


1 SN Excitabilidad
1 -1




















Prepara tus exámenes y mejora tus resultados gracias a la gran cantidad de recursos disponibles en Docsity

Gana puntos ayudando a otros estudiantes o consíguelos activando un Plan Premium

Prepara tus exámenes
Prepara tus exámenes y mejora tus resultados gracias a la gran cantidad de recursos disponibles en Docsity
Prepara tus exámenes con los documentos que comparten otros estudiantes como tú en Docsity
Encuentra los documentos específicos para los exámenes de tu universidad
Estudia con lecciones y exámenes resueltos basados en los programas académicos de las mejores universidades
Responde a preguntas de exámenes reales y pon a prueba tu preparación
Consigue puntos base para descargar
Gana puntos ayudando a otros estudiantes o consíguelos activando un Plan Premium
Comunidad
Pide ayuda a la comunidad y resuelve tus dudas de estudio
Ebooks gratuitos
Descarga nuestras guías gratuitas sobre técnicas de estudio, métodos para controlar la ansiedad y consejos para la tesis preparadas por los tutores de Docsity
Asignatura: Fisiologia, Profesor: Vicente , Carrera: Enfermería, Universidad: UNILEON
Tipo: Apuntes
1 / 28

Esta página no es visible en la vista previa
¡No te pierdas las partes importantes!
1 SN Excitabilidad
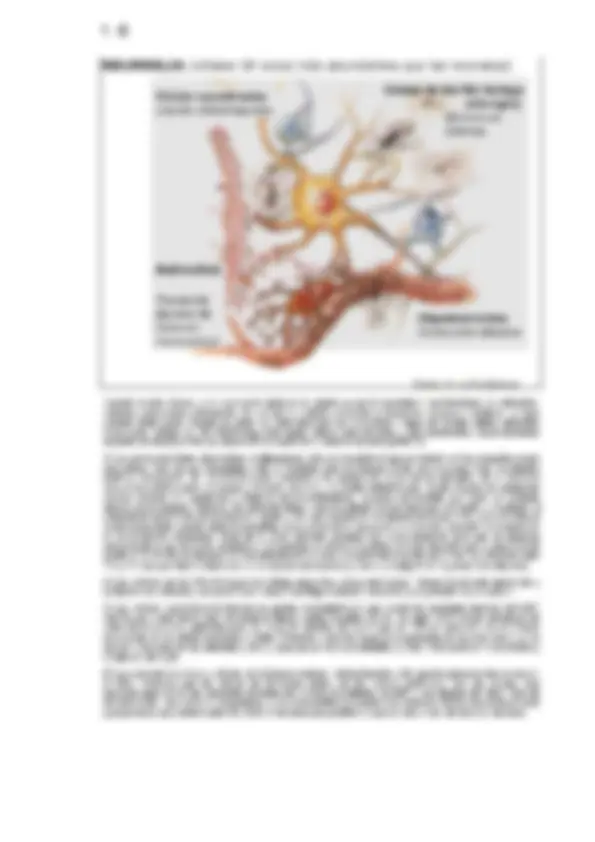
1 SN Excitabilidad 11 1. El sistema nervioso Excitabilidad 1.1 Organización funcional del sistema nervioso 1.2 Las neuronas. Tipos de neuronas 1.3 La neuroglia 1.4 Excitabilidad 1.5 Potencial de membrana 1.6 Introducción al impulso nervioso 1.7 Bases iónicas del potencial de acción 1.8 Umbral y períodos refractarios Animación A1.1 - El potencial de membrana en reposo Animación A1.2 - El potencial de acción Fisiologia - Dr. Juan Pablo Barrio Lera 1-2 Sistema Nervioso: Funciones Los receptores captan + Función sensorial información y la envían hacia centros nerviosos a través de las vías aferentes o. El sistema nervioso procesa + Función integradora la información y promueve respuestas apropiadas Las vías motoras (eferentes) + Función motora transmiten las respuestas a los efectores * Más de 100.000 millones de células nerviosas (neuronas) * Más de 1015 conexiones sinápticas Principios generales de funcionamiento del SN + Unidad estructural (neuronas + sinapsis) +. Niveles jerárquicos (centros inferiores £> superiores) + Estimulación-Inhibición Fisiologia - Dr. Juan Pablo Barrio Lera Podemos definir tres funciones generales en el SN: función sensorial función motora y función integradora. 1) La función sensorial corre a cargo de los diferentes receptores que se encuentran en la pil en los músculos, tendones y articulaciones, y en los órganos de los sentidos (ojo, oído, etc.) que captan la estímulos sensoriales, y que posteriormente envían la información a centros nerviosos superiores por las vías sensoriales (aferentes). 2) La función motora se encarga de responder a la información sensorial, a través de las vías motoras (eferentes), utilizando los efectores. Pueden considerarse como efectores el músculo esquelético y liso y las géndutas, exocrinas y endocrinas. ) Función integradora. La función principal del SN es la de procesar toda la información que recibe para promover respuestas adecuadas. Se estima que sólo el 1% de la función sensorial se integra en los centros superiores; el 99% restante es información rutinaria. Las diferentes zonas del SNC están dedicadas total o en parle a estas funciones. Estas funciones están llevadas a cabo por centros sensoriales, centros motores y centros de asociación en el cerebro y la médula espinal, y se desarrollan atendiendo a unos principios generales de funcionamiento: a) Unidad estructural.