’estimulació és prou intensa (per sobre l’estímul llindar), s’aconsegueix de produir un nou potencial d’acció; per tant, és un període en el qual el llindar de descàrrega és més elevat. 27. Què passaria si les neurones no tinguessin període refractari? Creus que el potencial d’acció podria propagar-se pel soma i les dendrites? I creus que seria possible que s’alliberés neurotransmissor des de les dendrites? Per què?
Si les neurones no tinguessin període refractari, el potencial d’acció que es propaga per l’axó en direcció als botons terminals, podria també propagar-se en direcció al soma de la neurona. Els canals de Na+ i K+ responsables del potencial d’acció, que són dependents de voltatge, s’estarien obrint constantment, independentment de la història recent d’activacions (és a dir, independentment de si el potencial d’acció acaba de passar pel punt de la membrana on estan situats), pel que el potencial d’acció no es podria propagar en una sola direcció. Per que les dendrites i/o el soma alliberessin neurotransmissors es necessitaria que aquestes estructures tinguessin canals de Ca2+ dependents de voltatge, iguals als que trobem als botons terminals. En qualsevol cas, hi ha alguns tipus de neurotransmissors, com els gasos (l’òxid nítric, per exemple), que poden travessar lliurament la membrana cel·lular, i que per tant poden ser alliberats per les dendrites i els somes de les neurones postsinàptiques en direcció a les presinàptiques, on actuen sobre receptors situats als botons terminals i poden modular l’alliberació de neurotransmissors en el “sentit” clàssic (de neurona presinàptica a postsinàptica).
28. Explica la conducció de la informació. Explica la conducció en fibres amielíniques i mielíniques.
L’axó condueix la informació des del extrem més proper al cos cel·lular (segment inicial /con axònic) fins a l’extrem dels botons terminals ( Direcció ortrodròmica ), encara que també pot ser al revés ( Direcció antidròmica ). [Si apliquem l’estímul al mig va en totes dues direccions]. La velocitat de conducció és més alta si l’axó és més ampli, és a dir si té més diàmetre.
Conducció en fibres amielíniques : Es dóna l’estímul llindar en un punt, que desencadena el potencial d’acció, entrant els ions Na+^ al interior. Aquests ions positius creen corrents d’ions que són atrets per les zones negatives adjacents de l’interior (les càrregues positives són atretes per les negatives veïnes). Aquest moviment produirà la despolarització a la membrana de les zones veïnes desencadenant un nou potencial d’acció. Aquest nou potencial implicarà l’entrada de ions de Na+, i es repetirà tot el procés. � El potencial d’acció es condueix al llarg de tot l’axó i s’autoregenera en cada punt de la membrana � Conducció continua.
Conducció en fibres mielíniques : El potencial d’acció només es podrà produir en els Nòduls de Ranvier, perquè la resta de membrana està envoltada de mielina. El procés és el mateix, però les zones veïnes pròximes no podran generar el potencial perquè estan recobertes de mielina i no permeten l’entrada de Na+. Per tant, els corrents locals hauran de circular fins al proper Nòdul. Aquest corrent s’haurà debilitat, però tindrà prou força per despolaritzar la membrana en el proper Nòdul. � Conducció saltatòria � La velocitat de conducció és molt més ràpida perquè hi ha corrents locals al interior i exterior però no hi ha intercanvis fins als Nòduls. A més a més, és més econòmic perquè requereix menys energia. [Els canals de Na+^ controlats per voltatge i bombes Na+/K+^ només es troben als nòduls de ranvier].
29. Què és i com es produeix la conducció saltatòria del potencial d’acció?
La conducció saltatòria és el nom del tipus de conducció del potencial d’acció en les fibres mielíniques. En les fibres mielíniques, la membrana de l’axó està envoltada per la mielina i, per tant, no està en contacte directe amb el líquid extracel·lular. Això fa que l’intercanvi d’ions entre el fluid intracel·lular i l’extracel·lular, característic del potencial d’acció, només tingui lloc en les zones de l’axó que no estan envoltades de mielina, és a dir, en els nòduls de Ranvier. Per tant, els potencials d’acció en axons mielínics només s’autoregeneren en el nòduls de Ranvier. Així, doncs, mentre que en les fibres amielíniques la conducció del potencial d’axó és contínua (es produeix en cada punt de l’axó), en les fibres mielíniques es condueix de nòdul a nòdul i s’anomena conducció saltatòria.
