¡Descarga Propiedades Eléctricas de las Células del Corazón: Potenciales de Acción y Conductividad y más Diapositivas en PDF de Cardiología solo en Docsity!
Elementos de la función cardíaca
C A P Í T U L O 16
PROPIEDADES ELÉCTRICAS
DEL CORAZÓN
Las células del corazón, como las neuronas, son excita- bles y generan potenciales de acción. Estos potenciales de acción inician la contracción, determinando así la fre- cuencia cardíaca. Los trastornos de la actividad eléctrica pueden causar graves alteraciones en el ritmo cardíaco que, en ocasiones, resultan mortales. En esta sección se analizan las propiedades eléctricas de las células cardíacas. Además, se explica la contribu- ción de estas propiedades eléctricas al electrocardio- grama (ECG). En una sección posterior, se abordará el inicio de la contracción como consecuencia de la activi- dad eléctrica de las células cardíacas.
El potencial de acción cardíaco
La figura 16-1 representa los potenciales de acción que se encuentran en distintas células cardíacas. En el corazón se producen dos tipos fundamentales de potenciales de ac- ción, que se muestran en la figura. Uno de estos tipos, el de respuesta rápida, se produce en los miocitos ventriculares y auriculares normales y en las fibras de conducción espe- cializadas (fibras de Purkinje del corazón), y se divide en cinco fases. La fase ascendente rápida del potencial de ac- ción se denomina fase 0. Esta fase de ascenso rápido va seguida de un breve período de repolarización parcial pre- coz (fase 1) y de una fase de meseta (fase 2), que persiste durante 0,1-0,2 segundos. Después, la membrana se repola- riza (fase 3) hasta que se recupera de nuevo el estado de reposo de la polarización (fase 4) (en el punto e). La repola- rización final (fase 3) es más lenta que la despolarización (fase 0). El otro tipo de potencial de acción, la respuesta lenta, se observa en el nódulo sinoauricular (SA) , que es el marcapasos natural del corazón, y en la región del nódulo auriculoventricular (AV) , que es el tejido especializado res- ponsable de conducir el impulso cardíaco desde las aurícu- las a los ventrículos. Las células de respuesta lenta no muestran la fase de repolarización precoz (fase 1). Otras diferencias en las propiedades eléctricas entre las célu- las de respuesta rápida y lenta son las siguientes. El poten- cial de membrana en reposo de las células rápidas (fase 4) es considerablemente más negativo que el potencial de las células lentas. Además, la pendiente de la corriente ascen- dente (fase 0), la amplitud del potencial de acción y el so- bredisparo son mayores en las células de respuesta rápida que en las lentas. La amplitud del potencial de acción y la inclinación de la pendiente de la fase ascendente son deter- minantes importantes de la velocidad de propagación a lo largo de las fibras miocárdicas. En el tejido cardíaco de res- puesta lenta, el potencial de acción se propaga más lenta- mente, y la conducción se puede bloquear con mayor faci- lidad que en el tejido cardíaco de respuesta rápida. La conducción lenta y la mayor tendencia al bloqueo de la mis- ma aumentan el riesgo de sufrir algunas alteraciones del ritmo (v. la sección sobre la reentrada). Como se ha comentado, el potencial de acción inicia la contracción del miocito. Las relaciones entre el potencial de acción y la contracción del músculo cardíaco se ilus- tran en la figura 16-2. La despolarización rápida (fase 0) antecede al desarrollo de la fuerza, y el final de la repolari- zación coincide aproximadamente con la fuerza máxima. La relajación del músculo se produce sobre todo durante la fase 4 del potencial de acción. La duración de la con- tracción suele ser paralela a la del potencial de acción. Las diversas fases del potencial de acción cardíaco se asocian con cambios en la permeabilidad de la membra- na celular, especialmente a los iones Na+, K+^ y Ca++. Los cambios en la permeabilidad de la membrana celular modifican la velocidad de desplazamiento de los iones a través de la misma y, de este modo, modifican el voltaje de la membrana (Vm). Estos cambios de permeabilidad se deben a la apertura y cierre de los canales iónicos es- pecíficos de cada ión concreto (v. capítulos 1 y 2). Al igual que sucede con las demás células corporales, la concentración de K+^ dentro de las células musculares car- díacas ([K+]i) supera a la extracelular ([K+]o). En el caso del Na+^ y el Ca++, el gradiente de concentración es inverso. En la tabla 16-1 se recoge la estimación de las concentraciones extracelulares e intracelulares de Na+, K+^ y Ca++^ y los poten- ciales de equilibrio de Nernst (v. capítulo 1).
Voltaje de la membrana en reposo
La membrana celular en reposo muestra una permeabili- dad relativamente alta para el K+, mientras que la per- meabilidad para el Na+^ y el Ca++^ es bastante menor. Dado que existe un gradiente químico para el K+^ y Vm, el K+ tiende a salir de la célula por difusión. Cualquier flujo de K+^ que se produce con el potencial en reposo de la mem- brana (es decir, durante la fase 4) se produce principal- mente a través de unos canales específicos del K+. En las membranas de las células cardíacas existen varios tipos de canales del K+. La apertura y el cierre de algunos de estos canales se regulan por Vm, mientras que otros se controlan por señales químicas (p. ej., la concentración de acetilcolina extracelular). El canal del K+^ específico a través del cual pasa el K+^ durante la fase 4 es un canal regulado por el voltaje que conduce la corriente de en- trada de K +^ rectificadora. Esta corriente se representa como IK1 y se comenta de forma detallada más adelante. De momento, sólo es necesario saber cómo se genera esta corriente. La dependencia de Vm de la conductancia y las concen- traciones intracelulares y extracelulares de Na+, K+^ y otros iones se describe mediante la ecuación de la conductan- cia de cuerda (v. capítulo 2). En una célula cardíaca en
Capítulo 16 Elementos de la función cardíaca 293 © ELSEVIER. Fotocopiar sin autorización es un delito. 40 0
0 100 200 300 0 100 200 300 Tiempo (ms) Tiempo (ms) Miliv oltios 40 0
Miliv oltios
A
B
Respuesta rápida Respuesta lenta (^1 ) PRE PRR PRE PRR 2 3 3 4 4 0 0 ● Figura 16-1. Potenciales de acción en las fibras cardíacas de respuesta rápida ( A ) y lenta ( B ). Las fases de los potencia- les de acción están marcadas (v. más detalles en el texto). El pe- ríodo refractario eficaz (PRE) y el período refractario relativo (PRR) también están marcados. Obsérvese que, en comparación con las fibras de respuesta rápida, el potencial de reposo de las de res- puesta lenta es menos negativo, la fase ascendente (fase 0) del potencial de acción tiene menos pendiente, la amplitud de los potenciales es menor, falta la fase 1 y el PRR llega ampliamente a la fase 4, después de que todas las fibras se hayan repolarizado por completo. 16 0 Tiempo (ms) Potencial de acción Fuerza contráctil Miliv oltios
● Figura 16-2. Relaciones temporales entre las fuerzas ge- neradas y los cambios en el potencial transmembrana en una delgada tira de músculo ventricular. (Reproducido de Kavaler F et al. Bull NY Acad Med 41:5925, 1965.) ● Tabla 16-1. Concentraciones y equilibrio de iones intracelulares y extracelulares y potenciales de equilibrio en las células musculares cardíacas Ión Concentraciones extracelulares (mM) Concentraciones intracelulares (mM) Potencial de equilibrio (mV)* Na+^145 10 K+^4 135 - 94 Ca++^2 10 -^4 *Las concentraciones intracelulares son estimaciones de las concentraciones libres dentro del citoplasma. Datos de Ten Eick RE et al. Prog Cardiovasc Dis 24:157, 1981. reposo, la conductancia al K+^ (gK) es unas 100 veces mayor que la conductancia al Na+^ (gNa). Por tanto, Vm se parece al potencial de equilibrio de Nernst para el K+. En consecuen- cia, las alteraciones de la [K+] extracelular pueden modi- ficar de forma significativa Vm, en el sentido de que la hipopotasemia produce una hiperpolarización y la hiper- potasemia una despolarización. Por el contrario, gNa es tan pequeño en la célula en reposo que los cambios de [Na+]o no afectan de forma significativa a Vm.
Potenciales de acción de respuesta rápida
Génesis de la fase ascendente (fase 0)
Cualquier estímulo que despolarice de forma brusca Vm hasta un valor crítico (umbral) induce un potencial de acción. Las características de los potenciales de acción de respuesta rápida se exponen en la figura 16-1, A. La despolarización rápida (fase 0) se debe de forma casi ex- clusiva a la entrada de Na+^ al miocito como consecuencia del aumento súbito de gNa. La amplitud del potencial de acción (cambio de potencial durante la fase 0) depende de la [Na+]o. Cuando ésta [Na+]o disminuye, la amplitud del potencial de acción también lo hace, y cuando esta concentración de Na+^ pasa de los 140 mEq/l normales a unos 20 mEq/l, la célula deja de ser excitable. Cuando el potencial de membrana, Vm, se despolariza sú- bitamente desde –90 mV a un valor umbral de unos –65 mV, las propiedades de la membrana celular cambian de for- APLICACIÓN CLÍNICA Las respuestas rápidas pueden convertirse en respuestas lentas en determinadas situaciones patológicas. Por ejem- plo, en la cardiopatía coronaria, una región del músculo cardíaco queda privada de su irrigación normal. En conse- cuencia, aumentará la [K+] en el líquido intersticial que rodea al músculo afectado, porque se pierde K+^ a partir de las células mal perfundidas (o isquémicas). Los potenciales de acción de algunas de estas células se convierten de una respuesta rápida a otra lenta. La conversión de una res- puesta rápida a otra lenta por aumento de la [K+] intersti- cial se representa en la figura 16-13.