- los centros nerviosos, ya sean sensoriales, motores o de asociación, constan de millones de células nerviosas (neuronas) y de muchas más sinapsis que conectan estas células entre sí, con las de otros centros, o con neuronas periféricas. La morfología de las neuronas puede variar en las diferentes regiones, pero son las conexiones entre las células nerviosas los principales determinantes de su función. b) Niveles jerárquicos. Los centros nerviosos están organizados siguiendo una jerarquía, esto es, los centros sensoriales pueden dividirse en inferiores Y superiores, del mismo modo que los centros motores y de asociación. Esta jerarquía implica que solamente los centros inferiores se encuentran en contacto directo con las estructuras nerviosas periféricas. A fin de que los centros superiores se comuniquen con las estructuras periféricas, deben hacerlo a través de los centros inferiores, y viceversa. €) Por último, todos los centros nerviosos actúan según los principios básicos de estímu/ación-inhibición, estando la función determinada por el tipo de sinapsis establecidas entre ellos. 1-4 Organización (a | funcional del sistema nervioso Fisiologia - Dr. Juan Pablo Barrio Lera (Se trata de un ejercicio de memorización de los niveles funcionales del Sistema Nervioso) 1-5 ESTRUCTURA Y FUNCIÓN EN EL ENCÉFALO 1-7 Tipos de neuronas según su morfología A. Anaxónicas Bipolares Pseudo-unipolares Multipolares DOoDo según su función *+ Neuronas sensoriales ¿2 Interneuronas + Neuronas motoras (a) Anaxonica AÁxon Dendritas -Axon terminales terminales Axon sinapticos sinapticos Mm (b) Bipolar (c) Unipolar (d) Multipolar ¡0 ESTÍMULO e RESPUESTA, Fisiologia - Dr. Juan Pablo Barrio Lera Existen distintas clasificaciones anatómicas, histológicas (según tamaño, forma, tinción, etc.) y funcionales de las neuronas. Siguiendo un criterio func/ona/podemos encontrar: - Neuronas sensoriales o aferentes, encargadas de recibir la información sensorial (tacto, dolor, etc.). - Interneuronas o neuronas intercalares, que establecen comunicación entre las neuronas sensoriales y las motoras; y finalmente, - Neuronas /motoras o eferentes, quienes transmiten los impulsos desde el SNC hacia los órganos efectores (músculos y glándulas). Siguiendo un criterio morfológico podemos hablar de neuronas: - anaxónicas, presentes en el cerebro y en sistemas sensoriales como el ojo de los vertebrados, que no poseen terminación axónica propiamente dicha; - pseudo-unipolares, muy abundantes como neuronas sensoriales e interneuronas, especialmente en invertebrados; - bipolares, especialmente neuronas sensoriales (vista, olfato); - multipolares, el tipo más abundante, principalmente interneuronas y neuronas motoras, con funciones de integración en el SNC. 1-8 NEUROGLIA (células 10 veces más abundantes que las neuronas) A, Células ependimarias Líquido cefalorraquídeo " I Río Hortega - (microglia) Movimiento Defensa Transporte Barrera HE Nutrición Homeostasis Oligodendrocitos Conducción eléctrica Fisiologia - Dr. Juan Pablo Barrio Lera Durante mucho tiempo, a la 2eurog//a (glia) se le asignó un pepal secundario con funciones de estruclura, nutrición, aislamiento y transporte, ya que no son células excitables ni transmiten impulsos eléctricos, aunque ueden multiplicarse. Desde un punto de vista fisiológico se consideran 4 tipos de células gliales: astrocitos Imaciadlá), células de Del RioHortega (microglía), células ependimarias y oligodendrocitos, cuyas funciones aislantes las desempeñan las células de Schwann en el sistema nervioso periférico. 1) Los astrocitos tienen abundantes ramificaciones (pies perivasculares) que se insertan en los pequeños vasos sanguíneos. Son células conectadas entre sí mediante uniones huecas, Entre sus funciones más importantes están el transporte de nutrientes a las neuronas y la constitución, junto con la piamadre, de la barrera hematoencefálica entre la sangre y el tejido cerebral, una malla defensiva que impide el paso de sustancias nocivas (excepto en hipotálamo y órganos circumventriculares, regiones del encéfalo que están en contacto directo con la sangre). Además, los astrocitos tienen una importante función fisiológica al ayudar a mantener el potencial de membrana de la neurona: captan el K+ que expulsa la membrana de la neurona, devolviéndolo al medio extracelular cuando éstas lo necesitan (tamponamiento espacial. La actividad neuronal hace aumentar la concentración extracelular local de K. Este aumento produce una despolarización glial que se dispersa pasivamente a los astrocitos acoplados. El potencial de membrana (Em) es más negativo que el potencial de equilibrio del K (Ek) al elevarse la concentración de K como resultado de la acción neuronal. La diferencia entre Em y Ek hace penetrar K donde su concentración está elevada y hace que salga K en regiones más alejadas). 