30. Imagina una alteració neurològica que afecti principalment les prolongacions dels oligodendròcits fent-les més petites. Així, en els axons mielínics es trobarien internodes més curts i, com a mecanisme compensatori, s’observaria una major quantitat de nodes en els axons afectats. Creus que aquesta malaltia afectaria la conducció del potencial d’acció? Explica de quina manera?.
Sí. La conducció en fibres mielíniques (saltatòria) està caracteritzada per ser molt més ràpida que la conducció contínua, pel fet que no hi ha intercanvis de corrents locals interiors i exteriors (despolarització) fins que arriben als nòduls de Ranvier. Doncs, si hi ha una quantitat més gran de nòduls de Ranvier als axons, tot i que la velocitat seguirà sent superior a la que es dóna en conducció continua, serà més lenta en comparació a la que es dóna en axons mielínics no afectats.
Segons el lloc de contacte , tot i que es poden donar moltes combinacions possibles entre els tres elements principals de la neurona (dendrites, soma i axó), les més freqüents són les axosomàtiques (un axó presinàptic fent sinapsi sobre un soma postsinàptic), les axodendrítiques (un axó presinàptic fent sinapsi sobre una dendrita postsinàptica, o sobre una espina dendrítica), i les axoaxòniques (un axó presinàptic fent sinapsi sobre un axó postsinàptic).
Segons el lloc de contacte podem observar diferents efectes postsinàptics. Així, les sinapsis axosomàtiques solen ser inhibidores, mentre que les axodendrítiques solen ser excitadores. Les axoaxòniques acostumen a modular la quantitat de neurotransmissor que l’axó postsinàptic alliberarà sobre una tercera neurona.
5. Explica tot el procés des de l’arribada del potencial d’acció al terminal axònic fins que el NT es troba a l’espai sinàptic.
L’alliberament de neurotransmissor depèn de l’entrada de Ca2+ al botó terminal. El Ca2+ està més concentrat en el medi extracel·lular, per la qual cosa tendirà a entrar a la cèl·lula a favor del seu gradient químic. El Ca2+ travessa la membrana gràcies a canals iònics selectius controlats per voltatge que s’obren quan es despolaritza la membrana per l’arribada de potencials d’acció. Per a alliberar el neurotransmissor, les vesícules sinàptiques han de fusionar-se amb la membrana presinàptica. En situació de repòs, algunes vesícules estan unides al citoesquelet de la cèl·lula i d’altres estan fixades a les zones actives preparades per a fusionar-se amb la membrana presinàptica. Les vesícules s’uneixen al citoesquelet per mitjà d’unes proteïnes conegudes com a sinapsines I.Per la seva banda, les vesícules situades a les zones actives estan preparades per a formar un canal d’unió amb la membrana presinàptica (un porus de fusió). Aquest porus connectarà l’espai de dintre la vesícula amb l’espai extracel·lular. La despolarització del terminal presinàptic, amb la consegüent entrada de Ca2+, tindrà un doble efecte: o Les sinapsines I es fosforilaran per l’acció d’una proteïnquinasa dependent de Ca2+. D’aquesta manera, les vesícules unides al citoesquelet quedaran lliures per a poder-se fixar a les zones actives. o A les zones actives, la membrana vesicular es fusionarà amb la membrana presinàptica per mitjà del canal d’unió, de manera que el neurotransmissor que conté la vesícula sortirà a l’espai extracel·lular. Aquest procés de fusió de membranes es coneix com a exocitosi.
6. Per què són importants els neurotransmissors químics que arriben a la neurona postsinàptica en la gènesi del potencial d’acció d’aquesta cèl·lula? I per què és necessari el potencial d’acció per l’alliberació del neurotransmissor en les neurones presinàptiques? Una manca de Ca2+^ extracel·lular, pot afectar sobre la conducció de potencials d’acció al llarg de tot l’SNC, o només sobre l’alliberació de neurotransmissor?