Capítulo 16 Elementos de la función cardíaca 295 © ELSEVIER. Fotocopiar sin autorización es un delito. A NIVEL CELULAR ( cont. ) paso de una corriente de 1,5 pA. Durante la primera y segunda aperturas del canal, se produjo la apertura de otro segundo canal, pero durante períodos de sólo 1 ms. En los breves momentos en que ambos canales estuvieron abiertos de forma simultánea, la corriente total fue de 3 pA. Después de que se cerrara el primer canal por terce- ra vez, ambos permanecieron cerrados durante una gran parte del registro, aunque la membrana mantuvo un valor constante de –45 mV. El cambio global de la conductancia iónica de toda la membrana celular en un momento determinado refleja el número de canales abiertos en este momento. Como los canales individuales se abren y cierran al azar, la conductan- cia global de la membrana refleja la probabilidad estadísti- ca de que cada uno de ellos esté abierto o cerrado. Las características temporales del proceso de activación repre- sentan la evolución temporal de la creciente probabilidad de que estén abiertos unos canales específicos, pero no las características cinéticas de las compuertas de activación de los canales individuales. Del mismo modo, las característi- cas temporales de la inactivación reflejan la evolución tem- poral de la probabilidad decreciente de que los canales es- tén abiertos, pero no las características cinéticas de las compuertas de inactivación de los canales individuales. 0 1, 3 4, 10 ms pA Corriente por el canal 1 Corriente por el canal 2 ● Figura 16-4. Corriente (en picoamperios) a través de dos canales individuales del Na+^ en una célula cardíaca en cultivo, registrada con la técnica de la pinza-parche. El voltaje de mem- brana se mantuvo en –85 mV y después se cambió de forma súbita a –45 mV en el lugar marcado por la flecha, y se mantuvo con este potencial para el resto del registro. (Reproducido de Cachelin AB et al. J Physiol 340:389, 1983.)
Génesis de la repolarización precoz (fase 1)
En muchas células cardíacas que tienen una meseta pro- minente, la fase 1 es un período breve y precoz de repo- larización limitada. Esta breva repolarización genera una incisura entre el final de la fase ascendente y el comienzo de la meseta (v. figs. 16-1 y 16-3). La repolarización es breve, porque se activa una corriente de salida transito- ria ( Ito ) principalmente debida al K+. La activación de los canales del K+^ durante la fase 1 determina una breve sali- da de K+^ de la célula, porque el interior de la misma tiene carga positiva y la [K+]i supera con mucho la [K+]o (v. figu- ra 16-3). La célula se repolariza de forma breve y parcial como consecuencia de esta salida transitoria de K+. El tamaño de la incisura en la fase 1 varía según la cé- lula cardíaca. Es prominente en los miocitos de las regio- nes epicárdica y miocárdica media de la pared ventricular izquierda (fig. 16-5) y en las fibras de Purkinje ventricula- res. Sin embargo, esta incisura es despreciable en los miocitos de la región miocárdica del ventrículo izquier- do (v. fig. 16-5), dada la menor densidad de canales Ito en estas células. La incisura también es menos aparente en presencia de 4-aminopiridina, que bloquea los canales del K+^ responsables de la corriente Ito.
Génesis de la meseta (fase 2)
Durante la meseta del potencial de acción, se produce la entrada de Ca++^ al miocito a través de los canales del cal- cio (v. más adelante), que se activan e inactivan mucho más lentamente que los canales del Na+^ rápidos. Durante la parte plana de la fase 2 (v. figs. 16-1 y 16-3), este flujo de entrada de Ca++^ se contraequilibra por la salida de K+. El potasio sale por canales que conducen principalmente co- rrientes Ito, IK e IK1. La corriente Ito es responsable de la fa- se 1, como se ha descrito anteriormente, pero no se inac- tiva por completo hasta terminar la fase 2. Las corrientes IK e IK1 se describen más adelante en este capítulo. El Ca++^ entra en la célula a través de unos canales del Ca++^ regulados por el voltaje, que se activan cuando Vm se va haciendo progresivamente menos negativa duran- te la fase ascendente del potencial de acción. Se han des- crito dos tipos de canales del Ca++^ ( tipos L y T ) en el tejido cardíaco. Algunas de sus características más importan- tes se recogen en la figura 16-6. Los canales de tipo L se llaman así porque una vez abiertos se inactivan lenta- mente v. (fig. 16-6, imagen inferior) y consiguen una co- rriente de calcio de «larga duración». Son el tipo princi- pal de canal del calcio en el corazón, y se activan durante la fase ascendente del potencial de acción cuando Vm lle- ga a –20 mV. Los canales de tipo L se bloquean por los antagonistas del calcio , como el verapamilo, amlodipino y diltiazem (fig. 16-7). Los canales del Ca++^ de tipo T (o transitorios) son mu- cho menos abundantes en el corazón. Se activan con po- tenciales mucho más negativos (unos –70 mV) que los de tipo L, y se inactivan con mayor rapidez que los canales de tipo L (fig. 16-6, imagen superior). Dado que los canales de tipo L son los más abundan- tes, la discusión que sigue se centra en sus propiedades y función. La apertura de los canales del Ca++^ provoca un incremento de la conductancia al Ca++^ (gCa) y una corrien- te (ICa) poco después de la fase ascendente del potencial de acción (v. fig. 16-3). Como la [Ca++]i es mucho menor que la [Ca++]o (v. tabla 16-1), el aumento de gCa potencia la entrada de calcio a la célula durante toda la meseta. Esta entrada de calcio está implicada en el acoplamiento en- tre la excitación y la contracción, como se describirá más adelante (v. también el capítulo 13). Diversos neurotransmisores y fármacos pueden influir de forma importante sobre gCa. El neurotransmisor adrenér- gico noradrenalina, el agonista del receptor β-adrenérgico isoproterenol y otras catecolaminas fomentan la gCa, mien- tras que el neurotransmisor parasimpático acetilcolina re- duce la gCa. La estimulación de gCa por las catecolaminas es el principal mecanismo mediante el cual estos compuestos potencian la contractilidad del músculo cardíaco. Durante la fase de meseta del potencial de acción (fase 2), el gradiente de concentración de K+^ a través de la membrana celular es prácticamente el mismo que du- rante la fase 4. Sin embargo, Vm será ahora positivo. Por tanto, existe un importante gradiente, que facilita la sali- da de K+^ de la célula (v. fig. 16-3). Si gK fuera la misma durante la meseta y la fase 4, la salida de K+^ durante la
296 Berne y Levy. Fisiología A B C LBC = 300 ms LBC = 8.000 ms 0 Epi 0 Medio 0 Endo 50 mV 200 ms ● Figura 16-5. Potenciales de acción registrados en las re- giones epicárdica ( A ), miocárdica media ( B ) y endocárdica ( C ) de la pared libre del ventrículo izquierdo canino. Las preparaciones fueron realizadas para una longitud bá- sica del ciclo (LBC) de 300 y 8. ms. (De Liu D-W et al. Circ Res 72:671, 1993.) A NIVEL CELULAR Para fomentar la gCa, las catecolaminas se unen en primer lugar a los receptores β-adrenérgicos de la membrana de la célula cardíaca. Esta interacción estimula la enzima de membrana adenilato ciclasa, que aumenta la concentra- ción intracelular de AMPc (v. también capítulo 3). Este in- cremento de la concentración de AMPc activa la protein- cinasa dependiente de AMPc, que, a su vez, aumenta la fosforilación de los canales del Ca++^ de tipo L de la mem- brana, lo que aumenta la entrada de Ca++^ a las células (fig. 16-6). Por el contrario, la acetilcolina interacciona con los receptores muscarínicos de la membrana celular para inhibir la adenilil ciclasa y, de este modo, antagoniza la activación de los canales del Ca++^ y reduce la gCa. APLICACIÓN CLÍNICA Los antagonistas del calcio son sustancias que bloquean los canales del Ca++. Entre estos fármacos se encuentran el verapamilo, la amlodipina y el diltiazem. Estos compuestos reducen gCa y, de este modo, dificultan la entrada de Ca++^ a las células miocárdicas. Los antagonistas del calcio reducen la duración de la meseta del potencial de acción y disminu- yen la potencia de la contracción cardíaca (fig. 16-7). Los antagonistas del calcio también reducen la contracción del músculo liso vascular, por lo que inducen una vasodilata- ción generalizada. Esta menor resistencia vascular reduce la fuerza (poscarga) que se contrapone a la propulsión de sangre desde los ventrículos hacia el sistema arterial, como se explica en el capítulo 17. Por tanto, los fármacos vasodi- latadores, como los antagonistas del calcio, suelen denomi- narse fármacos reductores de la poscarga.