2) Las células de Del Río-Hortega son células pequeñas y muy ramificadas. Tienen movimiento ameboide y su función es defensiva, comportándose como macrófagos retirando astrocitos y oligodendrocitos muertos. 3) Las células ependimarías forman un epitelio monostratificado Pl reviste las cavidades internas del SNC (ventrículos y epéndimo) q contienen el líquido cefalorraquideo | CR). Se unen entre sí por complejos de Unión similafes a los epiteliales pero sin zona de oclusión, de modo qué el LCR se comunica con el entorno extracelular de las células nerviosas y gliales. Presentan además largas prolongaciones en su zona basal que se asocian a los pies de los astrocitos y en su zona apical microvellosidades y cillos. Intervienen en la secreción y circulación del LCR. 4) Los oligodendrocitos y células de Sehwann realizan idéntica función, sólo que los primeros intervienen en el SNC, "mientras que las células de Schwann actúan en los nendos, poriÉricos. Son las células más espectaculares de la glía, presentes en todos los nervios importantes del SNP y en algunos del SNC. Son de tamaño medio, con pocas prolongaciones, y su característica es constituir la vaina de mielina que rodea al axón y proporciona una cubierta aislante a la fibra nerviosa que posibilita la rapida conducción del impulso nervioso. 1-10 Potencial de membrana La membrana plasmática en reposo presenta una diferencia de potencial eléctrico constante, que representa un estado de equilibrio sin ganancia o pérdida neta de ¡ones + Julius Bernstein relacionó la +//- sel membrana con los fenómenos 7 bioeléctricos a comienzos del siglo XX FS Ñ Bases del potencial de membrana: + Permeabilidad selectiva de la membrana e Distribución desigual de iones a ambos lados + Existencia de bombas de intercambio de ¡ones Fisiologia - Dr. Juan Pablo Barrio Lera Julius Bernsteín (1902, 1912) propuso que las células excitables están rodeadas por una membrana permeable selectivamente a los iones potasio en reposo, y que durante la excitación la permeabilidad de la membrana a otros jones se ve incrementada. Su "hipótesis de la membrana" explicaba el potencial de reposo del nervio y del músculo como un potencial de difusión constituido por la tendencia de los iones cargados positivamente a difundir desde su elevada concentración en el citoplasma hasta su baja concentración en la solución extracelular, mientras que otros iones eran retenidos. Durante la excitación, la negatividad interna se perdería temporalmente en tanto que otros iones podrían difundir a través de la membrana, cortocircuitando de hecho el potencial de difusión del potasio. La cara interna de una membrana celular presenta generalmente más carga negativa en su vecindad inmediata, y la superficie extracelular, por el contrario, tiene un predominio de carga eléctrica positiva. Esta diferencia de carga eléctrica genera una diferencia de potencial que puede tener una amplitud de hasta 100 mV, y tiene el efecto de que las cargas positivas son atraídas hacia el interior celular, y las negativas son alraídas hacia el exterior. Esta diferencia de potencial se denomina potencial de membrana, y se expresa siempre como la diferencia entre el voltaje dentro de la célula menos el voltaje fuera de la célula. Puesto que el interior es generalmente negativo (y el exterior positivo), el potencial de membrana normal será negativo. 