Quan els neurotransmissors s’uneixen als receptors postsinàptics produeixen l’obertura o el tancament de canals iònics de manera immediata (receptor ionotròpic), amb el qual es produeix un potencial postsinàptic immediat o de forma més lenta (receptor metabotròpic). En alguns casos es produeixen PEPs (potencials postsinàptics excitadors) perquè el neurotransmissor produeix l’obertura dels canals de Na+ i de K+. Si la suma de les despolaritzacions (PEPs) i hiperpolaritzacions (PIPs) que arriba al conus axònic es suficient per despolaritzar la membrana, és a dir, s’arriba al nivell del llindar de descàrrega (aproximadament -55mV), es generarà un potencial d’acció al conus axònic. El potencial d’acció és necessari per l’alliberació dels neurotransmissors en les neurones presinàptiques perquè fa que s’obrin els canals de Ca++ controlats per voltatge. El Ca++ s’uneix a les vesícules presinàptiques i fa que aquestes es fusionin amb el botó terminal i alliberin el neurotransmissor (NT) per exocitosi. Sense Ca++ el potencial d’acció no produirà alliberació de NT, i sense alliberació de NT no es produiran nous potencials d’acció en les neurones següents. Així doncs, una manca de Ca++ extracel·lular afectarà directament sobre l’alliberació de NT i indirectament sobre la generació de potencials d’acció en les següents neurones, ja que el senyal no es podrà conduir a través del SN. Per exemple, si la quantitat d’acetilcolina alliberada amb cada impuls nerviós disminuís fins al punt que fos insuficient per generar nous potencials d’acció a les fibres musculars, es produiria una paràlisi complerta.
7. Descriu les bases iòniques i el mecanisme de propagació al llarg de la membrana neuronal dels potencials excitadors postsinàptics (PEP) i dels potencials inhibidors postsinàptics (PIP).
Observarem un potencial excitador postsinàptic (PEP) quan, a conseqüència de l’arribada a la neurona postsinàptica de molècules de neurotransmissor, en la membrana postsinàptica s’obrin canals iònics que permetin simultàniament l’entrada a la neurona d’ions de Na+ i la sortida d’ions de K+. Com que el gradient electroquímic del Na+ és superior al del K+, el resultat net serà una despolarització de la membrana postsinàptica.
Observarem un potencial inhibitori postsinàptic (PIP) quan, a conseqüència de l’arribada a la neurona postsinàptica de molècules de neurotransmissor, en la membrana postsinàptica s’obrin canals iònics que permetin la sortida de la neurona, a favor de gradient, d’ions de K+, o bé s’obrin canals iònics que permetin l’entrada a la neurona, a favor de gradient, d’ions de Cl-.
En ambdós casos, la propagació d’aquests potencials locals al llarg de la membrana postsinàptica es fa de manera electrotònica, és a dir, de manera passiva i amb pèrdua gradual d’intensitat, fins arribar a desaparèixer.
8. Què és un PEP? Quines són les seves bases iòniques?
S’anomena potencial excitador postsinàptic (PEP) la despolarització de la membrana postsinàptica que s’observa en les sinapsis excitadores. L’amplitud d’un PEP és proporcional a la intensitat de l’estimulació que el provoca. La despolarització arribarà com a màxim a valors de –10 mV. En cap cas hi haurà inversió de potencial, és a dir, la membrana no es despolaritzarà mai per sobre de 0 mV. A causa que els PEP són un tipus de potencial local, aquests es propaguen per la membrana de la cèl·lula de manera electrotònica (de manera passiva i amb pèrdua gradual d’intensitat), fins a desaparèixer. Els PEP s’originen per l’obertura de canals que permeten el pas simultani d’ions de Na+^ i K+. És a dir, el mateix canal deixa passar ions de Na+^ i K+. Aquests canals són controlats per lligand, és a dir, s’obren en resposta a la
unió d’un neurotransmissor amb una proteïna receptora.
9. En què consisteix la facilitació presinàptica? ()*
S’activa presinàpticament una sinapsi axoaxònica que provoca un increment en la despolarització presinàptica i, per tant, un augment en l’alliberament de NT. En la neurona postsinàptica s’observa un augment en l’amplitud del PEP, fet que incrementa la possibilitat de que s’originin potencials d’acció.