298 Berne y Levy. Fisiología Fase 4 EK Fase^2 2 0
C orr ient e de K +^ (nA) Vm (mV) –120 –80 –40 0 40 ● Figura 16-9. Corriente de entrada de K+^ rectificada regis- trada en un miocito ventricular cuando se cambió el potencial desde un potencial mantenido en –80 mV a diversos potenciales de prueba. Los valores positivos en el eje vertical reflejan corrien- tes de salida, y los valores negativos, corrientes de entrada. La coordenada Vm del punto ( círculo hueco ) en el que la curva se cruza con el eje de las X es el potencial inverso, que indica el potencial de equilibrio de Nernst (Ek), momento en el que las fuerzas químicas y electrostáticas son iguales. (Reproducido de Giles WR, Imaizumi Y. J Physiol [Lond] 405:123, 1988.)
A
C
B
Ventrículo 1 00 ms
0
–1 00 0
–1 00 200 ms 1 00 ms Aurícula Nódulo SA 1 1 0 0 0 2 2 3 3 3 4 4 4 ● Figura 16-10. Potenciales de acción característicos (en mi- livoltios) registrados en células del ventrículo ( A ), del nódulo sinoau- ricular ( B ) y de la aurícula ( C ). Obsérvese que la calibración temporal en B es distinta a la de A y C. (De Hoffman BF, Cranefield PF. Electro- physiology of the Heart, Nueva York, McGraw-Hill, 1960.) yen al proceso de repolarización final (fase 3), como se describe más adelante. Existen dos tipos de canales IK según su velocidad de activación. El canal activador más lento se llama canal IKs , mientras que el que se activa más rápido se llama canal IKr (v. fig. 16-8). La duración del po- tencial de acción en los miocitos de distintas regiones del miocardio ventricular está determinada en parte por las distribuciones relativas de estos canales IKr e IKs. La meseta del potencial de acción persiste siempre que el flujo de salida de carga correspondiente principal- mente a K+^ se contrarreste con una entrada de cargas, que corresponde principalmente al Ca++. Los efectos de alterar este equilibrio se demuestran por la acción del calcio antagonista diltiazem sobre un preparado de múscu lo papilar aislado (v. fig. 16-7). Al aumentar las concentraciones de diltiazem, el voltaje de la meseta se va haciendo progresivamente menos positivo, y la dura- ción de la meseta se reduce. Por el contrario, la adminis- tración de determinados antagonistas de los canales del K+^ condiciona una prolongación notable de esta meseta.
Génesis de la repolarización final (fase 3)
El proceso de repolarización final (fase 3) se inicia al ter- minar la fase 2, cuando la salida de K+^ de la célula cardía- ca empieza a superar la entrada de Ca++. Como se ha co- mentado, por lo menos tres corrientes de salida de K (Ito, IK e IK1) contribuyen a la repolarización final (fase 3) de la célula cardíaca (v. figs. 16-3 y 16-8). La corriente de salida transitoria (Ito) y las corrientes rectificadoras tardías (IKr e IKs) ayudan a iniciar la repola- rización. Por tanto, estas corrientes son importantes de- terminantes de la duración de la meseta. Por ejemplo, la duración de la misma es notablemente menor en los mio- citos auriculares que en los ventriculares (fig. 16-10), porque la magnitud de Ito durante la meseta es mayor en los primeros. Como se ha comentado, la duración del po- tencial de acción en los miocitos ventriculares varía no- tablemente según la localización de los mismos dentro de las paredes ventriculares (v. fig. 16-5). Las corrientes Ito y rectificadora tardía (IK) explican principalmente es- tas diferencias. En los miocitos endocárdicos en los que la duración del potencial de acción es menor, la magni- tud de IK es máxima. Lo contrario sucede en los miocitos del tercio medio del miocardio. La magnitud de IK y la duración del potencial de acción son intermedios en los miocitos del epicardio. La corriente de entrada rectificadora de K+^ IK1 no parti- cipa en el inicio de la repolarización porque la conduc- tancia de estos canales es muy pequeña en el rango de valores de Vm que prevalecen durante la meseta. Sin em- bargo, los canales IK1 contribuyen de forma notable a la
Capítulo 16 Elementos de la función cardíaca 299 © ELSEVIER. Fotocopiar sin autorización es un delito. 100 mV 1 s A B C D E ● Figura 16-11. Efecto de la tetrodotoxina, que bloquea los canales rápidos del Na+, sobre los potenciales de acción registra- dos en una fibra de Purkinje. La concentración de tetrodotoxina era de 0 M en A, 3 × 10 -8^ M en B, 3 × 10 -7^ M en C y 3 × 10 -6^ M en D y E; E se registró más tarde que D. (Reproducido de Carme- liet E, Vereecke J. Pflügers Arch 313:300, 1969.) Zona polarizada Zona despolarizada Propagación
● Figura 16-12. Papel de las corrientes locales en la propa- gación de una onda de excitación por la fibra cardíaca. velocidad de repolarización cuando se ha iniciado la fase 3. Conforme Vm se va haciendo cada vez más negativa du- rante la fase 3, la conductancia de los canales que produ- cen la corriente IK1 aumenta de forma progresiva y esto acelera la repolarización (v. fig. 16-3).
Recuperación de las concentraciones iónicas
(fase 4)
La entrada constante y rápida de Na+^ a la célula durante la fase 0, y más lenta durante todo el ciclo cardíaco, des- polarizaría de forma gradual el voltaje de membrana en reposo si no fuera por la ATPasa Na + -K +, presente en la membrana celular (v. el capítulo 1). De modo similar, la mayor parte del exceso de iones Ca++^ que entran en la célula, principalmente durante la fase 2, se eliminan so- bre todo a través de un sistema de transporte inverso 3Na+-1Ca++, que intercambia tres iones de Na+^ por uno de Ca++. Sin embargo, algunos de los iones de Ca++^ se elimi- nan por la bomba de Ca++^ dependiente de ATP.
Potenciales de acción de respuesta lenta
Como se ha descrito anteriormente, los potenciales de ac- ción de respuesta rápida (v. fig. 16-1, A ) tienen cuatro com- ponentes principales: una fase ascendente (fase 0), una repolarización parcial precoz (fase 1), una meseta (fase 2) y una repolarización final (fase 3). Sin embargo, en el po- tencial de acción de respuesta lenta (v. fig. 16-1, B ) la pen- diente de la fase ascendente es mucho menor, no se reco- noce la fase de repolarización precoz, y la meseta es más corta y menos plana. Además, el paso de la meseta a la fase de repolarización final es menos definido. El bloqueo de los canales rápidos del Na+^ con tetrodo- toxina en una fibra de respuesta rápida puede generar respuestas lentas en condiciones apropiadas. El poten- cial de acción de la fibra de Purkinje que se muestra en la figura 16-11 refleja con claridad dos tipos de respuesta. En el registro control (A), el típico potencial de acción de respuesta rápida, presenta una prominente incisura por la corriente Ito que separa la fase ascendente de la mese- ta. En los potenciales de acción B a E la administración de cantidades cada vez más altas de tetrodotoxina deter- mina un bloqueo gradual de los canales rápidos del Na+. La fase ascendente y la incisura son cada vez menos pro- minentes en los potenciales de acción B a D. En el E, la incisura ha desaparecido por completo y la fase ascen- dente es muy gradual: este potencial de acción recuerda el de respuesta lenta típico. Algunas células cardíacas, sobre todo las de los nódu- los SA y AV, muestran potenciales de acción de respuesta lenta. En estas células, la despolarización se consigue so- bre todo por el flujo de entrada de Ca++^ a través de los canales del Ca++^ de tipo L en lugar de por la entrada de Na+^ a través de los canales rápidos del Na+. La repolariza- ción en estas fibras se debe a la inactivación de los cana- les del Ca++^ y el aumento de la conductancia al K+^ a través de los canales IK1 e IK (v. fig. 16-3).
CONDUCCIÓN EN LAS FIBRAS
CARDÍACAS
El potencial de acción que circula por las fibras musculares cardíacas se propaga mediante corrientes de circuito loca- les, del mismo modo que sucede en los nervios y las fibras musculares esqueléticas (v. capítulo 5). Cuando la onda de despolarización alcanza el extremo de la célula, el impulso se conduce a las células adyacentes a través de uniones en hendidura (v. capítulo 2). Los impulsos circulan con mayor facilidad a lo largo de la longitud de la célula (isotrópico) que de una célula a otra en sentido lateral (anisotrópico), dado que las uniones en hendidura se localizan principal- mente en los extremos celulares. Estos canales son bastan- te poco selectivos en la permeabilidad iónica, y muestran una baja resistencia eléctrica, lo que permite que la corrien- te iónica pase de una célula a otra. La resistencia eléctrica de las uniones en hendidura se parece a la citoplasmática. El flujo de cargas de una célula a la siguiente sigue los prin- cipios de las corrientes en los circuitos locales, y permite así la propagación intercelular del impulso.