1-11 Diferencia de potencial Flujo de cargas A negativas E A MÍ Flujo de cargas positivas DDP = (potencial intracelular) — (poteRabedrácellar) Fisiologia - Dr. Juan Pablo Barrio Lera La diferencia de potencial entre dos puntos permite el movimiento de cargas eléctricas, esto es, la corriente eléctrica. Por convención, el punto de mayor concentración de carga posiliva es el punto de mayor potencial eléctrico. Entre dos puntos con diferente potencial se establece un gradiente de potencial, que permitirá la aparicion de una corriente eléctrica si existe un vehículo para el movimiento de las cargas (iones). Las cargas posilivas se mueven siguiendo el gradiente de potencial eléctrico (y las negativas en sentido contrario). El potencial de membrana expresa la diferencia de potencial entre el lado extracelular y el lado intracelular de la membrana plasmática. En condiciones normales el lado extracelular está conectado a masa (0 voltios), por lo que a efectos prácticos la diferencia de potencial registrada corresponde al potencial intracelular. La diferencia de potencial puede medirse conectando dos electrodos (que pueden ser conductores metálicos debidamente acondicionados o micropipetas de vidrio rellenas de electrolitos) uno a cada lado de la membrana y uniendo estos electrodos a su vez en su otro extremo con un milivoltimetro. Se establecerá un flujo de electrones a través de los conductores desde el lado negativo hasta el lado positivo. La magnitud del flujo, detectada por el medidor, es proporcional a la diferencia de potencial. En la práctica deben tomarse precauciones especiales para asegurar que el drenaje de carga ocurrido a través de los electrodos no modifique el voltaje original. El potencial de membrana tiene una magnitud diferente en los diferentes tipos de células. Varia desde -10 mV en los eritrocitos hasta cerca de -90 mV en las células musculares esqueléticas y cardiacas. La magnitud de la fuerza que representa el potencial de membrana puede apreciarse si nos damos cuenta de que cuando los ¡ones potasio se encuentran diez veces más concentrados en un lado de la membrana, su difusión hacia el lado menos concentrado puede detenerse por completo cuando hacemos que a este gradiente de concentración se oponga un gradiente de potencial de solamente 60 mV. 1-13 En el equilibrio: - El gradiente eléctrico iguala al gradiente de concentración - La suma de fuerzas a ambos lados es cero ¿F-E,, + RT In] bor,11_, lion... ] E = potencial de equilibrio del ion (V) R = 8,31] K-1 mol-1 T = Temperatura absoluta (K) z = valencia F = constante de Faraday (96500 C mor-1) Fisiologia - Dr. Juan Pablo Barrio Lera Cuanto mayor sea el gradiente de concentración, tanto mayor será el potencial de equilibrio. Este hecho permite calcular el potencial de equilibrio de un determinado ión siguiendo la ecuación de Nernst, que liene el enunciado siguiente: E=- (RIMZF) Inflión dentroJflón fuera)) donde E es el potencial de equilibrio, R es la constante de los gases, T la temperalura absoluta, z es la carga del ión, y F es el número de Faraday (o la carga eléctrica por mol, 96500 Coulomb). [ión dentro] e [ión fuera] son las concentraciones del ión que estamos considerando dentro y fuera de la célula, respectivamente. A temperatura corporal (37 *C), si el potencial de membrana se expresa en milivoltios el factor RT/F tiene un valor de 26,6 y pasando a logaritmos en base diez (abreviado como /og; recordemos que |1/x)=2 303 tog(x) tenemos finalmente que: E = 61 log(lióndentrollióntuera]) mV 1-14 Potencial de equilibrio del potasio : T=37*C z=+1 [K+lext = 5,5 mM Ek = -26,6 mV * 2,303 log(150/5,5) [K+int = 150 mM = -88 mV Potencial de equilibrio del sodio : T=37 z=+1 [Na+Jext = 140 mm. ENa = -26,6 mV * 2,303 log(15/140) [Na+Jint = 15 mM mM Fisiologia - Dr. Juan Pablo Barrio Lera Atemperalura corporal (37 *C), si el potencial de membrana se expresa en milivoltios el factor RT/F tiene un valor de 26,6 y pasando a logaritmos en base diez (abreviado como /og; recordemos que |1/x)=2.303 log(x) tenemos finalmente que: E = -61 logílióndentrol/[iónfuera]) mV A37C, si [K*lexi= 5,5 mM y [K*int = 150 mM, el potencial de equilibrio del potasio será: E= -61 log(150/5,5) = -88 mV Analogamente, si [Na*Jex= 130 mM y [Na*Jint = 15 mM, el potencial de equilibrio del sodio será: E=-61 log(15/149) = +59 mV 1-16 El potencial de membrana es distinto del potencial de equilibrio Sodio (Na+) El potencial de equilibrio del Na+ es +59 mV, pero el potencial de membrana es -70 mV. + Tanto el gradiente químico como el eléctrico favorecen la entrada de Na+. e Entra poco Na+ porque hay pocos canales de Na+ abiertos en reposo. La bomba