10. Què és un PIP? Quines són les seves bases iòniques? (inhibició postsinàptica)
S’anomena potencial inhibidor postsinàptic (PIP) la hiperpolarització de la membrana postsinàptica que s’observa en la inhibició postsinàptica. Els PIP són un tipus de potencial local, i, per tant, es propaguen per la membrana de manera electrotònica, fins a desaparèixer. L’amplitud dels PIP és proporcional a la intensitat de l’estímul que els desencadena. Generalment hiperpolaritzen la membrana postsinàptica fins a valors d’entre – 70 mV i –80 mV. Els PIP s’originen per l’obertura de canals iònics controlats per lligand. La unió del neurotransmissor amb el receptor fa que s’obrin un tipus de canals que hiperpolaritzen la membrana, concretament els canals de Cl-^ o de canals de K+. El Cl-^ està en més concentració fora de la cèl·lula, per tant tendirà a entrar a favor del seu gradient químic. En el cas del K+, més concentrat a l’interior de la cèl·lula, l’obertura de canals provocarà la seva sortida. En tots dos casos, el resultat final serà un augment de la negativitat a l’interior de la cèl·lula, és a dir, una hiperpolarització de la membrana que l’allunya del llindar per a produir un potencial d’acció.
15. Imagina que ampliem la sinapsi B de la imatge anterior per observar el procés d’alliberació dels neurotransmissors. Identifica els passos assenyalats a la imatge amb els números de l’1 al 6. Indica, de manera justificada, quin tipus de resposta s’observarà a la neurona postsinàptica quan el neurotransmissor interaccioni amb la neurona postsinàptica i explica perquè aquesta sinapsi afavorirà o dificultarà que es generi un potencial d’acció.
- Després de l’arribada d’un potencial d’acció a la membrana presinàptica, aquesta es despolaritza i s’obren els canals de Ca2+^ dependents de voltatge.
- Els ions de Ca2+^ que es troben a l’espai extracel·lular entren al botó terminal de la neurona presinàptica.
- L’entrada de Ca2+^ provoca que les vesícules sinàptiques que contenen els neurotransmissors es mobilitzin i s’acostin a les zones actives de la membrana presinàptica.
- La membrana vesicular es fusionarà amb la membrana presinàptica, de manera que el neurotransmissor que conté la vesícula sortirà a l’espai extracel·lular. Aquest procés de fusió de membranes es coneix com a exocitosi.
- Les molècules de neurotransmissor alliberades per exocitosi es difonen per l’espai sinàptic fins a arribar a la membrana postsinàptica. Allà interactuen breument amb receptors postsinàptics, que són proteïnes de membrana que reconeixen específicament un tipus de substància neurotransmissora. En aquest cas, la interacció neurotransmissor-receptor de canal de Na+/K+^ provoca l’obertura de canals iònics controlats per lligand, que al seu torn provoquen un PEP en la neurona postsinàptica.
6. Després d’aquest procés el NT es deslliga del receptor i ha de ser eliminat del espai sinàptic per a que no torni a unir-s’hi. Aquesta eliminació s’anomena inactivació del NT i es pot donar per degradació enzimàtica o per recaptació.
Tot i que les sinapsis axosomàtiques solen ser inhibidores, en aquest cas la resposta postsinàptica ha estat excitadora. S’ha produït un PEP al despolaritzar-se la membrana gràcies a l’obertura de canals Na+/K+, en comptes d’hiperpolaritzar-se (com hagués passat en cas d’haver-se obert canals C1-^ o K+). Doncs aquest fet afavorirà que es produeixi un potencial d’acció, ja que aquests són causa d’una despolarització que arribi a un llindar específic.
16. Què és la integració sinàptica? A quina part de la neurona es dóna? Enumera i descriu breument els tipus de mecanisme d’integració sinàptica que coneguis.
La integració sinàptica és la suma algebraica que es fa al segment inicial de tots els potencials excitadors postsinàptics (PEP) i els potencials inhibidors postsinàptics (PIP) que arriben a una neurona en un moment donat. La integració sinàptica es dóna al segment inicial de l’axó (o con axònic), ja que aquesta regió té una alta concentració de canals de Na+ controlats per voltatge i és aquí on s’integraran (sumaran) els potencials locals (PEPs i PIPs) provinents de les dendrites i el soma, el que determinarà si s’iniciarà el potencial d’acció (que necessita de la presència d’aquests canals de Na+ dependents de voltatge). Existeixen diferents tipus:
1- Sumació temporal : Es produeix quan arriben diverses informacions a una mateixa sinapsi en moments propers. Si immediatament després del primer potencial d’acció arriben consecutivament més potencials d’acció, quan encara no hagi acabat el primer PEP se li sumaran progressivament més PEP, augmentant l’amplitud i la durada del PEP final. Això facilitarà que es desencadeni un potencial d’acció o augmentarà la freqüència i durada del tren de potencials d’acció.