Conducción de la respuesta rápida
Las características de la conducción son distintas en las fi- bras de respuesta lenta o rápida. En las rápidas, los canales rápidos del Na+^ se activan cuando el potencial transmem- brana de una región de la fibra experimenta un cambio sú- bito desde un valor de reposo de unos –90 mV hasta llegar a un umbral de –65 mV. La corriente de entrada de Na+^ pro- duce entonces una rápida despolarización de la célula en este lugar. Posteriormente, esta parte de la fibra se convier- te en parte de la zona despolarizada, y el límite se va des- plazando en consecuencia. En el nuevo límite, vuelve a ini- ciarse el proceso, que se repite una y otra vez, por lo que el margen se va desplazando de forma continua por la fibra a modo de onda de despolarización (fig. 16-12). La velocidad de conducción a lo largo de la fibra varía de forma directa en función de la amplitud del potencial de acción y la velocidad de cambio de potencial (dVm/dt) du- rante la fase 0. La amplitud del potencial de acción equi- vale a la diferencia de potencial entre las regiones total-
Capítulo 16 Elementos de la función cardíaca 301 © ELSEVIER. Fotocopiar sin autorización es un delito.
0 200 ms a b c
mV ● Figura 16-15. Efectos de la excitación en diversos momen- tos tras el inicio de un potencial de acción en una fibra de respues- ta lenta. En esta fibra, la excitación muy al final de la fase 3 (o a principios de la fase 4) induce una respuesta pequeña no propaga- da (local) (a). Cuando se produce más tarde dentro de la fase 4, puede conseguirse una respuesta propagada (b), pero de pequeña amplitud y poca pendiente en la fase ascendente; esta respuesta se conduce muy lentamente. Cuando todavía se produce más tarde dentro de la fase 4, se recupera la excitabilidad completa, y la respuesta (c) muestra características normales. (Modificado de Singer DH et al. Prog Cardiovasc Dis 24:97, 1981.) distintas a nivel cuantitativo de las que se observan en las respuestas rápidas. El potencial umbral es de unos –40 mV para la respuesta lenta, y la conducción es mucho más len- ta que en la respuesta rápida. Las velocidades de conduc- ción de la respuesta lenta en los nódulos SA y AV son de unos 0,02-0,1 m/s. La velocidad de conducción de respues- ta rápida es de 0,3-1 m/s en las células miocárdicas, y de 1-4 m/s en las fibras especializadas de conducción (fibras de Purkinje) de los ventrículos. Las respuestas lentas se bloquean con mayor facilidad que las rápidas, de forma que la conducción se interrumpe antes de que el impulso alcance el extremo de la fibra miocárdica. Además, las fi- bras de respuesta rápida pueden responder a frecuencias de repetición mucho mayores que las fibras lentas.
EXCITABILIDAD CARDÍACA
Dado el rápido desarrollo de marcapasos artificiales y otros dispositivos eléctricos para corregir las alteracio- nes del ritmo cardíaco, es fundamental un conocimiento detallado de la excitabilidad cardíaca. Las característi- cas de excitabilidad de las distintas clases de células car- díacas muestran notables diferencias, según que los po- tenciales de acción sean de respuesta lenta o rápida.
Respuesta rápida
Cuando se inicia la respuesta rápida, la célula despolari- zada ya no se puede excitar hasta que se repolarice de forma parcial (v. fig. 16-1, A ). El intervalo que transcurre desde que se inicia el potencial de acción hasta que la fibra puede conducir otro potencial de acción se deno- mina período refractario eficaz. En la respuesta rápida, este período va desde el principio de la fase 0 hasta un punto en la fase 3 en el que la repolarización alcanza unos –50 mV (fase 3 de la fig. 16-1, A ). Para este valor de la velocidad, muchos de los canales rápidos del Na+^ han pasado del estado inactivado al estado cerrado. Sin em- bargo, la fibra cardíaca no es totalmente excitable hasta que se repolariza por completo. Antes de la repolariza- ción completa (es decir, durante el período refractario relativo), sólo será posible inducir un potencial de ac- ción cuando el estímulo sea más potente que el que ge- neraría un potencial durante la fase 4. Cuando se induce una respuesta rápida durante el pe- ríodo refractario relativo de una excitación previa, sus ca- racterísticas varían según el potencial de membrana exis- tente en el momento de la estimulación (fig. 16-14). Cuanto más tarde dentro del período refractario se produce la esti- mulación de la fibra, mayor será el incremento de la ampli- tud de la respuesta y de la pendiente de la fase ascendente, porque el número de canales rápidos del Na+^ recuperados de la inactivación previa aumenta al progresar la repola- rización. En consecuencia, la velocidad de propagación también será tanto mayor cuanto más tarde se estimule la fibra dentro del período refractario relativo. Cuando la fi- bra está repolarizada por completo, la respuesta será constante, independientemente del momento de la fase 4 en el que se aplique el estímulo.
Respuesta lenta
En las fibras de respuesta lenta el período refractario re- lativo suele extenderse mucho más allá de la fase 3 (v. fi- gura 16-1, B ). Incluso cuando la célula está totalmente repolarizada, puede resultar difícil provocar una res- puesta propagada durante algún tiempo. Esta caracterís- tica de las fibras de respuesta lenta se denomina refrac- tariedad postrepolarización. Los potenciales de acción que se inducen en la fase pre- coz del período refractario relativo son pequeños, y su fase ascendente no tiene mucha pendiente (fig. 16-15). Las ampli- tudes y las pendientes de la fase ascendente mejoran de forma progresiva cuanto más tarde se inducen los potencia- les de acción dentro del período refractario relativo. La re- cuperación de la excitabilidad total es mucho más lenta que en la respuesta rápida. Los impulsos que llegan en la prime- ra fase del período refractario relativo tienen una conduc- ción mucho más lenta que los que aparecen más tarde en este período. Los prolongados períodos refractarios tam- bién ocasionan bloqueos de la conducción. Aunque se repi- tan las respuestas lentas a baja frecuencia, la fibra es sólo capaz de conducir un porcentaje pequeño de estos impul- sos; por ejemplo, en algunas circunstancias sólo se pueden propagar impulsos alternantes (v. más adelante).
EFECTOS DE LA DURACIÓN DEL CICLO
La duración del ciclo es el tiempo que transcurre entre potenciales de acción sucesivos. Los cambios en esta APLICACIÓN CLÍNICA En un paciente que sufre despolarizaciones prematuras ocasionales (fig. 16-32), el momento de aparición de es- tos latidos prematuros puede determinar consecuencias clínicas. Si se produce al final del período refractario rela- tivo de la despolarización precedente o tras la repolariza- ción completa, posiblemente esta despolarización prema- tura no tenga consecuencias. Sin embargo, si estas despolarizaciones prematuras se originan al principio del período refractario relativo de los ventrículos, la conduc- ción de este impulso prematuro desde su lugar de origen será lenta, y es más probable que se produzca una reen- trada. Si esta reentrada es irregular (p. ej., si se produce una fibrilación ventricular), el corazón será incapaz de bombear de forma eficaz y puede producirse la muerte.
302 Berne y Levy. Fisiología LC =^ 2. 000 ms DPA=^200 ms LC = 630 ms DPA = 180 ms LC = 400 ms DPA =^170 ms LC =^250 ms DPA =^140 ms LC = 200 ms DPA = 130 ms Fibras de Purkinje Haz de His Ventrículo izquierdo Músculo papilar Vena cava superior Nódulo sinoauricular Aurícula derecha Ventrículo derecho Fibras de Purkinje Ramas del haz Aurícula izquierda Nódulo auriculoventricular ● Figura 16-16. Efecto de los cambios en la longitud del ciclo (LC) sobre la duración del potencial de acción (DPA) en las fibras de Purkinje. (Modificado de Singer D, Ten Eick RE. Am J Cardiol 28:381, 1971.) ● Figura 16-17. El sistema de conducción del corazón. duración modifican la duración del potencial de acción en las células cardíacas (fig. 16-16; v. también fig. 16-5) y también cambian sus períodos refractarios. En conse- cuencia, los cambios de la duración del ciclo también son factores importantes para el inicio o terminación de algunas arritmias (ritmos cardíacos irregulares). Los cambios en la duración del potencial de acción conseguidos mediante reducciones escalonadas de la duración del ciclo desde 2.000 a 200 ms para una fibra de Purkinje se muestran en la figura 16-16. Conforme va dis- minuyendo la duración del ciclo, también lo hace la du- ración del potencial de acción. Esta correlación directa entre la duración del potencial de acción y la duración del ciclo está mediada por cambios en gK, que implican por lo menos a dos tipos de canales del K+, en concreto los que conducen las corrientes de K+^ rectificadoras tar- días IKr e IKs y los que se encargan de las corrientes tran- sitorias de salida de K+, Ito. La corriente IK se activa con valores de Vm cercanos a 0, pero la corriente se activa lentamente, sigue activa- da durante varios cientos de milisegundos y se inactiva también de forma lenta. Por tanto, cuando se reduce la duración del ciclo básico, cada potencial de acción tien- de a aparecer antes durante el período de inactivación de la corriente IK iniciada por el potencial de acción pre- vio. Por tanto, cuanto más corta sea la duración del ciclo básico, mayor será la corriente de salida de K+^ durante la fase 2, y más corta la duración del potencial de acción. La corriente Ito también influye sobre la relación entre la duración del ciclo y del potencial de acción. La co- rriente Ito se activa también con un potencial próximo a 0, y su magnitud varía de forma inversa según la dura- ción del ciclo cardíaco. Por tanto, cuando la duración del ciclo disminuye, el aumento de la corriente de salida de K+^ acorta la fase de meseta.