Na+/K+ expulsa 3 Na+ al exterior introduciendo 2 K+ en la célula. Potasio (K+) + El potencial de equilibrio del K+ es -88 mV, pero el potencial de membrana es -70 mV. e El gradiente eléctrico no es suficiente para impedir la salida de K+ a través de los canales abiertos. + Este goteo de K+ por difusión se compensa por la acción de la bomba Na+/K+, Coruro (Cl-) El potencial de equilibrio del Cl- es -70 mv. + Puesto que el potencial de membrana es también de -70 mV, los ¡ones Cl- se distribuyen pasivamente. + Nose necesita bombeo para mantener niveles de Cl- constantes. Fisiologia - Dr. Juan Pablo Barrio Lera El potencial de membrana es distinto del potencial de equilibrio. En el caso del sodio (Na*), su potencial de equilibrio a 37 *C es +59 mV (compruébelo), pero el potencial de membrana es -70 mV. Tanto el gradiente químico como el eléctrico favorecen la entrada de Na. Entra poco Na* porque hay pocos canales de Na* abiertos en reposo, y lo poco que entra vuelve a salir por efecto de la bomba Na*/K+. Para el potasio (K*+), el potencial de equilibrio a 37 “C es de -88 mV (compruébelo), pero el potencial de membrana es -70 mV. El gradiente eléctrico no es suficiente para impedir la salida de K* a través de los canales abiertos; sin embargo, esta salida por difusión queda compensada por la acción de la bomba Nat/K+. En el caso del cloruro, el potencial de equilibrio a 37 *C es de -70 mV. Puesto que el potencial de membrana es también de -70 mV, no hay movimiento neto de iones CF. No se necesita bombeo para mantener niveles de CF constantes. El potencial de membrana debe ser entendido como la suma de las contribuciones de los distintos jones que atraviesan la membrana. Puede ser descrito por medio de la ecuación de Goldman-Hodgkin-Katz, que básicamente es una extensión de la ecuación de Nernst considerando las permeabilidades invididuales de la membrana a los iones sodio, potasio y cloruro: Em =-RTIF In(fpNa(Naji + pK[Kji + pCI[CIe)(pNa[Naje + pK[K]e + pCI[CIJ)) 1-17 Estimulación Despolarización: Fuido extacelier Disminución del d>eb : E E gradiente eléctrico Le ER 8; aora MEC ed - Potencial de membrana menos negativo Fisiologia - Dr. Juan Pablo Barrio Lera Las células que pueden transmitir mensajes eléctricos o impulsos a lo largo del sistema nervioso se denominan neuronas. La existencia de estos mensajes eléctricos o impulsos nerviosos puede demostrarse con facilidad. Si se secciona el nervio de la pata de un animal, la pata queda paralizada. Los músculos de la pata permanecen intactos, y funcionarán si son estimulados directamente con corrientes eléctricas débiles, pero si no es ariificialmente nunca recibirán la orden de moverse. Transcurrido cierto tiempo la parálisis puede remitir y la vuelta a la actividad normal coincidirá con la regeneración de los axones cortados, reanudando las conexiones originales. Un potencial de membrana en reposo de -70 mV implica que el lado interior de la membrana es negativo mientras el exterior es positivo. Debido a esta diferencia entre los lados interno y externo de la membrana, se dice que ésta se encuentra polarizada. Por definición, se dice que la membrana se Jespo/ariza cuando el potencial de membrana se vuelve progresivamente menos negativo (o más positivo) que el potencial de reposo; por el contrario, si el potencial se hace más negativo (o menos positivo), la membrana se repo/ariza o hiperpolariza (en relación al potencial de membrana en reposo: una hiperpolarización es un potencial de membrana mas negativo que el potencial de membrana en reposo). Cuando se estimulan las fibras nerviosas con descargas eléctricas, el impulso nervioso siempre surge en el electrodo cargado negativamente. La atracción de ¡ones positivos y repulsión de los negativos hace que el potencial de membrana (la diferencia de potencial entre el citoplasma y el fluido extracelular) sea menos negativo y la membrana se despolariza. De modo general, podemos decir que un estínulo solamente será eficaz sí consigue despolarizar la membrana. 