2- Sumació espacial :Es produeix quan arriben informacions al mateix temps a llocs diferents de la neurona postsinàptica. Si totes les informacions són excitadores, el resultat serà similar al de la sumació temporal, i augmentarà la probabilitat que es desencadeni un potencial d’acció o la freqüència i durada del tren de potencials d’acció. Si entre les diferents informacions que se sumen hi ha senyals inhibidors (PIP), aquests poden reduir l’amplitud i duració del PEP final, o modular la freqüència i duració dels trens de potencials d’acció postsinàptics.
17. Què és la inactivació del neurotransmissor? Explica els diferents tipus de mecanismes d’inactivació del NT.
La inactivación del neurotransmisor es el proceso por el cual se elimina el neurotransmisor del espacio sináptico. Los mecanismos de inactivación son: o Degradación enzimática : Por la acción de una enzima se rompen las moléculas de neurotransmisor. Los productos resultantes de esta degradación se denominan metabolitos, generalmente pasan a la sangre y después se eliminan por la orina. o Recaptación : El neurotransmisor es recaptado por el botón terminal de la neurona presinàptica gracias a un mecanismo de transporte activo de alta afinidad. Es un sistema específico, dado que cada proteína transportadora sólo recaptará un tipo de neurotransmisor, y de alta afinidad, dado que las moléculas de neurotransmisor tienen mucha facilidad para unirse a esta proteína transportadora.
18. Què són els receptors postsinàptics? Receptors inotròpics i receptors metabotròpics: semblances i diferències.
Els receptors postsinàptics estan formats per molècules proteiques insertades en la membrana cel·lular de la neurona postsinàptica. Cada NT pot ser reconegut per més d’un tipus de receptor. La unió NT-receptor provoca l’obertura de canals iònics (dependents de lligand). Depenent del tipus de canals que s’obrin es produirà un PEP o un PIP a la neurona postsinàptica. Tipus de receptors postsinàptics:
Els receptors ionotròpics contenen una regió receptora del neurotransmissor acoblada a un canal iònic. Així, quan el neurotransmissor s’uneix al receptor es produeix una obertura immediata del canal, el qual permet el pas d’ions.
Els receptors metabotròpics també contenen una regió receptora però no estan acoblats directament a cap canal iònic. Aquest tipus de receptors està unit a una proteïna G, de manera que quan el neurotransmissor s’uneix a un receptor metabotròpic es produeix un canvi en l’estructura d’aquesta proteïna G. Aquest canvi inicia una cascada bioquímica que acaba activant unes molècules anomenades segons missatgers, que són les que produeixen l’obertura dels canals iònics. L’activació de segons missatgers sol produir l’obertura de més d’un canal iònic, els quals solen estar a distància del receptor que rep el neurotransmissor. Així doncs, l’activació dels receptors metabotròpics produeix una amplificació del senyal del neurotransmissor, però de manera més lenta que en el cas dels receptors ionotròpics.
19. Què són els receptors presinàptics? Quins tipus hi ha?
Los receptores presinápticos son proteínas de membrana que reconocen de manera específica una substancia neurotransmisora y están localizados en la membrana presinaptica. Podemos dividir los receptores presinàptics en: o Autoreceptores : son los que reconocen la substancia que libera el mismo terminal sináptico. o Heteroreceptores : son los que reconocen las substancias liberadas por otras neuronas. Los receptores presinápticos, cuando son activados, modulan la liberación del neurotransmisor desde el terminal presináptico. Esta modulación es generalmente inhibidora, es decir, la activación de los receptores presinápticos hace que se libere menos cantidad de neurotransmisor. En el caso de los autoreceptores , es el mismo neurotransmisor liberado por el terminal sináptico el que inhibe/facilita su subsiguiente liberación; en el caso de los heteroreceptores es un segundo neurotransmisor el que realiza la acción.