EXCITACIÓN NATURAL DEL CORAZÓN
Y ELECTROCARDIOGRAMA
La excitación del corazón suele producirse de forma or- denada, lo que permite un bombeo eficaz de la sangre. Esta excitación ordenada se produce por el sistema de conducción cardíaco (fig. 16-17). El nódulo SA es el mar- capasos del corazón, e inicia la diseminación de los po- tenciales de acción por las aurículas. Esta diseminación de la excitación llega al nódulo AV, en el que la conduc- ción se retrasa, de forma que se produce la contracción auricular y los ventrículos se pueden llenar bien. La exci- tación pasa después con rapidez a los ventrículos a tra- vés de las fibras de Purkinje, de modo que los miocitos ventriculares se contraen de forma coordinada. A conti- nuación se describen las propiedades de cada compo- nente del sistema de conducción del corazón. El sistema nervioso autónomo controla diversos as- pectos de la función cardíaca, como la frecuencia cardía- ca y la potencia de la contracción. Sin embargo, para la función cardíaca no es necesario que la inervación esté intacta. De hecho, un paciente trasplantado del corazón, cuyo nuevo corazón está totalmente denervado, puede adaptarse bien a situaciones de estrés. La capacidad del corazón trasplantado denervado de adaptarse a las con- diciones cambiantes se debe a ciertas propiedades del tejido cardíaco, sobre todo a su automatismo. Las propiedades de automatismo (capacidad de ini- ciar su propio latido) y ritmicidad (regularidad de la ac- tividad marcapasos) permite que el corazón perfundido pueda latir aunque se separe por completo del cuerpo. El latido cardíaco de los vertebrados tiene un origen miógeno. Si la vasculatura coronaria de un corazón rese- cado se perfunde de forma artificial con sangre o con una solución de electrólitos oxigenada, las contraccio- nes cardíacas rítmicas pueden persistir durante muchas horas. Por lo menos algunas células de las aurículas y de los ventrículos pueden iniciar latidos; estas células se localizan principalmente en los tejidos nodulares o en las fibras de conducción especializadas del corazón.
304 Berne y Levy. Fisiología Haci a fu er a Haci a^ de ntro 30 pA 30 mV 200 ms ICa If IK ● Figura 16-19. Los cambios del potencial transmembrana ( mitad superior ) que se observan en las células del nódulo SA se producen por tres corrientes principales ( mitad inferior ): 1) la co- rriente ICa; 2) la corriente de entrada inducida por hiperpolarización, If, y 3) la corriente de salida de K+, IK. El trazado verde con ruido muestra la corriente de membrana neta, y la evolución temporal aproximada de 1), la corriente de salida de K+, IK; 2) la corriente de entrada inducida por hiperpolarización If, y 3) la corriente ICa por los canales de Ca++^ de tipo L. La línea gruesa en rojo del trazado indica la magnitud y dirección de la If estimada. (Reproducido de Vn Gin- neken ACG, Giles W. J Physiol 434:57, 1991.) A NIVEL CELULAR La corriente «f» (If) de las células del nódulo SA cardíaco se activa por hiperpolarización, y está controlada por los nu- cleótidos cíclicos; se denomina HCN. Existen cuatro miem- bros de la familia de genes HCN y estos canales se encuen- tran en las neuronas del SNC que generan potenciales de acción de forma repetida. El segmento transmembrana 4 (S 4 ) tiene muchos aminoácidos de carga positiva que ac- túan como sensores de voltaje, como también se encuen- tran en los canales del Na+, K+^ y Ca++^ regulados por el vol- taje. El canal que se expresa de forma predominante en el corazón es el derivado del gen HCN4. Las mutaciones de los aminoácidos de S 4 y del factor que une S 4 con S 5 produ- cen notables cambios en la dependencia del voltaje de la activación, de forma que se necesita una hiperpolarización mayor para abrir el canal. Este efecto se parece al de la acetilcolina, y se ha predicho que la aparición de estas mu- taciones en el corazón humano podría ser la base de la bradicardia sinusal y del síndrome del seno enfermo. El incremento del potencial umbral retrasa la aparición de la fase 0 y se reduce la frecuencia cardíaca. De modo pare- cido, cuando se incrementa el máximo potencial negativo, se necesita más tiempo para alcanzar el potencial umbral, en el que la pendiente de la fase 4 no sufre cambios y, por eso, se reduce la frecuencia cardíaca.
Base iónica del automatismo
Varias corrientes iónicas contribuyen a la despolarización diastólica lenta que se produce de forma característica en las células con automatismo del corazón. En las células marcapasos del nódulo SA se producen por lo menos tres corrientes iónicas que contribuyen a esta despolarización diastólica lenta: a ) una corriente de salida de K+, IK; b ) una corriente de entrada de K+, If inducida por la hiperpolariza- ción, y c ) una corriente de entrada de Ca++^ ICa (fig. 16-19). Los disparos repetidos de las células marcapasos co- mienzan con la corriente rectificadora tardía de K+, IK. La salida de K+^ tiende a repolarizar la célula tras la fase as- cendente del potencial de acción. El K+^ sigue saliendo de la célula mucho después del momento de la repolariza- ción máxima, pero este flujo de salida se reduce durante la fase 4 (v. fig. 16-19). Cuando esta corriente disminuye, su oposición a los efectos despolarizantes de las dos corrien- tes de entrada (If e ICa) también disminuye de forma gra- dual. La progresiva despolarización diastólica está media- da por dos corrientes de entrada If e ICa, que se oponen al efecto repolarizador de la corriente de entrada IK. La corriente de entrada If se activa cerca del final de la repolarización y está mediada principalmente por el Na+ a través de unos canales específicos, distintos de los ca- nales rápidos del Na+. La corriente se llamó « funny » por- que sus descubridores no esperaban encontrar una co- rriente de entrada de Na+^ en las células marcapasos al final de la repolarización. Esta corriente se activa cuando el potencial de membrana se hiperpolariza por encima de –50 mV. Cuanto más negativo sea el potencial de mem- brana en este momento, mayor será la activación de If. La segunda corriente responsable de la despolariza- ción diastólica es la corriente de Ca++, ICa. Esta corriente se activa al final de la fase 4 cuando el potencial trans- membrana llega a un valor de unos –55 mV (v. fig. 16-19). Cuando se activan los canales del Ca++, aumenta la entra- da de calcio a la célula. Esta entrada acelera la velocidad de la despolarización diastólica, que determina la fase ascendente del potencial de acción. Una reducción de la [Ca++]o (fig. 16-20) o la adición de antagonistas de los ca- nales del calcio reduce la amplitud del potencial de ac- ción y la pendiente de la despolarización diastólica lenta en las células del nódulo SA. Pruebas recientes indican que la actividad marcapasos también puede estar media- da por otras corrientes iónicas, como una corriente de entrada de Na+^ mantenida (de fondo) (INa), la corriente de Ca++^ de tipo T o la corriente de intercambio Na/Ca es- timulada por la liberación espontánea de Ca++^ del retícu- lo sarcoplásmico (RS). Estas observaciones ilustran las múltiples formas de mantener esta vital función*. *La base iónica del automatismo de las células del nódulo AV se parece a la descrita en el nódulo SA. Mecanismos parecidos justifican el automatismo en las fibras de Purkinje ventriculares, salvo porque está más implicada la co- rriente rápida de Na+^ que la ICa. También se ha sugerido que una corriente de K+^ dependiente del tiempo y del voltaje es la implicada en la despolarización diastólica lenta, en lugar de la corriente If de entrada inducida por hiperpolari- zación; sin embargo, esto todavía no se ha demostrado.
Capítulo 16 Elementos de la función cardíaca 305 © ELSEVIER. Fotocopiar sin autorización es un delito.
- 60 0 Ca++^ 2 mM 500 ms Ca++^ 0,2 mM M ili vo ltio s ● Figura 16-20. Potenciales transmembrana registrados en una célula de marcapasos del nódulo SA. La concentración de Ca++^ se redujo en ambos baños de 2 a 0,2 mM. (Modificado de Kohlhardt M et al. Basic Res Cardiol 71:17, 1976.) Los neurotransmisores autónomos afectan al automa- tismo mediante la modificación de las corrientes iónicas en la membrana. Los transmisores adrenérgicos incre- mentan las tres corrientes implicadas en el automatismo del nódulo SA. Para aumentar la pendiente de la despola- rización diastólica, el incremento de If e ICa generado por los transmisores adrenérgicos debe superar la potencia- ción de Ik por estos mismos transmisores. La hiperpolarización inducida por la acetilcolina libera- da en las terminaciones del nervio vago a nivel cardíaco se consigue mediante la activación de unos canales del K+ específicos, los canales del K+^ regulados por la acetilcoli- na (KACh). Además, la acetilcolina deprime las corrientes If e ICa. En el capítulo 18 se describen con mayor detalle los efectos neurales autónomos sobre las células cardíacas. Cuando se reseca o destruye el nódulo SA u otros compo- nentes del complejo marcapasos auricular, las células mar- capasos del nódulo AV suelen asumir la función de marcapa- sos para todo el corazón. Pasado un tiempo, que puede durar de minutos a días, las células automáticas auriculares suelen asumir de nuevo el dominio, y reinician su función como marcapasos. Las fibras de Purkinje de las vías de con- ducción especializadas de los ventrículos muestran tam- bién automatismo. Es característico que estas fibras dispa- ren a una frecuencia muy lenta. Cuando la unión AV no consigue transmitir el impulso cardíaco desde las aurículas a los ventrículos, estos marcapasos idioventriculares de la red de fibras de Purkinje inician las contracciones ventricu- lares, pero a una velocidad de 30-40 latidos/minuto.