1-19 Estimulación SUB-UMBRAL Aumenta la salida de ¡ones potasio... ...Ocasionando repolarización Fisiologia - Dr. Juan Pablo Barrio Lera Existen dos tipos de canales para el potasio. Un tipo de ellos se regula por voltaje, y en su mayor parte se encuentran cerrados en reposo, cuando el potencial de membrana está próximo a -70 mV. El otro tipo, el canal de fuga de potasio, no depende del voltaje: se encuentra abierto siempre y provee la vía para la salida continua aunque escasa de potasio que es responsable del potencial de membrana en reposo. Un potencial de membrana en reposo de -70 mV implica que el lado interior de la membrana es negativo mientras el exterior es positivo. Debido a esta diferencia entre los lados interno y externo de la membrana, se dice que ésta se encuentra polarizada. Por definición, se dice que la membrana se despolariza cuando el potencial de membrana se vuelve progresivamente menos negativo que el potencial de reposo; por el contrario, si el potencial se hace más negativo, la membrana se hiperpolariza. Una estimulación origina un cambio en el potencial de membrana. Si el estímulo es débil, el potencial de membrana se modificará poco; no obstante, disminuye el gradiente eléctrico y los iones potasio, que ahora pueden pasar más libremente al exterior, hacen recuperar el equilibrio elevando nuevamente el gradiente eléctrico (ver figura). Sin embargo, si el estímulo es mayor que un nivel crítico llamado umbral, entonces se abren los canales de sodio dependientes de voltaje. Los iones sodio, a elevada concentración fuera del axon, abandonan sus contralones negativos y penetran con la suficiente rapidez como para contrarrestar los ¡ones potasio que siguen saliendo. El interior de la célula se inunda de carga positiva en el punto de estimulación, y la polaridad se invierte: ahora el interior es positivo y el exterior negativo. Un momento más tarde los canales de Na se cierran y se abre otro tipo de canales de K. La membrana se vuelve entonces más permeable que antes al K. Los ¡ones K salen al exterior, haciendo el potencial de membrana aún más negativo, hasta situarlo muy cerca del nivel del potencial de equilibrio para el K. Finalmente, tras varios milisegundos, los canales adicionales de K se cierran y la permeabilidad de la membrana vuelve a su condición de reposo. 1-20 POLARIZACION (Reposo) ALTA PERMEABILIDAD AL K+ Fisiologia - Dr. Juan Pablo Barrio Lera Cuando un axon es estimulado tienen lugar una serie de cambios antes de que el mensaje elécirico llegue a término en la célula musculaar, glandular o nerviosa destinataria del mismo. Entre estos cambios, los cambios eléctricos son relativamente fáciles de medir e interpretar. En reposo, el axon se comporta como cualquier otra célula; su interior presenta carga eléctrica negativa en comparación con el exterior. Si colocamos electrodos a ambos lados de la membrana del axon, se registra una diferencia de potencial próxima a 70 mV a través de la membrana, con el interior negativo: el potencial de membrana es pues de -70 mV. Cuando el axon es estimulado, este voltaje se invierte, pasando momentáneamente a positivo en el interior y volviendo rápidamente al estado negativo original en reposo. Este cambio repentino en potencial de membrana que acompaña a la estimulación supraumbral se denomina potencial de acción. Aparece en primer lugar en el punto de estimulación y después se registra progresivamente a lo largo del axon. Cuanto más apartado esté el punto de estimulación del punto de registro más tiempo tardará en aparecer el potencial de acción. En otras palabras, el potencial de acción comienza en el punto de estimulación y viaja a lo largo del axon hacia el músculo. La velocidad de este movimiento es idéntica a la velocidad del "mensaje" tal como describimos antes. De hecho, comparando las propiedades de los "mensajes" con las de los potenciales de acción se llega a la conclusión de que unos y otros son lo mismo: los "mensajes" son potenciales de acción. En el punto máximo del potencial de acción, el exterior de la membrana está cargado negativamente, y esta región negativa es trasladada a lo largo del axon. Una forma de visualizar el potencial de acción es una onda de electronegatividad que viaja a lo largo del axon.