Supresión por superación
El automatismo de las células marcapasos disminuye cuando se excitan estas células a alta frecuencia. Este fenómeno se conoce como supresión por superación. Dado que la ritmicidad intrínseca del nódulo SA es ma- yor que la de cualquier otro marcapasos latente en el corazón, los disparos del nódulo SA tienden a suprimir el automatismo de estos otros focos. La supresión por superación se debe a la actividad de la ATPasa Na+-K+^ de la membrana. Una determinada can- tidad de Na+^ entra en la célula cardíaca en cada despola- rización. Cuanto mayor sea la frecuencia de despolariza- ción de la célula, mayor será la cantidad de Na+^ que entra en la célula por minuto. Cuando la frecuencia de excita- ción es alta, la actividad de la ATPasa Na+-K+^ tiende a sa- car esta mayor cantidad de Na+^ de la célula. La actividad de esta ATPasa hiperpolariza la célula, porque se extraen tres iones Na+^ por la bomba en intercambio por dos io- nes K+^ que entran a la célula (v. capítulo 1). Por tanto, la despolarización diastólica lenta necesita más tiempo para llegar al umbral de disparo. Además, cuando la su- presión por superación se interrumpe de forma súbita, la actividad de la ATPasa Na+-K+^ no se interrumpe de forma instantánea sino que permanece hiperactiva de for- ma temporal. La salida continuada de Na+^ se opone a la despolarización gradual de la célula marcapasos durante la fase 4, de forma que se suprime temporalmente el au- tomatismo intrínseco de la célula.
Conducción auricular
Desde el nódulo SA, el impulso cardíaco se dispersa de forma radial por toda la aurícula derecha (v. fig. 16-17) siguiendo las fibras miocárdicas auriculares normales, con una velocidad de conducción aproximada de 1 m/s. Una vía especial, la banda miocárdica interauricular an- terior (o haz de Bachmann), conduce los impulsos del nódulo SA directamente hacia la aurícula izquierda. La onda de excitación se dirige en sentido inferior a través de la aurícula derecha hasta alcanzar el nódulo AV (v. figu- ra 16-17) que, en condiciones normales, es la única vía de entrada de los impulsos cardíacos en los ventrículos. Cuando se comparan con los potenciales de acción registrados en una fibra ventricular típica, la meseta au- ricular (fase 2) es más corta y está menos desarrollada, APLICACIÓN CLÍNICA Algunas regiones del corazón distintas del nódulo SA pueden iniciar latidos en circunstancias especiales. Estos lugares se denominan focos o marcapasos ectópicos. Los focos ectópicos pueden convertirse en marcapasos cuan- do: a ) su propia ritmicidad se potencia; b ) la ritmicidad de los marcapasos de mayor orden se deprimen, o c ) todas las vías de conducción entre el foco ectópico y las regio- nes de mayor ritmicidad se bloquean. Los marcapasos ectópicos pueden comportarse como un mecanismo de seguridad ante un fallo de los centros marcapasos norma- les. Sin embargo, si un centro ectópico dispara mientras el centro marcapasos normal sigue funcionando, la activi- dad ectópica puede inducir alteraciones esporádicas del ritmo, como despolarizaciones prematuras, o alteraciones continuas del mismo, como las taquicardias paroxísticas (v. sección posterior). APLICACIÓN CLÍNICA Si un foco ectópico de una aurícula empezara a disparar a alta velocidad de forma súbita (p. ej., 150 impulsos/min) en un individuo con una frecuencia cardíaca normal de 70 lpm, el foco ectópico se convertiría en el marcapasos para todo el corazón. Si el foco ectópico rápido dejara de disparar de forma repentina, el nódulo SA seguiría quies- cente durante un corto período de tiempo, por la supre- sión por superación. El intervalo que va desde el final del período de superación hasta que el nódulo SA recupera la capacidad de disparar se denomina tiempo de recupera- ción del nódulo sinusal. En los pacientes con síndrome del seno enfermo, el tiempo de recuperación del nódulo si- nusal está prolongado, y el consiguiente período de asis- tolia (ausencia de latido cardíaco) puede ocasionar una pérdida de conciencia.
Capítulo 16 Elementos de la función cardíaca 307 © ELSEVIER. Fotocopiar sin autorización es un delito. 500 ms 50 Fibra del nódulo AV mV A 1 A 2 A 3 H St ● Figura 16-22. Efectos de un breve estímulo vagal (St) sobre el potencial de membrana registrado en una fibra del nó- dulo AV. Obsérvese que poco después de producirse la estimula- ción vagal, la membrana de la fibra se hiperpolarizó. La excitación auricular (A 2 ) que llegó al nódulo AV mientras la célula estaba hiperpolarizada no se condujo, lo que se reconoce por la ausencia de una despolarización en el electrocardiograma de His (H). Las excitaciones auriculares precedentes (A 1 ) y posteriores (A 3 ) a la excitación A 2 se condujeron al haz de His. (Reproducido de Maz- galev T et al. Am J Physiol 251:H631, 1986.) Puede producirse una conducción retrógrada a través del nódulo AV. Sin embargo, el tiempo de conducción es significativamente más largo, y el impulso se puede blo- quear a frecuencias de repetición menores cuando el im- pulso se conduce en dirección retrógrada en lugar de anterógrada. Por último, el nódulo AV es lugar frecuente de reentradas (v. más adelante). Igual que sucede en el nódulo SA, el sistema nervioso autónomo regula la conducción AV. Una débil actividad vagal puede simplemente prolongar el tiempo de con- ducción AV. Por tanto, para cualquier duración determi- nada del ciclo auricular, el tiempo de conducción entre las aurículas y el haz de His (A-H) o entre las aurículas y los ventrículos (A-V) se prolonga por la estimulación va- gal. Una actividad vagal más intensa puede determinar que algunos o todos los impulsos procedentes de la aurí- cula queden bloqueados en el nódulo. El patrón de con- ducción en el que ninguno de los impulsos auriculares llega a los ventrículos se denomina bloqueo AV de tercer grado o completo (v. más adelante). El retraso o ausen- cia de conducción a través de la unión AV inducido por la actividad vagal se produce principalmente en la región N del nódulo. Este efecto de la estimulación vagal refleja la acción de la acetilcolina, que hiperpolariza el poten- cial de membrana de las fibras de conducción de la re- gión N (fig. 16-22). Cuanto mayor es la hiperpolarización en el momento en que llega el impulso auricular, más al- terada estará la conducción AV. Por el contrario, las fibras cardíacas simpáticas facili- tan la conducción AV. Estas fibras reducen el tiempo de conducción AV y potencian la ritmicidad de los marcapa- sos latentes en la unión AV. La noradrenalina liberada en las terminaciones nerviosas simpáticas posganglionares aumenta la amplitud y la pendiente de la fase ascendente de los potenciales de acción del nódulo AV, sobre todo en las regiones AN y N del nódulo.
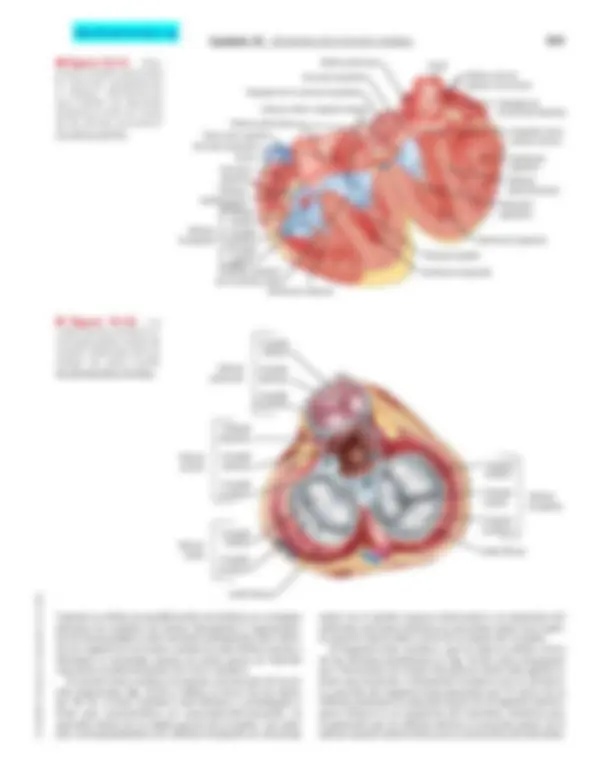
Conducción ventricular
El haz de His desciende aproximadamente 1 cm de forma subendocárdica por el lado derecho del tabique interven- tricular, y se divide en las ramas derecha e izquierda (v. fi- gura 16-17). La rama derecha del haz, que es continuación directa del haz de His, sigue descendiendo por el lado dere- cho del tabique interventricular. La rama izquierda, que es notablemente más gruesa que la derecha, se origina casi perpendicular al haz de His y perfora el tabique interventri- cular. En la región subendocárdica del lado izquierdo del tabique interventricular, la rama izquierda se divide en una división anterior, delgada, y otra posterior, gruesa. La rama derecha del haz y las dos divisiones de la rama izquierda se acaban subdividiendo en una comple- ja red de fibras de conducción, denominadas fibras de Purkinje, que se dispersan por encima de las superficies subendocárdicas de los dos ventrículos. Las fibras de Purkinje tienen sarcómeros abundantes dispuestos de forma lineal, igual que los miocitos. Sin embargo, el sistema de túbulos T, que está bien desarro- llado en los miocitos, no existe en las fibras de Purkinje de muchas especies. Las fibras de Purkinje son las cé- lulas más anchas del corazón, con diámetros de 70 a 80 μm, que contrastan con los 10-15 μm de los miocitos ventriculares. En parte, este gran diámetro es responsa- ble de que la velocidad de conducción en las fibras de Purkinje (1,4 m/s) sea superior a la que se observa en cualquier otro tipo de fibra dentro del corazón. Esta ma- yor velocidad de conducción permite una activación rá- pida de toda la superficie endocárdica ventricular. Los potenciales de acción registrados en las fibras de Purkinje se parecen a los observados en las fibras miocárdi- cas ventriculares normales. Sin embargo, dado que el perío- do refractario de estos potenciales de acción de las fibras de Purkinje es más largo, muchas excitaciones prematuras de las aurículas se conducen a través de la unión AV, pero quedan bloqueados en las fibras de Purkinje. El bloqueo de estas excitaciones auriculares impide la contracción ventri- cular prematura. Esta función de protección ventricular frente a los efectos de una despolarización auricular pre- matura es especialmente importante cuando la frecuencia cardíaca es lenta, porque la duración del potencial de acción y el período refractario eficaz de las fibras de Pur- kinje varían de forma inversa con la frecuencia cardíaca (v. fig. 16-16). Cuando la frecuencia cardíaca es baja, el pe- ríodo refractario eficaz de las fibras de Purkinje es especial- mente polongado*. A diferencia de lo que sucede con las
- Se observan cambios direccionales del período refractario similares en los miocitos ventriculares como respuesta a los cambios de frecuencia. APLICACIÓN CLÍNICA Se pueden producir alteraciones en la conducción de im- pulsos por la rama derecha o izquierda del haz o en cual- quiera de las ramas del haz izquierdo. Los bloqueos de la conducción pueden afectar a una o más de estas vías de conducción como consecuencia de una arteriopatía co- ronaria o procesos degenerativos asociados con el envejeci- miento, y ocasionan unos patrones del ECG característicos. El bloqueo de cualquiera de las ramas principales del haz se denomina bloqueo de la rama derecha o izquierda. El blo- queo de cualquiera de las divisiones de la rama izquierda se denomina hemibloqueo izquierdo anterior o posterior.
308 Berne y Levy. Fisiología S R C L S R C L S R C L S R C L
A B
C D
● Figura 16-23. Papel del bloqueo unidireccional en la reentrada. En A la onda de excitación que circula por un solo haz (S) de fibras continúa por las ramas izquierda (I) o derecha (D). La onda de despolarización entra en la rama conectora (C) desde ambos extremos, y se extingue en la zona de colisión. En B la onda queda bloqueada en las ramas D e I. En C existe un bloqueo bidireccional en la rama D. En D existe un bloqueo unidireccional en la rama D. Se bloquean los impulsos anterógrados, pero los retrógrados se conducen y entran de nuevo en el haz S. células de Purkinje, el período refractario eficaz de las célu- las del nódulo AV no sufre cambios apreciables dentro de los valores normales de frecuencia cardíaca y, de he- cho, aumenta con las frecuencias cardíacas muy rápi- das. Por tanto, cuando la aurícula se excita a frecuen- cias de repetición altas, es el nódulo AV el responsable de proteger a los ventrículos frente a esta frecuencia excesivamente elevada. Las primeras porciones de los ventrículos que se exci- tan por los impulsos que proceden del nódulo AV son el tabique interventricular (salvo en su parte basal) y los músculos papilares. La onda de activación se dispersa por la sustancia del tabique desde sus superficies endo- cárdicas derecha e izquierda. La contracción precoz del tabique aumenta su rigidez y le permite servir como pun- to de anclaje durante la contracción del resto del miocar- dio ventricular. Además, una contracción precoz de los músculos papilares impide la eversión de las válvulas AV hacia las aurículas durante la sístole ventricular. Las superficies endocárdicas de ambos ventrículos se activan con rapidez, pero la onda de excitación se extien- de desde el endocardio al epicardio a una velocidad me- nor (unos 0,3-0,4 m/s). La superficie epicárdica del ven- trículo derecho se activa antes que la izquierda, dado que el grosor de la pared ventricular derecha es notable- mente menor que en el izquierdo. Además, las regiones epicárdicas apical y central de ambos ventrículos se ac- tivan un poco antes que las correspondientes regiones basales. La última región de los ventrículos que se excita es la región epicárdica basal posterior y una pequeña zona en la parte basal del tabique interventricular.
Reentrada
Las condiciones que se exigen para que se produzca reentrada se muestran en la figura 16-23. En cada uno de los cuatro esquemas se divide un haz único de fibras car- díacas (S) en dos ramas, una derecha (R) y otra izquier- da (L). Existe un haz de comunicación (C) entre las dos ramas. En condiciones normales, un impulso que des- ciende por el haz S se conduce por las ramas R y L (v. fi- gu ra 16-23, A ). Cuando el impulso alcanza la rama de co- ne xión C, entra desde los dos lados y se extingue en el punto de colisión. El impulso procedente del lado izquier- do no puede progresar porque el tejido distal está en pe- ríodo refractario absoluto, ya que se acaba de despolari- zar desde el otro lado. El impulso tampoco puede atravesar el haz C desde el lado derecho por el mismo motivo. La figura 16-23, B muestra que el impulso no puede circular por todo el circuito cuando existe un bloqueo anterógrado en las ramas L y R del haz de fibras. Ade- más, si se produce un bloqueo bidireccional en cualquier punto del circuito (p. ej., en la rama R de la fig. 16-23, C ), el impulso no puede reentrar. Una condición necesaria para que se produzca reentra- da es que en algún punto del circuito el impulso pueda pasar en una dirección, pero no en la contraria, fenómeno conocido como bloqueo unidireccional. Como se muestra en la figura 16-23, D , el impulso puede desplazarse con normalidad por la rama L, pero se queda bloqueado en dirección anterógrada en la rama R por algún cambio pa- tológico en las células miocárdicas de esta rama. El impul- so que se estaba conduciendo por la rama L y la rama de conexión C puede penetrar entonces en la región deprimi- da de la rama R en sentido retrógrado, incluso aunque el impulso anterógrado haya quedado bloqueado previa- mente a este mismo nivel. ¿Por qué se bloquea el impulso anterógrado, pero no el retrógrado? La razón es que el an- terógrado llega a la región deprimida de la rama R antes que el retrógrado, porque la longitud del trayecto para el primero es mucho menor que el trayecto que debe reco- rrer el impulso retrógrado. Por tanto, el impulso anteró- grado puede quedar bloqueado sencillamente porque lle- ga a la región bloqueada durante su período refractario eficaz. Si se produce un retraso suficiente del impulso re- trógrado, el período refractario puede haber terminado en la región afectada, y esto permite la conducción del impulso por esta región y su regreso a la rama S. Aunque para que se produzca la reentrada es preciso un bloqueo unidireccional, este bloqueo no puede causar la re- entrada por sí solo. Para que ésta tenga lugar, el período re- fractario eficaz de la región de reentrada debe ser más corto que el tiempo de conducción por todo el circuito. En la figu- ra 16-23, D si el tejido situado justo distal a la zona deprimi- APLICACIÓN CLÍNICA En determinadas condiciones, el impulso cardíaco puede volver a excitar una región miocárdica que ya había atra- vesado previamente. Este fenómeno, que se denomina reentrada, es responsable de muchas arritmias clínicas (al- teraciones del ritmo cardíaco). La reentrada puede ser or- denada o aleatoria. En la variante ordenada, el impulso viaja por una vía anatómica fija, mientras que en la alea- toria esta vía cambia de forma continua.
310 Berne y Levy. Fisiología A B C D 0 MV 100 LCB = 800 LCB = 700 LCB = 600 LCB = 500 ● Figura 16-25. Potenciales de acción transmembrana re- gistrados en las fibras de Purkinje. Se añadió acetilestrofantidina, un glucósido cardíaco, al baño y se produjeron secuencias de seis latidos conducidos (que se marcan con puntos), con una longitud de ciclo básica (LCB) de 800 ( A ), 700 ( B ), 600 ( C ) y 500 ( D ) mili- segundos. Observar que se produjeron pospotenciales tardíos tras los latidos conducidos, y que estos pospotenciales alcanzaron el umbral tras el último latido conducido en B a D. (Tomado de Ferrier GR et al. Circ Res 32:600, 1973.) La [Ca++]i elevada puede activar también el sistema de transporte inverso 3Na+-1Ca++. Este sistema de transpor- te inverso electrógeno, que introduce tres iones Na+^ en la célula por cada ión Ca++^ que extrae de ella, también gene- ra una corriente neta de entrada de cationes que contri- buye a la aparición de las PDPT.
ELECTROCARDIOGRAFÍA
El ECG permite a los médicos averiguar el trayecto del impulso cardíaco mediante el registro de las variaciones de potencial eléctrico en diversos lugares de la superficie corporal. Mediante el análisis de los detalles de estas fluc- tuaciones en el potencial eléctrico, se consigue importan- te información sobre: a ) la orientación anatómica del co- razón; b ) los tamaños relativos de las cámaras; c ) los diversos trastornos del ritmo y la conducción; d ) la exten- sión, localización y progresión de las lesiones isquémicas del miocardio); e ) los efectos de las alteraciones de las concentraciones de electrólitos, y f ) la influencia de deter- minados fármacos (sobre todo, digital, antiarrítmicos y antagonistas del calcio). Dado que la electrocardiografía es una materia amplia y compleja, en esta sección sólo se abordan sus principios más elementales.
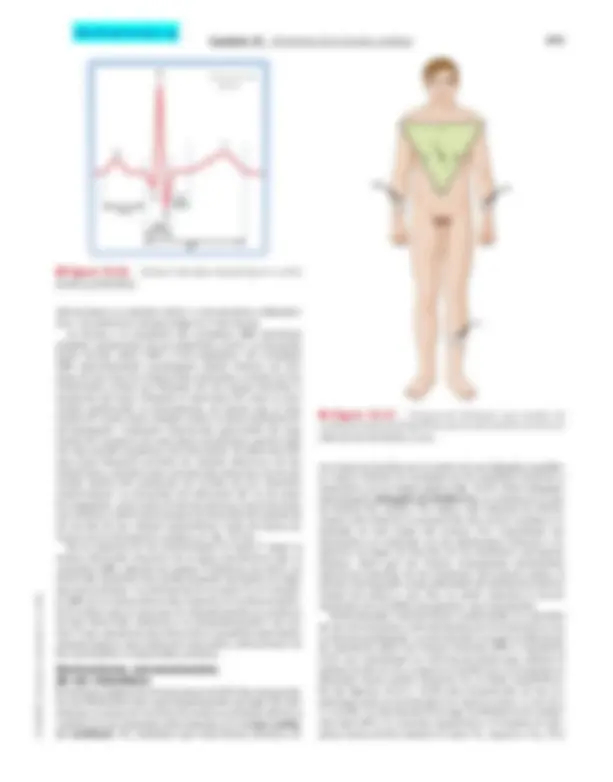
Electrocardiografía escalar
En la electrocardiografía, una derivación es la conexión eléctrica desde la piel del paciente a un dispositivo de registro ( electrocardiógrafo ) que mide la actividad eléc- trica del corazón. El sistema de derivaciones que se em- plea para registrar un ECG convencional se orienta en algunos planos del cuerpo determinados. Los diversos acontecimientos eléctricos que tienen lugar en el cora- zón en un momento determinado pueden representarse con un vector tridimensional (una cantidad con magni- tud y dirección). Un sistema de derivaciones de registro orientado en un plano determinado detecta sólo la pro- yección del vector tridimensional sobre este plano. La diferencia de potencial entre dos electrodos de registro representa la proyección del vector sobre la línea entre las dos derivaciones. Los componentes de los factores que se proyectan sobre estas líneas no son vectores, sino cantidades escalares (que tienen magnitud, pero no dirección). Por tanto, el registro de cambios en la dife- rencia de potencial entre dos puntos de la superficie cu- tánea a lo largo del tiempo se denomina ECG escalar. Un ECG escalar detecta cambios temporales en el po- tencial eléctrico entre algún punto de la superficie de la piel y un electrodo indiferente, o entre pares de puntos de la superficie cutánea. El impulso cardíaco circula por el corazón siguiendo un patrón tridimensional complejo. Por tanto, la configuración exacta del ECG varía de un paciente a otro, y en el mismo individuo lo hace según la localización anatómica de las derivaciones. La represen- tación gráfica del impulso eléctrico registrado por el ECG se denomina trazado. En general, en el trazado se observan unas ondas P, QRS y T (fig. 16-26). La onda P refleja la despolarización que se extiende por las aurículas, la onda (o complejo) QRS se corresponde con la despolarización de los ven- trículos, y la onda T representa la repolarización de los ventrículos (se produce una repolarización de las aurícu- las, que queda oculta dentro de la despolarización ven- tricular). El intervalo PR (o para ser más exactos, PQ) es una medida del tiempo transcurrido desde el comienzo de la activación auricular hasta el comienzo de la activa- ción ventricular; su duración normal oscila entre 0,12 y 0,20 segundos. Una gran parte de este tiempo correspon- de al paso del impulso a través del sistema de conduc- ción AV. Las prolongaciones patológicas del intervalo PR se asocian con alteraciones de la conducción AV. Estas
Capítulo 16 Elementos de la función cardíaca 311 © ELSEVIER. Fotocopiar sin autorización es un delito. P-R QRS Q P R S T QT ST 0,1 s LA LL RA II I III
- (^) – ● Figura 16-26. Ondas e intervalos importantes en un ECG escalar característico. ● Figura 16-27. Triángulo de Einthoven que muestra las conexiones electrocardiográficas para las derivaciones convencio- nales de los miembros I, II y III. alteraciones se pueden deber a mecanismos inflamato- rios, circulatorios, farmacológicos o nerviosos. La forma y la amplitud del complejo QRS muestran amplias variaciones de un individuo a otro. La duración suele oscilar entre 0,06 y 0,10 segundos. Un complejo QRS anormalmente prolongado puede indicar un blo- queo de las vías de conducción normales a través de los ventrículos (como un bloqueo de las ramas derecha o izquierda del haz). Durante el intervalo ST, todo el mio- cardio ventricular se despolariza, de forma que el seg- mento ST suele estar situado sobre la línea isoeléctrica, normalmente. Cualquier desviación apreciable del seg- mento ST respecto de esta línea isoeléctrica puede indi- car una lesión isquémica del miocardio. El intervalo QT, que suele llamarse período de «sístole eléctrica» de los ventrículos, muestra una correlación estrecha con la du- ración media del potencial de acción de los miocitos ventriculares. La duración del intervalo QT es de unos 0,4 segundos, pero varía de forma inversa con la frecuen- cia cardíaca, sobre todo porque la duración del potencial de acción de las células miocárdicas varía de forma in- versa con la frecuencia cardíaca (v. fig. 16-16). En la mayoría de las derivaciones la onda T sigue la misma dirección respecto de la línea isoeléctrica que el complejo QRS, aunque las ondas T bifásicas (es decir, en dirección opuesta) son perfectamente normales en algu- nas derivaciones. La desviación de la onda T y el comple- jo QRS en la misma dirección respecto de la línea isoeléc- trica indica que el proceso de repolarización se produce en una dirección contraria a la despolarización. Las on- das T que muestran una dirección o amplitud anormales pueden indicar una lesión del miocardio, alteraciones de los electrólitos o hipertrofia cardíaca.
Derivaciones convencionales
de los miembros
El sistema original de derivaciones del ECG fue desarrolla- do por Einthoven hace aproximadamente un siglo. En este sistema, la suma de vectores de toda la actividad eléctrica cardíaca en un momento determinado es el vector cardía- co resultante. Se considera que esta fuerza eléctrica di- reccional se localiza en el centro de un triángulo equiláte- ro cuyos vértices se localizan en los hombros derecho e izquierdo y en la región púbica (fig. 16-27). Este triángulo, denominado triángulo de Einthoven , se orienta en el pla- no frontal del cuerpo. Por tanto, este sistema de deriva- ciones sólo detecta la proyección del vector cardíaco re- sultante en este plano del cuerpo. Por comodidad, los electrodos se conectan en los antebrazos derecho e iz- quierdo en lugar de hacerlo en los hombros correspon- dientes, dado que los brazos representan extensiones eléctricas sencillas de los hombros. Del mismo modo, la pierna corresponde a una extensión del sistema de deriva- ciones del pubis y, por ello, se suele conectar el tercer electrodo en el tobillo (en general, en el izquierdo). Determinadas convenciones condicionan la conexión de las derivaciones convencionales de los miembros con el electrocardiógrafo. La derivación I recoge la diferencia de potencial entre los brazos derecho (RA) e izquierdo (LA). Las conexiones se colocan de forma que cuando el potencial del LA (VLA) supera el del RA (VRA) el trazado se desplaza hacia arriba respecto de la línea isoeléctrica. En las figuras 16-27 y 16-28 esta disposición de las co- nexiones para la derivación I se marca como (+) en LA y (–) en RA. La derivación II recoge la diferencia de poten- cial entre RA y LL (pierna izquierda) y el trazado se des- plaza hacia arriba cuando el valor VLL supera a VRA. Por